
Abstract
Abstract
Transcription factors NANOG, OCT4, SOX2, and NESTIN are expressed in both human embryonic stem cells (hESCs) and cancer stem cells and they play a crucial role in maintaining characteristics of stemness such as self-renewal and pluripotency. This article evaluates the expression of variants of the main stem cell-specific transcription factors NANOG and OCT4 critically and accurately with specific primers designed for identifying the most important variants that maintain stemness. We have examined four variants of NANOG along with a processed pseudogene and seven variants of OCT4 in human teratocarcinoma cell lines (NTERA2D1, SuSa, GCT-27, and 833KE), hESCs, and ovarian cancer cells by reverse transcriptase-polymerase chain reaction. In addition, we have examined their expression in NTERA2D1 cells on differentiation with all-trans-retinoic-acid. We show that NANOG1 is expressed in all teratocarcinoma cells and can be distinguished from NANOGP8, which is an expressed pseudogene. NANOG2 was not expressed in any of the cell lines, including ESCs. OCT4A was expressed in all cells, whereas the variant OCT4B-variant 3 was expressed only in NTERA2D1 cells. On differentiation of NTERA2D1 with retinoic acid, only NANOGP8 and OCT4A were expressed. In ovarian cancer cells, only 3/6 expressed NANOG1 and OCT4A. All malignant cells from patients with ovarian cancer (N = 6) expressed NANOG1 and OCT4A. These results demonstrate the necessity to precisely evaluate the expression of stem cell transcription factors when defining stemness.
Introduction
Unlimited undifferentiated propagation of cells by self-renewal and the potential to differentiate into specific cells of all three germ layers remain the most important property of embryonic stem cells (ESCs), referred to as “stemness” (Aponte and Caicedo, 2017). The maintenance of “stemness,” as opposed to differentiation in vitro, is strictly controlled. Functional assessment of stemness includes properties such as transplantation ability and hierarchy, supported by the expression of stem cell-specific transcription factors. The essential transcription factors to maintain a “stem cell state” or stemness in ESCs have been identified to be NANOG, OCT4, NESTIN, and SRY (Sex determining region Y)-box 2 or SOX2. The expression of OCT4 and SOX2 are equally important in the maintenance of iPSCs (Takahashi and Yamanaka, 2006).
Cancer stem cells (CSCs) or tumor initiating cells have also been known to express these transcription factors to a varying degree depending on the type of tumor. Increased expression of these stemness genes is also observed in CSCs from different tumor types, and it is essential for maintenance of the stem cell program (Chambers et al., 2003). The technique for studying their expression has been by immunofluorescence, reverse transcriptase-polymerase chain reaction (RT-PCR), and functional analysis at the protein level. The availability of human genome sequence and other comparative species has led to the identification of splice variants and pseudogenes of these factors. This is particularly important while evaluating the expression of NANOG and OCT4, as they have several variants unlike NESTIN or SOX2. It is still unclear which variants of these factors are expressed in human ESCs (hESCs) or CSCs.
Octamer binding transcription factor 4, also known as OCT4, OCT3/4, or POU5F1, is a master regulator of pluripotency and self-renewal in embryonic stem (ES) and embryonal carcinoma (EC) cells. It belongs to the family of transcription factors having a POU-DNA binding domain (Rosner et al., 1990). Other than ESCs, this factor has been detected in adult stem cells, including bone marrow-derived mesenchymal stem cells and malignant cells of various origins. The population of OCT4 positive cells in human cancers may represent CSCs, which are responsible for sustained growth and propagation of tumors (Webster et al., 2007). Knockdown of OCT4 results in differentiation of ES and EC cells (Hay et al., 2004). It is localized to chromosome 6p, in the region of major histocompatibility complex.
OCT4 has been shown to encode three types of splice variants originally described as OCT4A, OCT4B, and OCT4B1 (Atlasi et al., 2008). Recently, several other variants of OCT4B have been identified: OCT4B-variant 2/5, OCT4B-variant 5, OCT4B2, OCT4B3, and OCT4B4 (Miyamoto et al., 2018; Poursani et al., 2017a, 2017b, 2017c). OCT4A is primarily responsible for maintaining the pluripotency and self-renewal properties of ESCs and is localized in the nucleus, whereas OCT4B is expressed in the cytoplasm of cancer cells. It is induced on cell stress and lacks any major role in sustaining stemness. High expression of OCT4B1 and OCT4B4 in ES/EC cells and their downregulation on differentiation suggest a vital role of these variants in stem cells (Atlasi et al., 2008; Miyamoto et al., 2018).
Knockdown of OCTB4 using shRNA resulted in the enhanced accumulation of EC cells (NCCIT) in the G0/G1 phase of the cell cycle in comparison to the control (Eini et al., 2014). In most of the studies, expression of OCT4A is evaluated as a marker of stemness rather than OCT4B1. Moreover, the main transcribed pseudogenes of OCT4 resemble OCT4A to a higher extent than OCT4B or OCT4B1 (Suo et al., 2005). The analysis of expression of all variants alone will determine which one is indispensable for the maintenance of stemness in ESCs.
NANOG gene, a part of the “core NANOG network” is a homeodomain transcription factor that plays a vital role in maintaining stemness in ESCs along with OCT4 and SOX2 (Chambers et al., 2003; Mitsui et al., 2003). The presence of 10 pseudogenes and various alternatively splice variants hinder accurate detection of NANOG as a marker of stemness (Eberle et al., 2010). Previous reports have shown that, among all the pseudogenes and splice variants, only transcripts of NANOG1, NANOG2, and NANOGP8 are translated to proteins as the other pseudogenes do not exhibit intact open reading frames (Fairbanks and Maughan, 2006).
Another key pluripotency marker, SOX2, despite being an intronless gene, has been shown to express SOX2OTS1 and SOX2OTS2 as splice variants, which lie embedded in the third intron of an lncRNA gene, termed as SOX2 overlapping transcript (SOX2OTS) (Shahryari et al., 2014). Similarly, NESTIN, a type IV intermediate filamental protein, is an essential marker of pluripotency in stem cells, especially in neural stem cells (Lendahl et al., 1990). In humans, splice variants of NESTIN have not been described. However, a transcript variant Nes-S was observed in the neurons of dorsal root ganglia of adult rats (Su et al., 2013).
In this article, we have evaluated the expression at the mRNA level of transcription factors OCT4, NANOG and their variants in four teratoma cell lines and a hESC line. Testicular germ cell tumor cell lines have similar gene expression patterns to hESCs and, thus, serve as an excellent prototype model of ESCs (Andrews, 2002). They aid in substantiating the expression of the variants that are necessary to maintain stemness.
Methods
Primary samples and cell culture
Malignant ascites from untreated patients (N = 6) diagnosed with high-grade serous ovarian adenocarcinoma (HGSOC) were collected for this study with informed consent. This study was approved by the Institutional Ethical Committee at Cancer Institute, Chennai. Samples of ascites containing malignant cells from patients with HGSOC confirmed by both ascitic fluid cytology and true-cut biopsy were chosen for the study. Samples were centrifuged at 3000 rpm for 15 minutes to pellet down the cells. The red blood cells (RBCs) were later eliminated by treating the cells with RBC lysis buffer for 10 minutes. The cells were plated in the supernatant of the ascites obtained after centrifugation.
The ovarian cancer cell lines were obtained from American Type Tissue Culture Collection (ATCC) (Manassas, VA, www.atcc.org) (Bignone et al., 2007). Testicular cell lines 833KE, GCT27, and SuSa were obtained from Dr. Beate Koberle, Mainz, Germany and NTERA2D1 was a kind gift from Dr. Maneesha S. Inamdar (Jawaharlal Nehru Centre for Advanced Scientific Research, Bangalore, India) (Inamdar et al., 2009; Koberle et al., 1997). mRNA from hESCs was a kind gift from Dr. M. Panicker (National Centre for Biological Sciences, Bangalore, India) (Muthusamy et al., 2014).
All cells were maintained in recommended growth medium supplemented with 10% fetal bovine serum (Invitrogen), 2 mM
Preparation of egg white layer for 3D culture
The preparation of egg white was performed according to the protocol previously described (Kaipparettu et al., 2008). Briefly, freshly obtained avian eggs were surface sterilized with 70% ethanol and a 5-cm window was cut open by using a scalpel. The egg white was carefully collected in a sterile tube without disturbing the yolk. One milliliter of the egg white was added per well of a six-well tissue culture dish and heated at 60°C for 20 minutes. Special care was taken to ensure that the egg white was not over heated and opaque.
Spheroid assay
Cells isolated from malignant ascites or cell lines were cultured in serum-free stem cell medium containing DMEM-F12, 20 ng/mL basic fibroblast growth factor, 10 ng/mL epidermal growth factor, and 1 × insulin transferrin selenium. The number of cells plated varied according to the size of the dish; for example, in a six-well dish, 104 cells were added on a layer of semisolid egg white.
In silico analysis and designing of primers
We have identified, by a composite approach, four variants of NANOG and seven variants of OCT4. The GenBank numbers of these genes are: NANOG1 (NM_024865.2), NANOG2 (AY455283.1), NESTIN (NM_006617.1), SOX2 (NM_003106.2), and OCT4 (NM_002701.4). Specific exon spanning primers were designed for all the variants by using the software Primer3 plus and confirmed by an in silico PCR (Serial cloner2.1) (Table 1).
Primers Designed for Detecting Specific Variants of Stem Cell Transcription Factors
Total RNA extraction, cDNA synthesis, and RT-PCR
Total RNA extraction was performed by using Trizol (Invitrogen) reagent following the manufacturer's instructions. The quantification of the extracted RNA was done by spectrophotometry and confirmed by electrophoresis on a 1% agarose gel. The first cDNA strand was synthesized in a 20-μL reaction volume with M-MuLV reverse transcriptase (NEB), using total RNA (1 μg) as a template and oligo (dT) as a primer. For RT-PCR, the cDNA template (2 μL) was used along with Taq DNA polymerase (New England Biolabs) with a total reaction volume of 25 μL. For each sample, controls without reverse transcriptase (No-RT) control and DNA were also used along with RNA to detect any nonspecific amplification of genomic DNA.
In addition, beta-actin was used as the positive control to confirm the integrity of RNA. The amplifications were performed after initial denaturation at 94°C for 5 minutes followed by 35 cycles with 94°C for 45 seconds, 54–60°C for 45 seconds, and 72°C for 45 seconds, with an additional step of final extension at 72°C for 10 minutes. A gel documentation system (Bio-Rad) was used to capture digital images of PCR amplicons, which were run on a 2% agarose gel. The identity of PCR amplified targets was confirmed by sequencing reactions using Big Dye Terminator v1.1 Cycle Sequencing Kit (Applied Biosystems) according to the manufacturer's instructions (ABIPRISM 3100 Genetic Analyzer; Applied Biosystems). Sequences were analyzed by using Sequencher 5.4 software.
Differentiation of NTERA2D1 cells
For neural differentiation, NTERA2D1 cells were seeded at a density of 5 × 105/well of a six-well culture dish in medium containing 10−5 M all-trans retinoic acid (ATRA) diluted from stock solution of l0−2 M in dimethyl sulfoxide (DMSO). The cells were fed with fresh medium containing retinoic acid weekly. After 3 weeks, the cells were harvested by using 0.25% trypsin and 2 mM EDTA. The cells were reseeded at 105 cells/well of a six-well culture dish, and they were cultured for a period of 3–4 weeks in the absence of ATRA. The differentiated neuronal cells were observed after 14–17 days of exposure to ATRA.
Immunofluorescence
Expression of neurotrophic tyrosine kinase, receptor, type 1 (TRK A), and glial fibrillary acidic protein (GFAP) was evaluated in NTERA2D1 cells by immunofluorescence (IF) using specific mouse antihuman antibodies (Santa Cruz Biotechnologies) and FITC/PE as a secondary antibody (Santa Cruz Biotechnologies). Briefly, cells were grown overnight on coverslips in DMEM and 10% FBS and washed thrice with PBS. This was followed by fixation of cells with methanol and treatment with 1.5% BSA in PBS for blocking nonspecific fluorescence. Cells were incubated with primary antibody (1:50) for 60 minutes followed by secondary antibody (1:100) for 120 minutes. DAPI was used as the counterstain. Coverslips were mounted on glass slides and observed under a fluorescence microscope (Axio Imager.A2; Carl Zeiss).
Results
Identification of splice variants in teratocarcinoma cell lines
Various splice variants and pseudogenes have been identified for OCT4 and NANOG (Booth and Holland, 2004; Eberle et al., 2010; Poursani et al., 2017a). The reference sequences from NCBI for all the four genes were used to derive accurate intron-exon structures in comparison with genomic sequence. In addition, public databases such as NCBI and ENSEMBL were used to evaluate all splice variants. Using this composite approach, we have identified four potential variants of NANOG and seven variants of OCT4 (Fig. 1A, B).

Splice variants of NANOG and OCT4. The exons of NANOG
Isoforms of NANOG
We identified four splice variants of NANOG based on literature and a comparison between redundant human genomic and transcript databases from NCBI (Fig. 1A, B). In addition to NANOG1, NANOG2, and pseudogene NANOGP8, we identified transcripts with alternate upstream exons. We have labeled them as NANOG3 and 4. This was based on an analysis of all mRNA sequences against the genomic databases (Das et al., 2012; Eberle et al., 2010). To determine and analyze these variants in human cancer cell lines, we designed specific primer combinations for each one of them as shown in Figure 1A and B. The main variants of NANOG (NM_024865.2, NANOG1) and NANOG Pseudogene (LOC100293888, NANOG P8) are almost similar except for the 22 bp (1685–1707 bp) sequence present exclusively in exon 5 of NANOGP8 (Fairbanks et al., 2012; Vaidya et al., 2018).
Therefore, the reverse primer was specifically designed in this region so as to amplify only NANOG1 excluding the pseudogene. Amplification of both sets of primers implies that NANOG1 is expressed. Alternatively, if only one primer (NANOG FP2/RP2) that is commonly used detects a transcript, then the pseudogene alone is expressed. This strategy was adopted to exclude the occurrence of false positive expression of NANOG1 (Table 1). It has been previously reported that ESCs, iPSCs, and CSCs express NANOG1, as a marker of stemness (Das et al., 2012). However the primers designed in exon 1, exon 2, or exon 3 amplify both NANOG1 and NANOGP8, giving an erroneous result that NANOG1 is expressed. To circumvent this issue, specific primer sets were designed that can exclude the expression of the pseudogene from the main variant, NANOG1.
We examined a series of teratocarcinoma cell lines (N = 4) for the expression of all stemness genes and their variants. hESC line was used as a positive control (Fig. 2). To eliminate nonspecific amplifications, no-RT and no cDNA controls were used (data not shown). The main variant (NANOG1, NM_024865.2) was present in all the cell lines, as products specific for both NANOG pseudogene and NANOG1 were detected. The transcript, NANOG3 was detected in all cell lines whereas NANOG4 was observed only in three cell lines examined. GCT27 did not express NANOG4. The primers designed for amplifying NANOG4 also amplified NANOG3 (data not shown) (Eberle et al., 2010). NANOG2 (AY455283.1) was not expressed in any of the cell lines, including ESCs. All the transcripts were purified, and sequencing (BigDye v.3.1 ABI 377) confirmed the identity of the PCR products (data not shown).
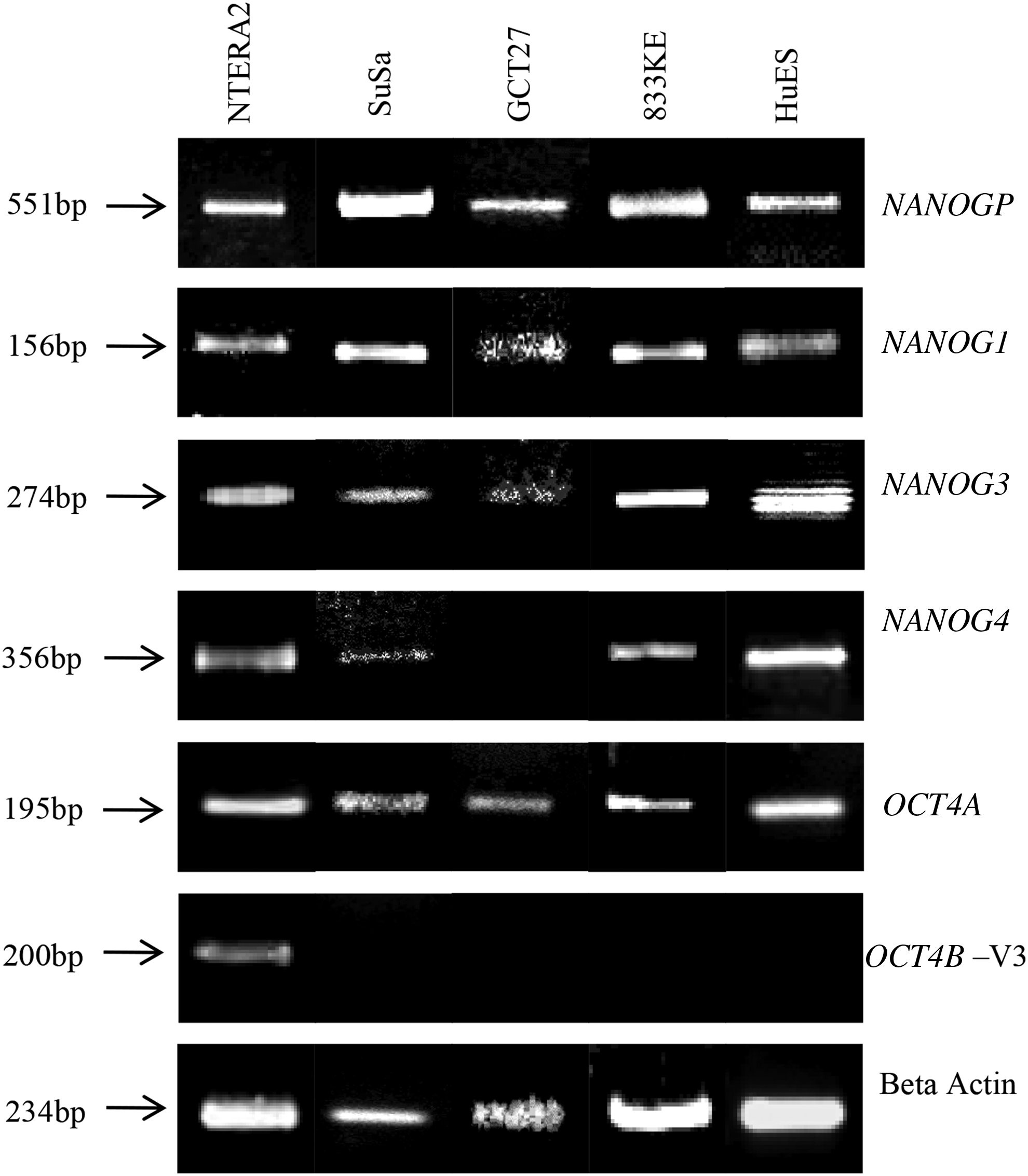
Expression of the NANOG and OCT4 variants. RT-PCR showing expression of the NANOG and OCT4 variants. The variants of NANOG (NANOG1, NANOG3, NANOG4, and NANOGP) (top panel) and OCT4 (Oct4B-V3) (bottom panel) were analyzed in the teratocarcinoma cell lines (SuSa, GCT27, 833KE, and NTERA2D1) and hESC line. Beta-actin (ACTB) was used as the positive control. The image is a composite of different RT-PCR experiments. hESC, human embryonic stem cell; RT-PCR, reverse transcriptase-polymerase chain reaction.
Isoforms of OCT4
The expression of OCT4 isoforms was evaluated in teratocarcinoma cell lines (N = 4) with hESCs as a positive control (Fig. 2). All the primers were designed to amplify the variants that were essential for stemness (OCT4A, OCT4B-variant 2/5, OCT4B-variant 3, OCT4B1, and OCT4B4) excluding the possibility of amplifying all pseudogenes known till date. We have not evaluated the expression of OCT4B2 and OCT4B3. Expression of the main isoform OCT4A (Fig. 2), previously shown to be necessary for maintenance of “stemness” in cells, was found to be present by RT-PCR in all the cell lines. Variant OCT4B1, previously shown to have a critical role in maintaining the stemness and pluripotency state of hESCs, was not expressed in any of the cell lines (Atlasi et al., 2008) (Figs. 1B and 2).
Similarly, OCT4B-variant 2/5, believed to be indispensable, was not expressed in any of the cell lines, including the hESC line. However, OCT4B-variant 3 was expressed exclusively in NTERA2D1 cells (Fig. 2). Surprisingly, the recently identified novel splice variant OCT4B4 could not be detected in any of the cell lines.
Other transcription factors that play a key role in maintaining stemness include SOX2 and NESTIN. Expression of NESTIN and SOX2 was observed in all cell lines (data not shown).
Differentiation of NTERA2D1 cells
The cell line NTERA2 was grown in the presence of retinoic acid for 2 weeks to allow for differentiation to occur, as previously shown (Eini et al., 2014). Cells differentiated to form neuronal cells and expressed both GFAP and Trk A receptor, confirming their neuronal identity (Fig. 3A). GFAP is an intermediate filament protein expressed in the glial cells known as astrocytes and TRK A is a neurotrophic high-affinity receptor for NGF, both of which are used as neural markers for identifying progenitor cells (Casper and McCarthy, 2006).
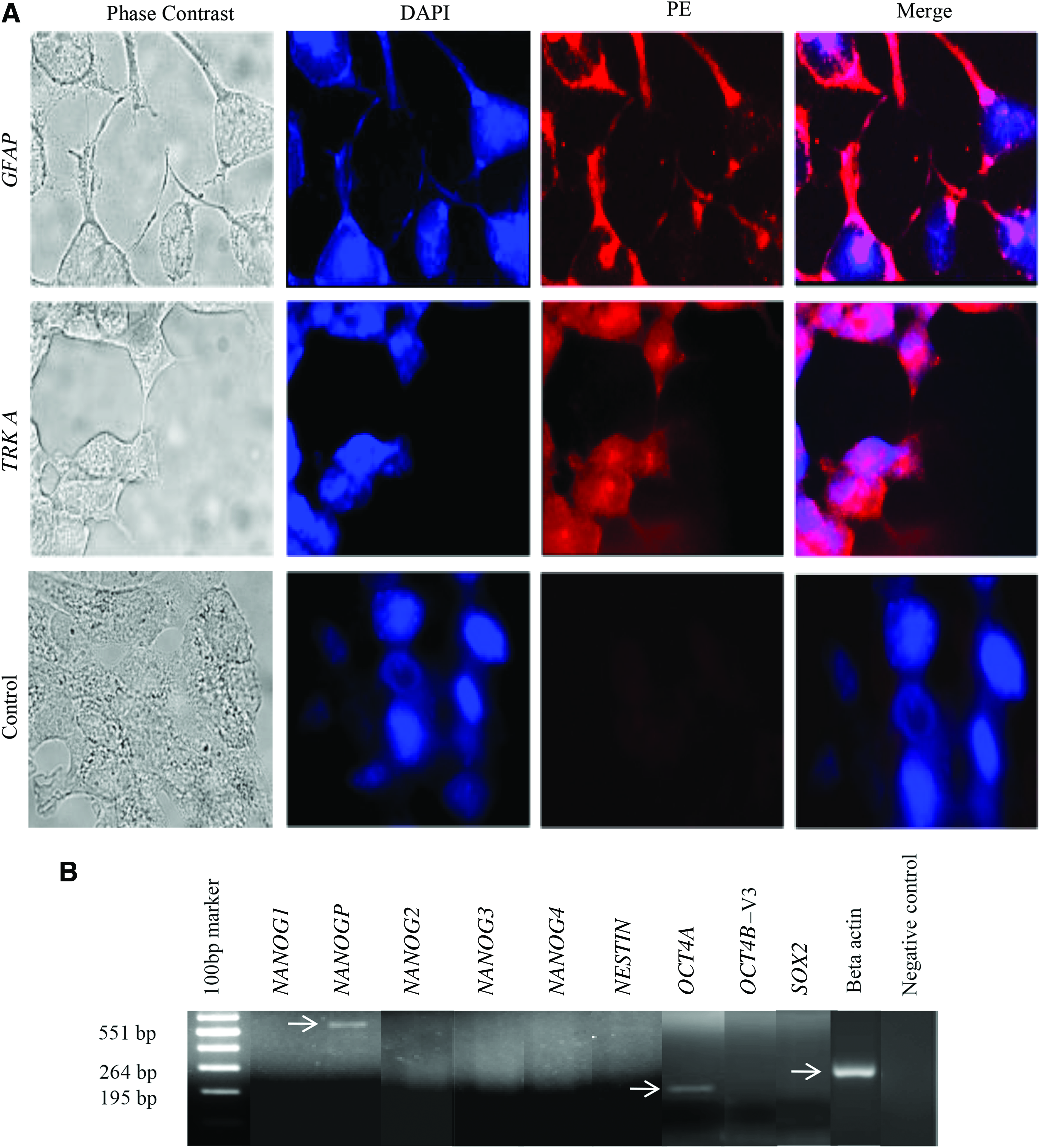
Immunofluorescence image showing differentiation of NTERA2D1 cells with retinoic acid to neuronal lineage.
RNA was extracted from these differentiated cells to assess which isoform of NANOG and OCT is expressed in these cells. Only NANOGP8 pseudogene and OCT4A were expressed in these cells. This suggests that all the variants of stem cell-specific genes, NANOG, OCT4, SOX2, and NESTIN, at least by RT-PCR, were repressed on differentiation of NTERA2 cells. Only the pseudogene for NANOG and the main variant of OCT4 (OCT4A) were expressed (Fig. 3B). This information is vital with respect to genes required to be analyzed for stem cell attributes. The expression of all the genes was assessed only on terminal differentiation at 2 weeks. Evaluation at intermediate time points was not performed.
Analysis of splice variants of stem cell factors in ovarian cancer cells, monolayer and spheroids
The transcripts of NANOG, OCT4, SOX2, and NESTIN were analyzed in ovarian cancer cell lines (N = 6) and malignant ascites (N = 6). The cell lines were OVCAR3, OVCAR5, PEO4, UCI101, OAW28, and 41M. NANOG1 was expressed in the cell lines UCI101, OAW28, and 41M. The remaining cell lines did not express or expressed only the pseudogene for NANOG1. NANOG2 and NANOG4 were not detected in any of the cell lines; whereas NANOG3 was expressed only in 41M cells. Among the variants of OCT4, OCT4A was expressed in all cell lines except OVCAR3, OAW28, and PEO4. Apart from OVCAR3 and UCI101, SOX2 was expressed in all other cell lines. NESTIN was expressed only in UCI101 cells (Table 2).
Expression of Splice Variants of Nanog and Oct 3/4 in Ovarian Cancer Cell Lines and Spheroids
NP8, NANOG pseudogene; N1, NANOG1; N2, NANOG2; N3, NANOG3; N4, NANOG4; V1, OCT3/4 Variant 1; V2/5, OCT3/4 Variant 2/5; V3, OCT3/4 Variant 3; B1, OCT4B1; B4, OCT4B4.
To compare the expression of these genes in 3D culture, cell lines (OVCAR3, OVCAR5, and PEO4) grown as spheroids on egg white were analyzed by RT-PCR. Spheroids are enriched for CSCs and have some features of stem cells. In ovarian cancer, they have been used in the absence of specific surface markers, to study CSCs (Zhang et al., 2017). We have used cells plated on egg white as a substrate to grow spheroids (Nagare et al., 2015). Interestingly, spheroids from OVCAR5 cells showed the expression of the main variant of NANOG (NANOG1) that was absent in monolayer culture. NANOG3 and NANOG4 were expressed in the spheroids obtained from OVCAR3 and OVCAR5 cells (Fig. 4). The main variant of OCT4 (OCT4A) and NESTIN was expressed in spheroids from OVCAR5 cells.

Expression of splice variants of NANOG in OVCAR3. RT-PCR of OVCAR3 cells with specific primers in monolayer and spheroids detected the expression of the variants of NANOG. The image is a compilation of different RT-PCR experiments. Left and right panel shows the expression of variants in OVCAR3 cells grown as monolayer and spheroids, respectively. 1. NANOG pseudogene (551 bp). 2. NANOG3 and NANOG4 (356 and 195 bp). 3. 100 bp ladder. 4. Beta actin (264 bp). *Negative control.
The expression of stem cell genes was also evaluated in primary malignant cells from patient ascites (N = 6). NANOG1 and OCT4A were present in all the samples. Other variants of NANOG and OCT4 could not be detected in any of the samples except NANOG3. SOX2 was expressed in five samples (83%), and NESTIN was present in only four samples (66%) (Table 3).
Expression of Splice Variants of Nanog and Oct 3/4 in Cells from Malignant Ascites
Discussion
This article demonstrates that the NANOG gene identified by many other laboratories as a primary marker of stemness could have been confused with expression of the pseudogene (NANOGP8). Unless evaluated by specific primers, the expression of NANOG cannot be interpreted accurately. The homeodomain containing transcription factor NANOG has been reported to have 11 pseudogenes till date (Booth and Holland, 2004). Among these pseudogenes, only NANOG homeobox pseudogene 8 (NANOGP8) could be transcribed and translated to a functional protein due to the presence of a complete open reading frame (Fairbanks et al., 2012). The protein translated from the pseudogene differs from the embryonic NANOG protein only by a single amino acid change from Gln-253 in NANOG to His-253 in NANOGP8 (Ibrahim et al., 2012).
The functional role and expression pattern of NANOG and NANOGP8 in human cancers still remain elusive. In prostate cancer, it was shown by molecular and functional analysis that NANOG mRNA found in the cancer tissue was contributed mainly by the NANOGP8 transcript, whereas expression of NANOG was confined only to the nuclei of tumor cells (Jeter et al., 2009). Similarly, in colon cancer, expression of NANOG protein collectively resulted from transcription of both NANOG and NANOGP8 genes (Ibrahim et al., 2012). Majority of the published literature have reported NANOG as a CSC-specific transcription factor using primers that can amplify both NANOG and NANOGP8. This does not clarify whether the expression of NANOG in these cells truly reflects stemness or a false expression signal due to the pseudogene.
However, there is now reasonable evidence to suggest that NANOGP8 may also have a role in maintaining stemness (Zhang et al., 2013). Further, it is expressed in malignant cells from gastric cancer, prostate cancer, and leukemia (Palla et al., 2014). One can speculate that both NANOG1 and NANOGP8 have a role in maintaining stem cells but the specific roles of each gene are still unclear.
Although the expression of OCT4 has been considered a hallmark of ES and EC cells, there are studies that show that the expression of this gene is not particularly restricted to progenitor or stem cell compartments alone (Dangles-Marie et al., 2007). OCT4 has also been shown to be expressed in cancer cell lines, in addition to normal and tumor cells (Poursani et al., 2016). The presence of multiple isoforms and pseudogenes of OCT4 gene makes it difficult to interpret the data acquired by RT-PCR and quantitative real-time PCR.
This is mainly due to the absence of specific discriminating primers. OCT4 has two possible promoters that give rise to two groups of variants: OCT4A and OCT4B (OCT4B-variant 2, OCT4B-variant 3, OCT4B-variant 5, OCT4B2, OCT4B3, and OCT4B4). OCT4A, which is responsible for maintaining the stem cell properties of ES, EC cells, and pluripotent primordial germ cells, remains the principal variant of OCT4 (Nichols et al., 1998). The OCT4B group of variants mainly play a critical role in determining the response of cells to heat stress conditions and are expressed in differentiated cells along with some of the cancer cells (Gao et al., 2012). OCT4B transcript translates into three different protein isoforms (OCT4B-265, OCT4B-190, and OCT4B-164) by alternative translation initiation (Wang and Dai, 2010).
The results obtained in our study showed that neither NANOG2 nor OCT4B1 was expressed in any of the teratocarcinoma cell lines examined, which is different from the previous results in the literature (Atlasi et al., 2008; Poursani et al., 2017a). Moreover the observation that the main variant of OCT4A was expressed in all the teratoma cell lines as well as the differentiated NTER2D1 cells questions the importance of this gene in the maintenance of ESC state. In contrast, the presence of OCT4B-variant 3 only in NTERA2D1 cells that harbor the ability of self-renewal and pluripotency may suggest the possibility of expression of this specific variant at a later stage of development, explaining its lack of expression in hESCs.
Further, the absence of all stem cell gene variants except OCT4A and NANOGP8 in differentiated NTERA2D1 cells revealed the relevance of examining the expression of these genes to determine stemness. It is known that OCT4A can be expressed in differentiated cells (Zangrossi et al., 2007).
NANOG3 and NANOG4 were differentially expressed in spheroids from OVCAR3 and OVCAR5 cells in comparison to monolayer. Spheroids are enriched in CSCs (Nagare et al., 2015). This result indicates that transcript variants NANOG3 and NANOG4 might be preferentially expressed in CSCs. In malignant cells from ascites, the expression in the majority of the samples of the four main stem cell transcription factors suggests that there is a recapitulation of the stem cell phenotype. However, none of the other variants was expressed. To address which variant is expressed in CSCs in malignant cells from ascites, additional experiments to sort them have to be performed. This can be performed by using functional assays such as ALDH1A1 expression.
To our knowledge, this study is the first to report the expression of variants of the main stem cell-specific transcription factors critically and accurately with specific primers designed for identifying the most important variants that maintain stemness. The results present in this article address the results of expression by qualitative PCR of stem cell transcription factors alone in teratocarcinoma cells. Additional experiments specifically with the various isoforms of NANOG and OCT4 have to be performed to understand which variant is required to maintain the stem cell state. These may require the use of siRNAs targeting these isoforms.
However, this may be challenging as there is probably some redundancy within the system as cells express many of the isoforms. As majority of the reports examine the expression of these factors to document the stemness of cells by RT-PCR or immunofluorescence (IF) alone, these results will help in more precise inference. The development of isoform-specific antibodies will also help in better delineation of expression in ESCs.
Footnotes
Acknowledgments
The authors acknowledge the Department of Biotechnology (DBT), University Grants Commission (UGC), and Indian Council for Medical Research (ICMR) for financial support.
Authors' Contributions
S.S. performed the experiments and wrote the article. R.P.N. assisted with the experiments. P.M., S.V., and A.S. provided experimental suggestions and analyzed the data. T.S.G. conceived the study, supervised the findings of this work, and was in charge of overall direction and planning. He has reviewed, edited all versions of the article and approved the final version to be published.
Note Added in Proof
The expression of NANOGP8 has to be verified by also evaluating at the genomic level to distinguish it from NANOG1. The expression of NANOGP8 may be aberrant in ES as compared to malignant cells. The presence of widespread polymorphisms in NANOG and NANOGP8 can lead to an erroneous interpretation of reference sequences of variants (Vaidya et al., ![]() ).
).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.