
Abstract
Abstract
Berberine, a Chinese medical herbal extract, plays a key role in antidiabetic, antiangiogenesis, anti-inflammatory, antimicrobial, anticancer, and antihypercholesterolemic. Our previous studies revealed that berberine exerted odontoprotective effect by increasing odontoblast differentiation. However, the mechanisms involved in the odontoprotective effect of berberine have not been fully explored. The Wnt/β-catenin pathway is involved in odontoblast differentiation of dental pulp stem cells (DPSCs). If β-catenin is nuclear translocation, the Wnt/β-catenin pathway is activation. In this study, DPSCs were treated with or without berberine. Then, we examined the accelerative effects of berberine on odontoblast differentiation and mineralized nodules formation by real-time polymerase chain reaction, alizarin red S staining, and alkaline phosphatase staining. In addition, while treated with berberine, β-catenin translocated to the nucleus evaluated by western blot and immunofluorescent staining. Our results revealed that berberine functions as a promoter of odontoblast differentiation by promoting Wnt/β-catenin pathway, suggesting that it may be useful in guiding therapeutic strategies for the treatment of dental caries.
Introduction
Mesenchymal stem cells (MSCs), isolated from several human tissues, have the ability of self-renewal and multilineage differentiations as potential for regenerative cell therapy. The differentiation capabilities of MSCs into osteocytes, chondrocytes, adipocytes, and odontoblasts as well as immunosuppressive properties have been widely certified (Trento and Dazzi, 2010). However, MSCs are associated with limitations: ethical issues, prone to tumor formation, viral pollution, the shortage of donated, and so on (Huang et al., 2009; Jeong et al., 2011). Thus, there is an urgent need to find new sources for MSCs.
Dental pulp stem cells (DPSCs) first were isolated from normal human extracted third molars and were known to have higher osteogenic, dentinogenic, adipogenic, chondrogenic, myogenic, neurogenic, and regenerative potentials than many other kinds of stem cells (Chen et al., 2016). DPSCs play a crucial role in tooth homoeostasis by giving rise differentiating into odontoblasts that induce reparative dentinogenesis in response to inflammatory or pathological stimuli in tooth lesions. Although dental caries or physical or chemical trauma may result in hurt or destruction of enamel and dentine, reparative dentin can still be formed to preserve the deep-seated pulp tissue. This regenerative process is controlled by the newly differentiated odontoblasts that result from DPSCs (Mitsiadis et al., 2011). However, by reason of the dental pulp having been heavily damaged, there are often shortage of quantity of endogenetic DPSCs to renovate major tooth lesions (Cao et al., 2015). Hence, a primary challenge in dental clinical treatment is to accelerate odontoblast differentiation of DPSCs.
Berberine, a Chinese medical herbal extract, is isolated from several Chinese herbs. Emerging evidence indicates that berberine plays a prominent role in antidiabetic, antiangiogenesis, anti-inflammatory, antimicrobial, anticancer, and antihypercholesterolemic (Dong et al., 2011; Lee et al., 2006; Peng et al., 2006; Wang et al., 2014).
Recent studies have shown that berberine is widely researched in preclinical and clinical tests, and several mechanisms, namely, enhancing insulin expression and β cell regeneration, promoting glucagon-like peptide release, increasing insulin-like growth factor (IGF), and controlling osteoclast and osteoblast function in vitro and in vivo (He et al., 2018; Hu et al., 2008; Lee et al., 2008; Ye et al., 2016; Yu et al., 2015). But whether berberine exerts any effects in odontoblast differentiation of DPSCs is still unknown. Therefore, in this study, we investigated whether berberine could control odontoblast differentiation of DPSCs and which signaling pathway was potentially associated with the function.
Materials and Methods
Cell cultures
All samples were acquired from healthy patients (18- to 25-year-old healthy donors) after obtaining the informed consents that were approved by the ethics committee of the Stomatological Hospital of Jilin University. Healthy pulp tissues (n = 20) were isolated from the caries-free teeth of patients without oral infection undergoing extraction of fully erupted third molars. Tooth surface was cleaned, cut around the cementoenamel junction by sterilized dental fissure burs, and opened to reveal the pulp chamber. The collected pulp tissue was digested in a solution of 3 mg/mL collagenase type I for 1 hour at 37°C to release the cells. Single-cell suspensions were obtained by passing the digested tissues through a 70-μm cell strainer (BD Falcon).
The isolated cells were seeded into 25 cm2 plates, cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS, Gibco-BRL, Life Technologies, Inc., Gaithersburg, MD), 100 U/mL penicillin and 100 μg/mL streptomycin and maintained in 5% CO2 at 37°C (Ledesma-Martinez et al., 2016). The medium was changed every 3 days. Approximately 7–10 days after seeding, the cells became nearly confluent. Cells were passaged at the ratio of 1:3 when they reached 80% confluence. Cells at the third to fifth passages were used in all the experiments. The stemness of DPSCs was evaluated by positive staining with anti-CD34, STRO-1 and c-kit, and the lack of expression of CD45 (Wang et al., 2018).
Odontoblast differentiation
DPSCs were seeded onto 35 mm culture dishes (Costar, Cambridge, MA). The following protocol was used to induce odontoblast differentiation of DPSCs differentiation: α-minimum essential medium (α-MEM; Invitrogen, Carlsbad, CA), 10 mmol/L β-glycerophosphate, 50 mg/mL α-ascorbic acid, 10 nmol/L dexamethasone (Sigma-Aldrich, St. Louis, MO), 0.292 mg/mL glutamine, 100 mg/mL streptomycin, and 100 U/mL penicillin G were added to the cells cultured in DMEM and 15% FBS, respectively, for 0, 7, and 14 days. The medium was replaced every 2 days. Then, the cells were collected for determination of the odontoblast differentiation by evaluating mRNA expression of odontogenic markers, including dentin matrix protein-1 (DMP1) and dentin sialophosphoprotein (DSPP).
The experimental treatment groups consisted of differentiation media with berberine (Sigma) and/or DKK-1 (Santa Cruz). After induced for 0, 7, and 14 days, cells were prepared for alizarin red S staining, alkaline phosphatase (ALP) staining, and immunofluorescence. Protein and RNA were extracted for western blot analysis and real-time polymerase chain reaction (RT-PCR).
Western blot analysis
After 0, 7, and 14 days of culture, total protein was extracted from the DPSCs cultured in the different groups by lysis buffer (NCM Biotech, Suzhou, China). The protein in the supernatant was extracted after centrifugation at 20,000 g for 20 minutes at 4°C, after which protein concentration was determined (BCA; Beyotime Institute of Biotechnology, Shanghai, China). For western blot analysis, equal amounts of proteins (30 mg) per sample were separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis and electrotransferred onto polyvinylidene fluoride membranes. The membranes were blocked with 5% nonfat dry milk in Tris-buffered saline Tween at room temperature for 2 hours, and then incubated with primary monoclonal antibodies at 4°C overnight.
Subsequently, the membranes were incubated with horseradish peroxidase-conjugated secondary antibody at room temperature for 2 hours. Proteins on the blots were detected using the Western-Light Chemiluminescent Detection System (Peiqing, China). GAPDH and β-tubulin were used as the internal control for the cytoplasmic and nuclear proteins. The following primary antibodies were used: GAPDH (anti-mouse; Santa Cruz), β-tubulin (anti-mouse; Sigma), β-catenin (anti-mouse; Cell Signaling), DMP1 (anti-rabbit; Santa Cruz), and runt-related transcription factor 2 (Runx2) (anti-rabbit; Santa Cruz).
Real-time polymerase chain reaction
Total cellular RNA was isolated from the cells and reverse transcribed using conventional protocols. PCR amplification was performed using the following primer sequences: GAPDH, forward: 5′-TCCATGACAACTTTGGTATCG-3′ and reverse: 5′-TGTAGCCAAATTCGTTGTCA-3′; DMP1, forward: 5′-TGGGGATTATCCTGTGCTCT-3′ and reverse: 5′-GCTGTCACTGGGGTCTTCAT-3′; DSPP, forward: 5′-GGAGACAAGACCTCCAAGAGTA-3′ and reverse: 5′-TGCTGGGACCCTTGATTTCTA-3′; Runx2, forward: 5′-AACCCACGAATGCACTATCCA-3′ and reverse: 5′-CGGACATACCGAGGGACCTG-3′. All the primer sequences were determined using established GenBank sequences. The primers were used to amplify the duplicate PCRs. Each sample was calculated from three independent experiments and GAPDH was used as an internal control.
Alizarin red S and ALP staining
The DPSCs were cultured in odontoblast differentiation medium with berberine or DKK-1. Subsequently, cells were fixed with 4% PFA for 1 hour, washed with PBS, and then stained with 40 mmol/L alizarin red S (pH = 4.2) for 10 minutes under conditions of gentle agitation. Absorbance of the extracted alizarin red S stain was measured at 570 nm. DPSCs were subjected to ALP staining using the ALP assay kit (Beyotime Institute of Biotechnology) according to the manufacturer's protocol.
Immunofluorescent staining
DPSCs were seeded into the 24-well plates, fixed with 4% PFA for 1 hour, rinsed with PBS containing 0.1% Triton X-100 (PBST), and blocked for 30 minutes in PBST supplemented with 10% FBS. Cells were incubated with a primary antibody against β-catenin (1:100) (anti-mouse; Cell Signaling) in the same solution overnight at 4°C. Then cells were rinsed and incubated with secondary antibodies for 2 hours at room temperature. Nuclei were stained with 4′,6-diamidino-2-phenylindole dihydrochloride (1:1000; Santa Cruz). Finally, the cells were examined using a Leica fluorescence microscope (Germany).
Statistical analysis
For data analysis of cytotoxicity assays, one-way analysis of variance followed by post hoc Tukey's test was applied. All experiments were analyzed by post hoc Tukey's test using SPSS software, version 19.0 (SPSS, Inc., Chicago, IL). Data are presented as the mean ± standard error of the mean. p < 0.05 was considered to indicate a statistically significant difference.
Results
Chemical structure of berberine
In brief, Figure 1A is the structure of berberine sketched by ChemDraw Professional 16.0.
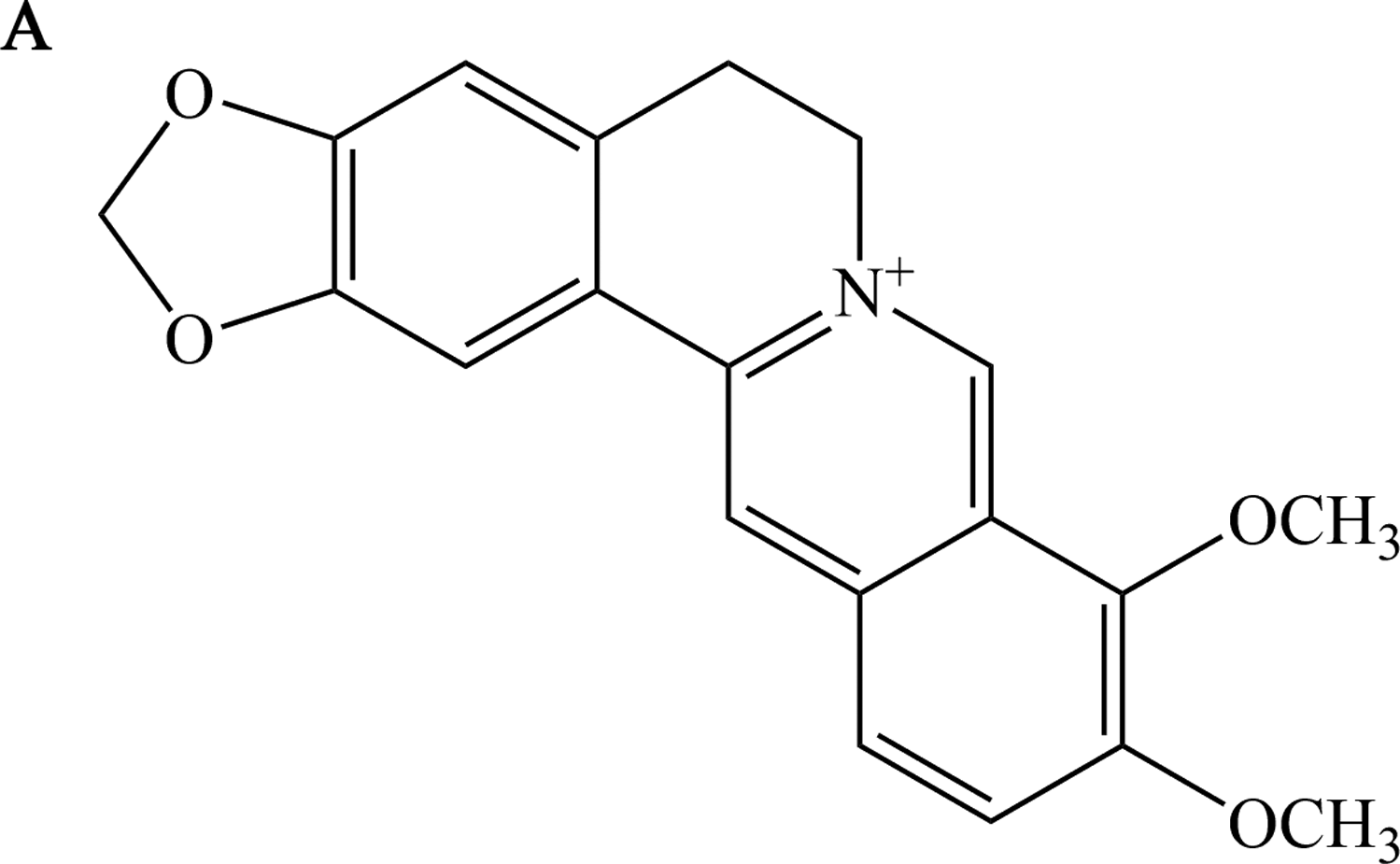
Chemical structure of berberine. Structure of berberine.
Effect of berberine on odontoblast differentiation of DPSCs
To determine the function of berberine in odontoblast differentiation of DPSCs, we detected the cell mineralization of DPSCs with various concentrations (0, 0.1, 1.0, 5, and 10 μg/mL) of berberine on day 14. There was more intense alizarin red S staining for the berberine-treated DPSCs versus the untreated control group (Fig. 2A). Meanwhile, the expression of odontoblast differentiation marker genes, including DMP1 and DSPP, was elevated by berberine (Fig. 2B, C). These data indicated that berberine might promote odontoblast differentiation of DPSCs. Furthermore, 5 μg/mL berberine could effectively enhance the expression of odontoblast differentiation markers. Therefore, 5 μg/mL was selected as the optimal concentration for subsequent experiments.

Effect of berberine on odontoblast differentiation of DPSCs. DPSCs were cultured in odontoblast differentiation medium and added berberine (0, 0.1, 1.0, 5, and 10 μg/mL) for 14 days.
Effect of berberine on β-catenin protein expression
It is well known that classical Wnt/β-catenin pathway plays a crucial role in promoting odontoblast differentiation of DPSCs (Song et al., 2017; Wang et al., 2018). Therefore, we detected the odontogenic effect of berberine on the production of β-catenin. To confirm the effect of berberine on the differentiation, we treated DPSCs with or without berberine during odontoblast differentiation. We found that berberine increased the expression levels of nuclear β-catenin by western blot (Fig. 3A–D). Immunofluorescence research also confirmed the same results (Fig. 3E).
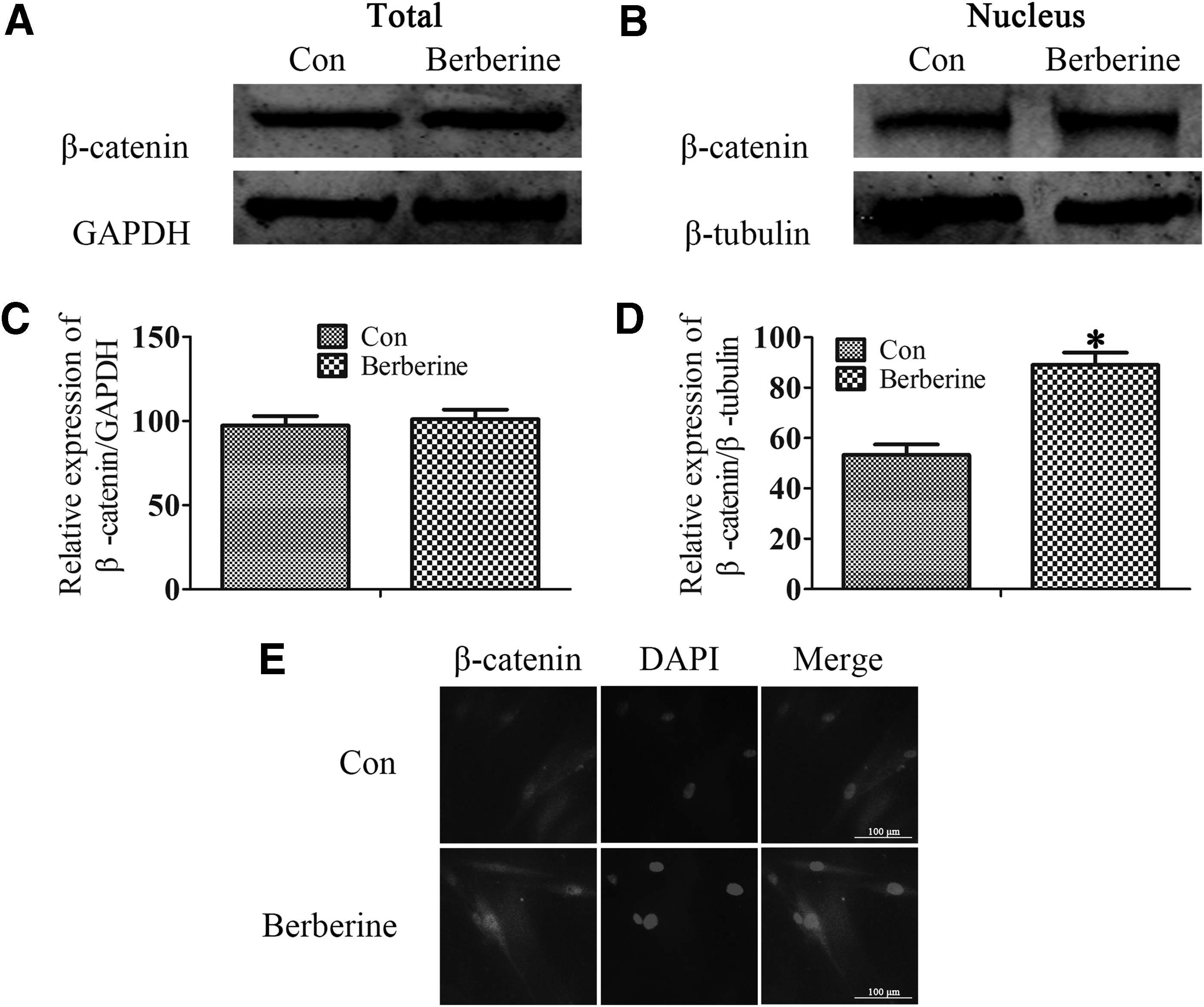
Effect of berberine on β-catenin protein expression. DPSCs were cultured in odontoblast differentiation medium and added berberine (5 μg/mL) for 14 days.
Canonical Wnt/β-catenin signaling is crucial for berberine-induced odontoblast differentiation of DPSCs
DKK-1 is one of the most well-characterized inhibitors of the Wnt/β-catenin pathway, preventing nuclear translocation of β-catenin (Song et al., 2017; Wang et al., 2018). DPSCs were treated with berberine and/or DKK-1 during odontoblast differentiation and the level of expression of β-catenin was examined by western blot analysis. As we expected, in the group treated with DKK-1 still suppressed nuclear β-catenin, even if the cells cultured berberine together (Fig. 4A–D). Immunofluorescence research also confirmed the same results (Fig. 4E). Taken together, these data indicated that the promotion of odontoblast differentiation by berberine was associated with activation of canonical Wnt/β-catenin signaling.
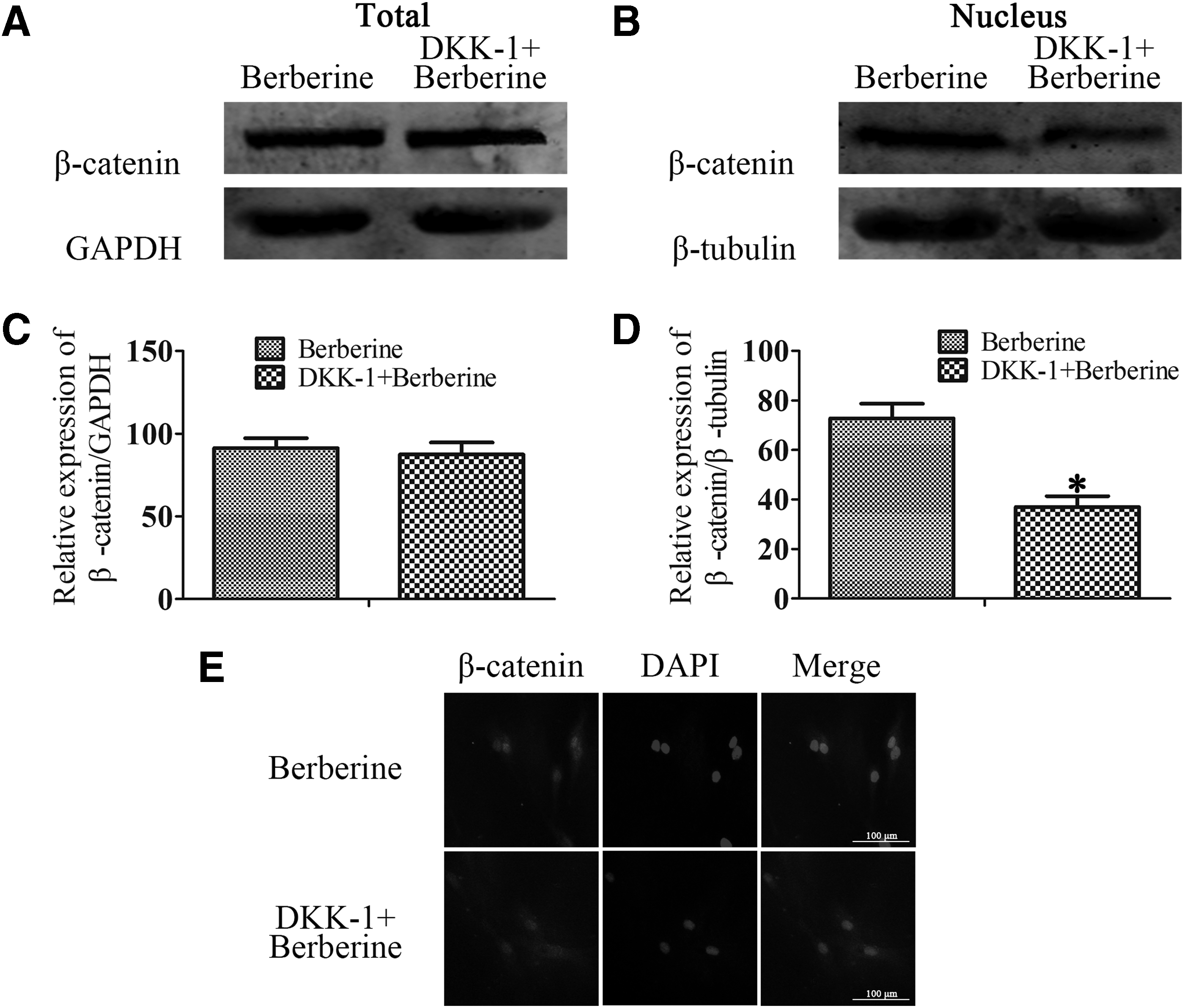
Canonical Wnt/β-catenin signaling is crucial for berberine-induced odontoblast differentiation of DPSCs. DPSCs were cultured in odontoblast differentiation medium and added berberine (5 μg/mL) or DKK-1 for 14 days.
Effect of berberine on downstream protein expression of Wnt/β-catenin signaling
Finally, a significant downregulation of the protein and mRNA levels of DMP1 and Runx2 could be detected when DPSCs were exposed to DKK-1 cocultured with berberine for 14 days in this experiment (Fig. 5A–C). These results further validated that berberine accelerated odontoblast differentiation by Wnt/β-catenin activation.
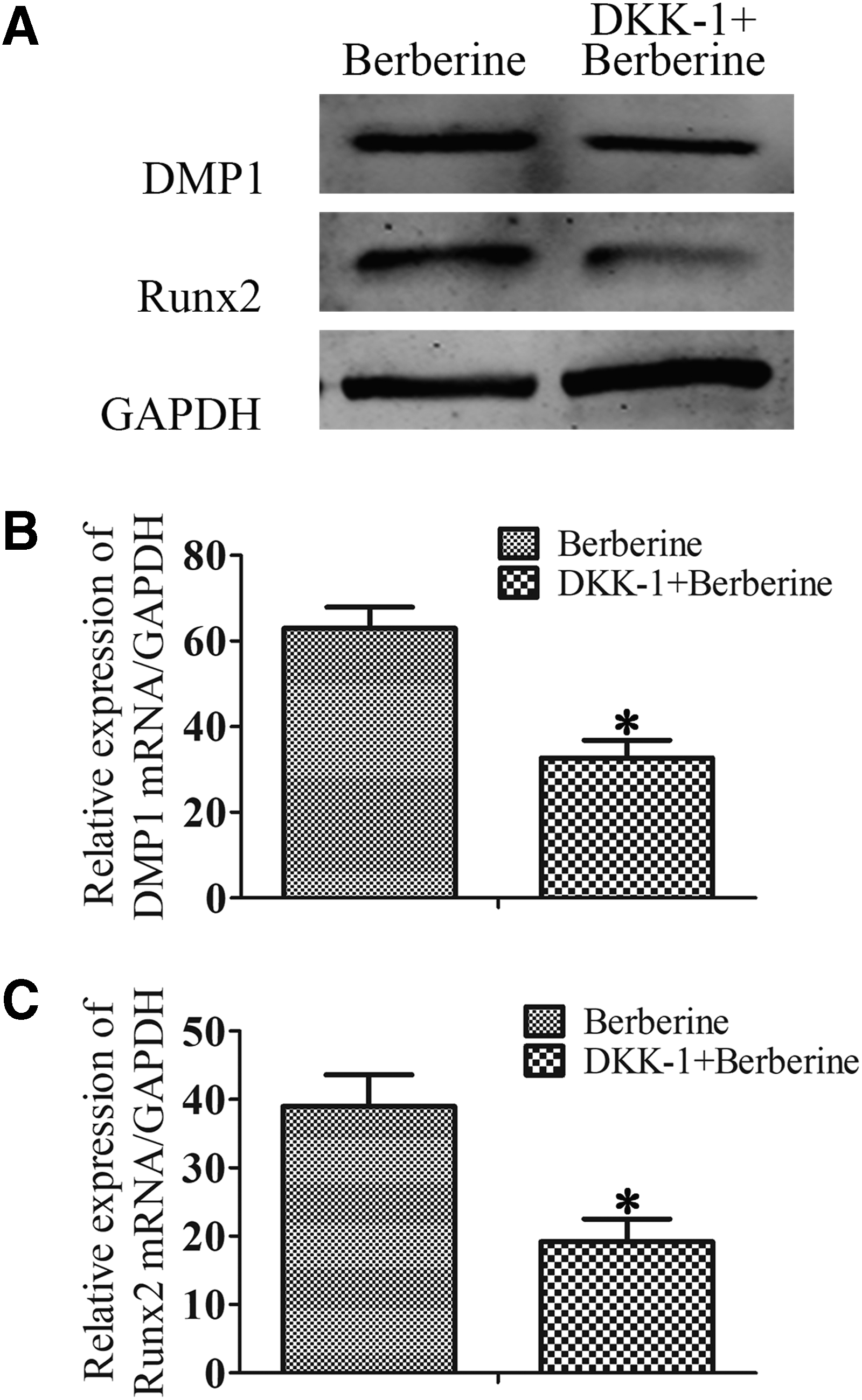
Effect of berberine on downstream protein expression of Wnt/β-catenin signaling. DPSCs were cultured in odontoblast differentiation medium and added berberine (5 μg/mL) or DKK-1 for 14 days.
Discussion
DPSCs possess the capability of regeneration and are thought to be the putative candidate for dental tissue engineering. DPSCs are ectodermal-derived stem cells, originating from migrating neural crest cells and possess MSCs properties (Nuti et al., 2016; Yan et al., 2011). These cells could differentiate into osteoblast, odontoblast, adipocyte, chondroblast, and neuron (Anitua et al., 2018). Among them, odontoblast differentiation of DPSCs controls the regenerative pulp-dentin complex tissues (Kawashima and Okiji, 2016). DMP1 is an extracellular matrix protein, which is responsible for differentiation of DPSCs into odontoblasts (Ching et al., 2017). DSPP is expressed by odontoblast-like cells underlying the reparative dentine (Da et al., 2018). Thus, DMP1 and DSPP have been used as two indicators of odontoblastic differentiation.
Berberine, as in Figure 1, is a nonbasic and quaternary benzylisoquinoline alkaloid, a relevant molecule in pharmacology and medicinal chemistry (Neag et al., 2018). It is a main bioactive substance used as a chemical marker for quality control of many prescriptions such as Huanglian-Jie-Du-Tang, Gegen-Huangqin-Huanglian-Tang, and Zuo-Jin-Wan, which have been used in clinical treatment for centuries (Wang et al., 2017). As we mentioned earlier, berberine possesses multiple pharmacological effects and its influence on the differentiation of DPSCs differs.
Jin Liu et al. found that berberine promoted human periodontal ligament stem cells osteogenesis in the early, middle, and late stages (Liu et al., 2018). Yan Zhou et al. indicated that berberine increased SNP-stimulated chondrocyte proliferation by promoting G1/S phase transition and synthesis of PCNA in cartilage through activation of Wnt/β-catenin signaling pathway (Zhou et al., 2016). Naveen et al. investigated that berberine inhibited cancer-stem cell properties, metastasis, and EMT in neuroblastoma cells through negative regulation of PI3/AKT and RAS-RAF-ERK MAPK signaling pathways (Naveen et al., 2016). According to our results, berberine accelerated the odontoblast differentiation of DPSCs and increased the expression of odontoblast differentiation marker proteins by classical Wnt/β-catenin pathway.
The Wnt family comprises 19 proteins divided into two main categories, namely the β-catenin-dependent canonical and the β-catenin-independent noncanonical Wnt pathways. The classical Wnt/β-catenin signaling has been definitely confirmed to control cell proliferation, differentiation, apoptosis, and migration. Moreover, it also regulates dental tissue development and stem cell self-renewal (Song et al., 2017; Wang et al., 2018). During the dental development, Wnt/β-catenin as a pro-odontogenic signal at tooth initiation and in cusp development (Liu and Millar, 2010).
As we know that stem cells are intrinsically destined to differentiate, it seems that Wnt signals could block the default step, the differentiation of cells, possibly by suppressing differentiation-specific genes (Clevers and Nusse, 2012). Runx2 is a transcriptional factor and known master regulator in controlling osteoblast and odontoblast differentiation. Evidence showed that activing Wnt/β-catenin could led to the activation of Runx2 and the expression of downstream genes (Han et al., 2014; Liu et al., 2016).
In this study, we cultured DPSCs from the caries-free teeth of patients, and then induced odontoblast differentiation of DPSCs. On day 14, alizarin red S and ALP staining of odontoblast differentiation of DPSCs revealed that berberine-treated groups were stronger than untreated groups. Figure 2 indicated that berberine promoted the expressions of DMP1 and DSPP, especially at the concentration of 5 μg/mL. Hence, berberine has a positive effect on odontoblast differentiation of DPSCs.
To investigate the relationship with berberine and Wnt/β-catenin pathway, DPSCs were treated with berberine and/or DKK-1 during odontoblast differentiation. As it shows in the Figures 3 and 4, berberine enhanced the expression levels of nuclear β-catenin and DKK-1 inhibited berberine-induced nuclear β-catenin. Furthermore, berberine activated the levels of DMP1 and Runx2, which were downstream protein of Wnt/β-catenin signaling (Fig. 6).
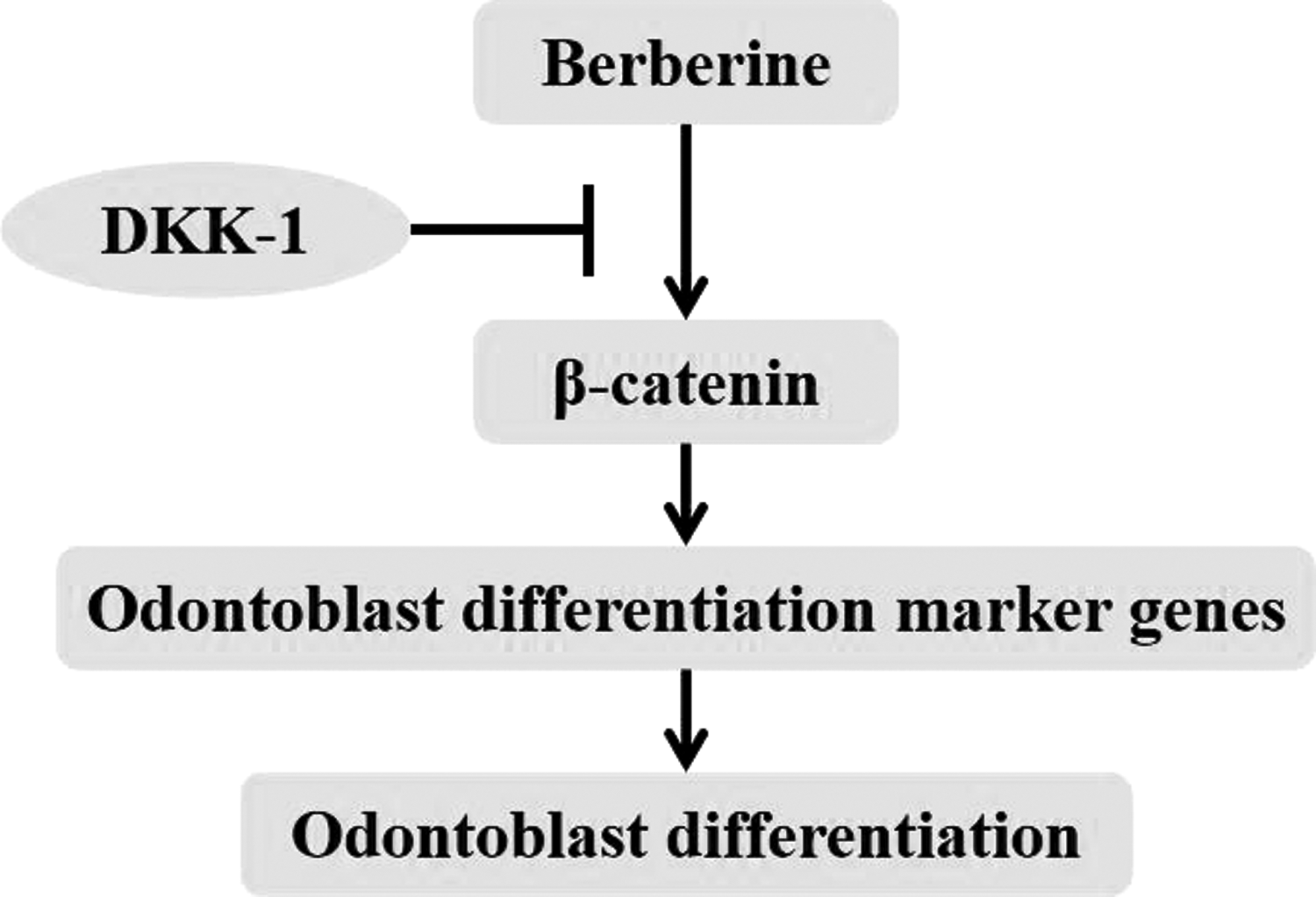
Schematic of berberine regulates odontoblast differentiation. Berberine activates classical Wnt/β-catenin pathway, increases the expressions of odontoblast differentiation marker proteins, and promotes the odontoblast differentiation of DPSCs.
In brief, berberine could be a novel medicine for the therapy of dental defect. Based on the complication of odontoblast differentiation of DPSCs, further research is necessary to determine whether berberine could regulate the odontoblast differentiation of DPSCs through other signaling pathways and whether berberine could affect the other pathological processes of dental defect.
Footnotes
Acknowledgments
The study was partly supported by Six Talent Peaks Project in Jiangsu Province (No. 2016-WSN-103) and Research Topic of Curriculum Reform in Nantong University (2018B55).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.