
Abstract
Abstract
microRNA-29b (miR-29b) plays an important role in controlling DNA methylation in cells. We investigated its role during early embryonic development in buffalo embryos produced by somatic cell nuclear transfer (SCNT) and in vitro fertilization (IVF). miR-29b expression was highest at the 2-cell stage, decreased (p < 0.001) at the 4-cell stage, and remained low thereafter at the 8-cell, morula, and blastocyst stages, showing a similar pattern in cloned and IVF embryos. Treatment of reconstructed embryos with miR-29b mimic for 1 hour after 1 hour of electrofusion increased (p < 0.05) the total cell number and decreased (p < 0.05) the levels of apoptosis and DNA methylation compared with controls. It also increased (p < 0.05) the ratio of inner cell mass:trophectoderm cell numbers of blastocysts compared with controls to the levels observed in IVF blastocysts. However, the blastocyst rate was not affected by treatment with miR-29b mimic (29.0% ± 2.0% vs. 27.0% ± 2.0% for controls). The treatment decreased (p < 0.001) the expression of epigenetic-related genes, DNMT3A and DNMT3B, but not DNMT1, and increased (p < 0.05) that of pluripotency- (NANOG, OCT4, and SOX2) and development-related genes (FGF4 and GLUT1) in blastocysts compared with controls. Our results suggest that miR-29b mimic treatment of reconstructed embryos improves the quality, reduces the level of apoptosis and DNA methylation, and changes gene expression in SCNT blastocysts without affecting the blastocyst rate.
Introduction
Somatic cell nuclear transfer (SCNT) is carried out by transferring the nucleus of a differentiated somatic cell to the cytoplasm of an enucleated oocyte, which following nuclear reprogramming, converts it to the totipotent state. Cloning by SCNT has been successfully applied for producing live offspring to many mammalian species. Due to a large number of applications, SCNT has emerged as a very important reproductive technology in the last decade. It enables multiplication of high genetic merit farm animals much faster than what can be achieved through conventional breeding programs. In addition to this, SCNT has been successfully used for producing embryonic stem cells and transgenic animals, xenotransplantation and disease modeling, and conservation and restoration of endangered species (Niemann and Lucas-Hahn, 2012).
Due to low conception rate and very high mortality during postimplantation development in vivo, cloned embryos offer a very low live birth rate thus limiting its application on a large scale. In cattle, the live birth rate obtained with cloned embryos is only 9% (Panarace et al., 2007), whereas across all the mammalian species, the live offspring rate is only 0.9%–6% (Loi et al., 2011). The low birth rate is primarily attributed to the epigenetic abnormalities resulting from aberrant reprogramming of the donor nucleus to the totipotent state after nucleus transfer. The epigenetic abnormalities observed in cloned embryos include altered patterns of X-chromosome inactivation, imprinted gene expression, and abnormally high levels of genomic DNA methylation, which suggests that the epigenome is not established in a correct manner (Niemann et al., 2008).
Epigenetic reprogramming is an important aspect of embryonic development. Epigenetic changes are reversible forms of gene regulation which are heritable but are not dependent on the sequence of DNA. Methylation of DNA involves addition of a methyl group to the nucleotide cytosine, the DNA, predominantly in DNA regions known as cytosine–phosphate–guanine (CpG) islands. This is one of the most important epigenetic changes associated primarily with gene silencing (Wu and Morris, 2001). DNA methylation is modulated by specific enzymes called DNA methyltransferases (DNMTs), which include DNMT1, DNMT3a, and DNMT3b. DNMT1, which is responsible for adding methyl groups to the unmethylated daughter strands of newly replicated DNA, acts as a maintenance methyltransferase.
Methylation of CpG residues by DNMT1 results in the recruitment of methyl-binding domain protein and histone deacetylases, which leads to inhibition of transcription and silencing of the gene. Following implantation, the de novo methylation is carried out by DNMT3a and DNMT3b, which are expressed at a high level in developing embryos. The enzymes establish DNA methylation during embryonic development by adding new methyl groups to previously unmethylated DNA (Bestor, 2000).
MicroRNAs (miRNAs) and small interfering RNAs (siRNAs), which are a class of small noncoding RNAs, form another important component of epigenetic changes. miRNAs are 20–25 nucleotides in length, RNA molecules which abrogate abundance of target messenger RNAs (mRNAs) through inhibition of translation or a decrease in the abundance of the target transcripts (Bartel, 2009). miRNAs are implicated in the regulation of several very important biological and developmental events in reproduction, including oocyte maturation, fertilization, and embryo development (Gilchrist et al., 2016; Hossain et al., 2012), maternal-to-embryonic transition (Mondou et al., 2012), and nuclear reprogramming (Onder and Daley, 2011).
Several studies indicate that microRNA-29b (miR-29b) plays an important role in controlling DNA methylation through regulation of DNMTs. In primordial germ cells, miR-29b has been reported to modulate methylation of genomic DNA by targeting Dnmt3a and Dnmt3b, suggesting that it may have an important role in gonadal development in females (Takada et al., 2009). miR-29b has been found to be indispensable for early embryonic development in mouse. Altering the activity of miR-29b through the use of miR-29b mimic and inhibitor changed the DNA methylation levels resulting in failure of mouse preimplantation embryos to develop from morula to blastocyst stage (Zhang et al., 2015).
To our information, there is only one report in farm animals on the effects of miR-29b on embryonic development (Liang et al., 2018). In this study, we compared the expression level of miR-29b at different preimplantation stages of embryonic development in buffalo embryos produced by hand-made cloning (HMC) and their counterparts produced by in vitro fertilization (IVF). Since hypermethylation of genomic DNA is one of the major epigenetic abnormalities observed in cloned embryos (Dean et al., 2001; Kang et al., 2001), we explored the possibility of reducing DNA methylation by treatment of reconstructed embryos with miR-29b mimic. We also examined if this treatment could increase the blastocyst development rate and improve the quality of cloned blastocysts.
Materials and Methods
All the media and chemicals used were purchased from Sigma Chemical Co. (St. Louis, MO) and the plasticware was obtained from Nunc (Rosklide, Denmark), unless indicated otherwise. Fetal bovine serum (FBS) was obtained from Gibco Life Technologies (Carlsbad, CA). In vitro maturation (IVM) and IVF of oocytes and in vitro culture (IVC) of somatic cells were carried out in a CO2 incubator (5% CO2 in air). The reconstructed embryos generated by HMC and the presumptive zygotes generated by IVF were subjected to IVC in a CO2 incubator, which provided 5% CO2, 5% O2, and 90% N2 atmosphere.
Production of blastocysts by IVF
Cumulus–oocyte complexes (COCs) obtained from slaughterhouse ovaries were divided on the basis of their morphological appearance into two categories. Compact COCs, which possessed evenly granular homogenous ooplasm and unexpanded cumulus mass with two or more layers of cumulus cells, were classified as usable, whereas those which were wholly or partially denuded, or had an expanded cumulus mass or irregular ooplasm were classified as nonusable. Staining with Brilliant Cresyl Blue (BCB), as described previously (Mohapatra et al., 2015), was used for selection of COCs of high developmental competence. The COCs were incubated for 90 minutes with 26 μM BCB at 38.5°C in a CO2 incubator and then washed thrice with Dulbecco's phosphate-buffered saline (DPBS).
COCs of high developmental competence, which were marked by a distinct blue color (BCB+), were further processed for IVM and IVF as described previously (Sharma et al., 2011). IVC of presumptive zygotes was carried out for 8 days at 38.5°C in a CO2 incubator in Research Vitro Cleave Medium (K-RVCL-50; Cook, Queensland, Australia) containing 1% fatty acid-free bovine serum albumin (BSA) on original beds of granulosa cells, which had been generated during IVM. Blastocysts obtained on day 8 of IVC were stored at −80°C until further processing for RNA isolation.
Production of blastocysts by HMC
Collection of COCs, selection of usable quality COCs and BCB staining were carried out as explained above. The skin fibroblasts used as nucleus donors were obtained from ear biopsy of a bull (Mu-4354) and were between passages 5 and 7. IVM was done as explained above but only for 21 hours. All the steps of HMC, which included removing the cumulus cells and zona pellucida, enucleation of the oocytes, fusing the donor cell with two demicytoplasts, and activating the reconstructed embryos and their IVC, were carried out as described earlier (Selokar et al., 2014). Blastocysts obtained on day 8 of IVC were stored at −80°C until further processing for RNA isolation.
miRNA-29b mimic treatment
miR-29b mimic (miRCURY LNA™ microRNA mimics, FAM; Cat. No. 339173YM00470851-ADB) was obtained from Qiagen (Montgomery, MD). It was diluted in nuclease-free water to make a stock solution of 66.67 μM. A negative control of miRNA mimic molecules (miRCURY LNA microRNA Mimic Negative Control; Cat. No. E71003) was used to confirm that the manifested effects were due to miR-29b.
Transfection of the reconstructed embryos was carried out by incubating them for 1 hour with 200 μL Opti-MEM + 40 nM miR-29b mimic + 4 μL Lipofectamine at 38.5°C in a CO2 incubator after 1 hour of electrofusion. In case of negative controls, random sequence miRNA mimic molecules were used in place of miR-29b mimic. Then, the reconstructed embryos were washed three times with TCM-199 + 2.0 mM
Assessment of embryo quality by TUNEL assay
The quality of day 8 blastocysts was assessed by measuring the total cell number (TCN) and the level of apoptosis by TUNEL staining using the In Situ Cell Death Detection Kit, Fluorescein (11684795910; Roche). Briefly, after washing the blastocysts three times with polyvinyl alcohol (PVA, 0.3% in phosphate-buffered saline [PBS]), these were fixed by treatment with 4% paraformaldehyde for 1 hour. The blastocysts were then permeabilized by treatment with 0.5% Triton X-100 for 1 hour, and then treated with FITC-conjugated dUTP and terminal deoxynucleotidyl transferase (TdT) enzyme for 1 hour at 37°C in the dark. The blastocysts were treated with RNase (50 μg/mL) following which, the nuclei were stained by treatment with Hoechst 33342 (10 μg/mL) for 20 minutes at 37°C.
The blastocysts of the positive control group were treated with DNase solution (100 U/mL) for 20 minutes at 37°C before incubation with FITC-conjugated dUTP and TdT for 1 hour. After washing thoroughly with Ca2+ and Mg2+-free DPBS, the blastocysts were mounted on glass slides in antifade solution (3 μL) and flattened with the help of a cover slip. An inverted fluorescence microscope was used for obtaining the digital images for counting the cell number. Each experiment was repeated three times or more. The apoptotic index of each blastocyst was calculated using the equation (number of TUNEL-positive nuclei/the total number of nuclei) × 100.
Differential staining
The inner cell mass (ICM):trophectoderm (TE) cell number ratio of cloned and IVF blastocysts was determined by differential staining. The blastocysts were treated with paraformaldehyde (4%) for fixing and Triton X-100 (0.5%) for permeabilization. Following overnight incubation with anti-CDX2 primary antibody (ready-to-use, AM392-10M; Bio-Genex, Inc., San Ramon, CA) at 4°C, the blastocysts were washed three times with PBS-PVA and then treated for 90 minutes with the anti-mouse antibody (1:200, F0257-0.5ML; Sigma Chemical Co.), which was conjugated with FITC. Following a thorough washing with PBS-PVA, the blastocysts were treated with Hoechst 33342 (10 μg/mL) for 20 minutes for staining the nuclei. After thorough washing with PBS-PVA, the blastocysts were mounted on slides using glycerol and examined under a fluorescence microscope (Eclipse; Nikon, Tokyo, Japan). The same optical conditions were employed for capturing all the images.
Immunofluorescence staining for 5-methylcytosine
Blastocyst-stage embryos were fixed with 4% paraformaldehyde, permeabilized with 0.5% Triton X-100, and after washing three to four times with PBS-PVA, these were treated with 4 N HCl for 15 minutes and then with 4 N Tris HCl for 10 minutes. After washing thoroughly with PBS-PVA, the blastocysts were blocked by treatment with 1% BSA for 1 hour and were then subjected to overnight incubation at 4°C with anti-5 mC primary antibody (1:100, Cat. No. ab10805; Abcam). After several washings with PBS-PVA, the blastocysts were incubated with FITC-conjugated anti-mouse antibody (1:100, F0257-0.5ML; Sigma Chemical Co.) for 90 minutes. Following a thorough washing with PBS-PVA, the blastocysts were treated with PI (1:80) for 20 minutes for counterstaining the nuclei. After another thorough wash with PBS-PVA, the blastocysts were mounted on slides in glycerol and observed under a fluorescence microscope. The same optical conditions were employed for capturing all the images.
qRT-PCR (quantitative real-time polymerase chain reaction) for expression analysis of miR-29b
RNA was isolated from embryos at different stages of development, produced by HMC and IVF (three biological replicates for each group). The Norgen's Single Cell RNA Isolation Kit (Thorold, Canada) was used for isolation of total RNA. The purity of the total RNA obtained from each pool of embryos was examined by measuring A260/280 ratio, which was found to be between 1.8 and 2.1. Complementary DNA (cDNA) of miRNAs was synthesized using the Universal cDNA Synthesis Kit (Exiqon, Vedbaek, Denmark). Briefly, 2 μL enzyme mix, 20 ng of total RNA, 1 μL synthetic RNA spike-in (UniSp6), and 4 μL 5 × reaction buffer were used. Nuclease-free water was used for adjusting the total volume to 20 μL.
Following incubation of the mixture for 60 minutes at 42°C, reverse transcriptase was heat inactivated by incubation for 5 minutes at 95°C. The mixture was then cooled to 4°C immediately. The cDNA produced was stored at −80°C until further processing for qRT-PCR.
CFX96 real-time system (Bio-Rad, Hercules, CA) was used for carrying out gene amplification for qRT-PCR. The reaction mixture contained 4 μL diluted cDNA and 6 μL of a mixture consisting of 5 μL ExiLENT SYBR® Green master mix (Exiqon) and 1 μL of LNA™ Universal RT primer-set (Supplementary Table S1). There were three technical replicates for each miRNA sample along with nontemplate control. The following amplification conditions were used for performing the reactions: 10 minutes at 95°C, 40 cycles of 15 seconds at 95°C (denaturation), and 1 minute at 60°C (annealing+extension). The internal control genes used for normalization of data included miR-423 and miR-103 (Mahdipour et al., 2015).
qRT-PCR for expression analysis of mRNAs
The expression level of genes in blastocysts produced by HMC was examined by qRT-PCR. The blastocysts produced from miR-29b mimic-treated reconstructed embryos and untreated controls were pooled to create three biological replicates. The method described above was used for the isolation of total RNA. The SuperScript III Kit (Invitrogen, Carlsbad, CA) was used for the preparation of cDNA with 1 μL dT12–18, 100 ng RNA, 1 μL random primers, 1 μL 10 mM dNTP mix, and 10 μL DNase-RNase-free water.
Following incubation at 65°C for 5 minutes, the mixture was kept on ice for 3 minutes. A total of 6 μL of Master Mix, composed of 4.5 μL 5 × First-Strand Buffer, 1 μL 0.1 M DTT, 0.25 μL (50 U) of SuperScript III RT, and 0.25 μL of RNase inhibitor (10 U; Promega, Fitchburg, WI), was added. The program used for performing the reaction was: 25°C for 5 minutes, 50°C for 60 minutes, and 70°C for 15 minutes.
Then, the cDNA was diluted 1:4 (v:v) using DNase-RNase-free water, following which gene amplification was carried out using Maxima SYBR Green Master Mix (Fermentas, St. Leon-Rot, Germany) using primer sets (Supplementary Table S2) at the following thermal cycling conditions: 95°C for 5 minutes, followed by 40 polymerase chain reaction cycles of 95°C for 15 seconds, 60°C for 30 seconds, and 72°C for 30 seconds. The internal control genes used for normalization of data included GAPDH and HPRT1 (Sood et al., 2017).
Experimental design and statistical analysis
Embryos at 2-, 4-, and 8-cell, morula, and blastocyst stages were produced by HMC and IVF. Then, we determined the expression level of miR-29b in these embryos by qRT-PCR and compared it between cloned and IVF embryos at each embryonic stage and among all the embryonic stages in each group. Following this, we examined the effects of treatment of reconstructed embryos with miR-29b mimic on the developmental competence and quality of cloned embryos. The reconstructed embryos were divided into the following three groups in each trial. No treatment was given to the reconstructed embryos of Group 1 (control). In case of Group 2, the reconstructed embryos were transfected with miR-29b mimic after 1 hour of electrofusion. Reconstructed embryos of Group 3 (negative control) were treated with random sequence miRNA mimic molecules in the same manner as those of Group 2.
The developmental competence was assessed by recording the blastocyst rate on day 8 of IVC. The quality of cloned embryos was assessed by measuring the TCN, apoptotic index, and the ratio of ICM:TE cell numbers. IVF blastocysts were used as controls for the last two parameters. Then, we examined the effects of miR-29b mimic treatment on the level of methylation in cloned blastocysts by immunofluorescence staining. Finally, we investigated the effects of this treatment on the relative mRNA abundance of some important epigenetic- (DNMT1, DNMT3A, and DNMT3B), pluripotency- (OCT4, NANOG, and SOX2) and development-related genes (GLUT1 and FGF4) by qRT-PCR in cloned blastocysts.
The data were analyzed by one-way analysis of variance followed by Tukey's or Dunnett's test using GraphPad Prism 7 software. Student's t test was used for comparing the mean of different groups. The percentage values were subjected to arcsine transformation before analysis. Data have been presented as mean ± standard error of the mean.
Results
Expression level of miR-29b in early embryos
miR-29b expression level was highest at the 2-cell stage, decreased significantly (p < 0.001) at the 4-cell stage, and was low thereafter at the 8-cell, morula, and blastocyst stages. Its expression was similar in embryos produced by HMC and IVF (Fig. 1A, B). miR-29b expression was significantly (p < 0.001) lower in cloned compared with IVF embryos at 2-, 4-, and 8-cell and blastocyst stages, however, at the morula stage, it was higher (p < 0.01) in cloned than in IVF embryos (Fig. 1C).

qRT-PCR (quantitative real-time polymerase chain reaction) was used for measuring the expression level of miR-29b in cloned and IVF early embryos. In
Effect of miR-29b on developmental competence and quality of cloned embryos
Occurrence of transfection was confirmed by examining the reconstructed embryos, transfected with miR-29b mimic, under a fluorescence microscope (Fig. 2). In the 14 trials conducted, the blastocyst rate, which was 27.0% ± 2.0% (48/185) for the controls, 23.0% ± 1.0% (46/198) for the negative controls, and 29.0% ± 2.0% (60/208) for the miR-29b-treated group, was not significantly different.
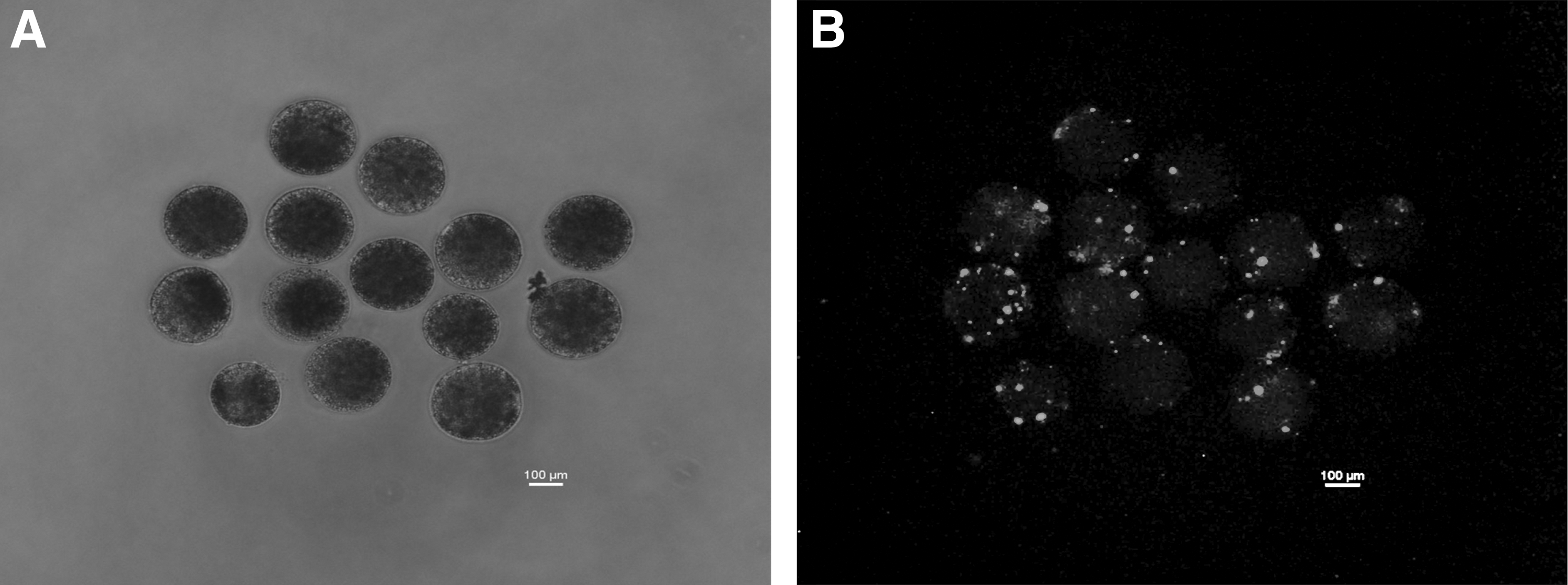
Reconstructed embryos transfected with FITC-conjugated miR-29b mimic by lipofection
TUNEL assay was used to determine the TCN and the apoptotic index in blastocyst-stage embryos produced by HMC and IVF. TCN was higher (p < 0.05) in blastocysts produced from miR-29b mimic-treated reconstructed embryos compared with the untreated controls (Table 1). miR-29b mimic treatment decreased (p < 0.05) the apoptotic index of blastocysts compared with nontreated controls to the level observed in IVF blastocysts (Table 1). miR-29b treatment increased (p < 0.05) the ICM:TE cell number ratio compared with untreated controls to values observed in IVF blastocysts (Fig. 3 and Table 2).
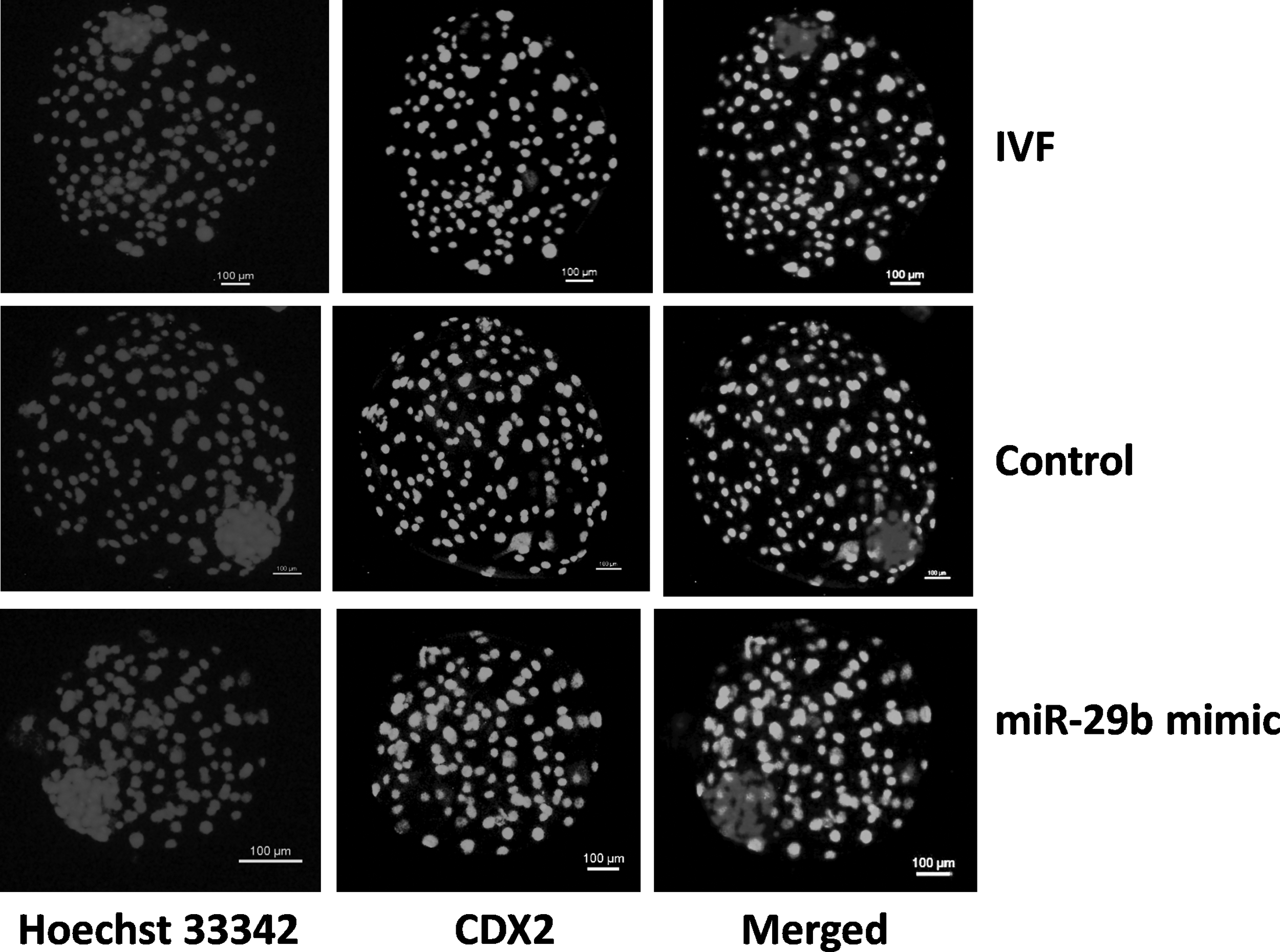
Differential staining was used for determination of the ratio of cell numbers in inner cell mass:trophectoderm in blastocysts produced from reconstructed embryos treated with miR-29b mimic, untreated controls and IVF blastocysts. Staining of nuclei with Hoechst 33342 was used for determining total cell number, whereas staining with FITC-conjugated anti-CDX2 was used for determining the cell number of trophectoderm.
Effect of Treatment of Reconstructed Embryos with MicroRNA-29b Mimic on the Total Cell Number and Apoptotic Index of Cloned Embryos
Values within a column having different superscripts differ significantly (p < 0.05).
IVF, in vitro fertilization; miR-29b, microRNA-29b.
Effect of Treatment of Reconstructed Embryos with MicroRNA-29b Mimic on the Ratio of Inner Cell Mass:Trophectoderm Cell Numbers in Cloned Blastocysts
Values within a column having different superscripts differ significantly (p < 0.05).
ICM, inner cell mass; TE, trophectoderm.
Effect of miR-29b on the level of methylation in cloned embryos
Immunofluorescence staining against 5-methylcytosine (5-mC) revealed that DNA methylation level was significantly lower (p < 0.05) in the blastocysts produced from miR-29b mimic-treated reconstructs compared with untreated controls (Table 3 and Fig. 4).

Immunofluorescence staining against 5-methylcytosine in blastocysts produced from reconstructed embryos treated with miR-29b mimic and untreated controls. As indicated by mean pixel intensity, DNA methylation level was lower in the blastocysts produced from miR-29b mimic-treated reconstructs compared with untreated controls.
Effect of Treatment of Reconstructed Embryos with MicroRNA-29b Mimic on the Level of DNA Methylation in Cloned Blastocysts
Values within a column having different superscripts differ significantly (p < 0.05).
Effect of miR-29b on gene expression in cloned embryos
Among epigenetic-related genes, the expression level of DNMT3A and DNMT3B, but not that of DNMT1, was lower (p < 0.001) in blastocysts produced from miR-29b mimic-treated reconstructs than in untreated controls (Fig. 5). The expression level of genes associated with pluripotency (OCT4, p < 0.05; NANOG and SOX2, p < 0.01) and development (GLUT1, p < 0.01; FGF4, p < 0.05) was higher in blastocysts generated from miR-29b mimic-treated reconstructs compared with the untreated controls.

Relative expression level of some important genes in blastocysts produced from reconstructed embryos treated with miR-29b mimic. Blastocysts produced from untreated reconstructs served as controls, whereas those from reconstructs treated with random sequence miRNA mimic molecules, served as negative controls (NC). Bars marked with an asterisk differ significantly from the corresponding value. *p < 0.05; **p < 0.01; ***p < 0.001. miRNA, microRNA.
Discussion
The overall aim of the present study was to investigate the role of miR-29b during early embryonic development. In the first part of the study, buffalo embryos at different stages of development were produced by HMC and IVF, and miR-29b expression was compared among these stages in each group. In the IVF embryos, the miR-29b expression, which was highest at the 2-cell stage, decreased significantly at the 4-cell stage, and was low thereafter at the 8-cell, morula, and blastocyst stages.
A similar expression pattern of miR-29b has been reported previously in mouse IVF embryos (Zhang et al., 2015). Our study extends these results by demonstrating that the expression pattern of miR-29b in cloned embryos is similar to that in IVF embryos. Then, we compared miR-29b expression between cloned and IVF embryos at each developmental stage. We found that miR-29b expression was lower in embryos produced by HMC compared with IVF embryos at the 2-, 4-, and 8-cell, and blastocyst stage, which agrees with the results of a recent study in which miR-29b expression was observed to be lower in bovine SCNT embryos at the pronuclear, 8-cell, and blastocyst stages compared with their IVF counterparts (Liang et al., 2018).
In the second part of the study, the reconstructed embryos were treated with miR-29b mimic and the effects of this treatment on the developmental competence and quality of cloned embryos were investigated. We found that miR-29b mimic treatment did not affect the blastocyst rate significantly compared with controls. In bovine, miR-29b inhibitor treatment of SCNT zygotes reduced the cleavage and blastocyst rate, whereas miR-29b mimic treatment did not affect them (Liang et al., 2018). Similarly, microinjection of miR-29b inhibitor into mouse IVF zygotes resulted in a developmental block between morula and blastocyst stages, and only 50%–60% embryos developed to blastocysts in comparison with negative and noninjected control groups (Zhang et al., 2015).
Altogether, these results suggest that overexpression of miR-29b does not improve the developmental competence of cloned embryos, whereas downregulation of miR-29b affects it adversely. Interestingly, treatment of mouse IVF zygotes with miR-29b mimic adversely affected the developmental competence resulting in a block between morula-to-blastocyst stages, similar to that observed following treatment with miR-29b inhibitor (Zhang et al., 2015). This could be due to differences in species or embryo type.
Since embryo quality is one of the most important factors influencing the live birth rate, we studied the effects of miR-29b mimic treatment on the quality of cloned embryos using several parameters. Since TCN is believed to directly influence the embryonic development after implantation in mammals, it is employed for evaluating the quality of blastocysts (Yu et al., 2007). TUNEL assay revealed that miR-29b mimic treatment of reconstructed embryos resulted in a significant increase in the TCN of cloned blastocysts compared with untreated controls. The incidence of apoptosis is another important criterion to judge embryo quality. Apoptosis level is higher in cloned compared with IVF embryos (Cui et al., 2011; Selokar et al., 2013), and has been reported to be correlated with a decrease in the TCN (Yu et al., 2007).
We found that miR-29b mimic treatment of reconstructed embryos significantly decreased the apoptotic index of cloned blastocysts compared with untreated controls to levels observed in IVF blastocysts. Interestingly, these apoptotic actions of miR-29 in embryos are in contrast to those observed in several other cell types such as hepatocellular carcinoma cells (Xiong et al., 2010) and chondrocytes (Chen et al., 2017), in which it has been reported to promote apoptosis.
Mammalian blastocysts are composed of the ICM, which forms the embryo, and the TE, which forms the extraembryonic tissues of the placenta. An appropriate ratio of cell numbers between ICM and TE is necessary for normal embryonic development and is employed for assessing the quality of blastocysts (Van Soom et al., 1996; Yu et al., 2007).
Abnormal allocation of ICM and TE cells in preimplantation embryos leads to high incidences of placental abnormalities and early embryonic mortality observed in cloned embryos (Im et al., 2006). Abnormally high ICM and low TE cell number in SCNT embryos has been reported to result in insufficient placental development (Koo et al., 2002). Lower number of ICM cells and TCN in SCNT embryos compared with IVF embryos is correlated with abnormal Oct4 expression (Boiani et al., 2003). It was found in the present study that treatment of reconstructed embryos with miR-29b mimic increased the ICM:TE cell number ratio compared with untreated controls to levels observed in IVF blastocysts. These results demonstrate that miR-29b treatment improved the quality of cloned embryos in terms of all the three parameters examined in the present study.
Then, we examined the effect of miR-29b mimic treatment on the level of DNA methylation in blastocysts by immunofluorescence staining against 5-mC. DNA methylation level was significantly lower in the treatment group than in untreated controls. This was further corroborated by significantly lower expression of DNMT3A and DNMT3B, but not that of DNMT1, observed in blastocysts of the treatment group, compared with untreated controls.
Similar results have been reported in a recent study in bovine (Liang et al., 2018). An inverse correlation of miR-29b with Dnmt3a/3b expression in mouse IVF embryos suggests that Dnmt3a/3b were targets of miR-29b and were negatively regulated by it (Zhang et al., 2015). miR-29 family has been reported to target DNMTs in several types of cells. miR-29b targets both DNMT3A and DNMT3B in lung cancer leading to reversal of aberrant DNA methylation (Fabbri et al., 2007). miR-29b targets DNMT3A and 3B directly and DNMT1 indirectly leading to induction of tumor suppressor gene reexpression and global DNA hypomethylation in acute myeloid leukemia (Garzon et al., 2009). miR-29b regulates genomic DNA methylation in mouse primordial germ cells by targeting Dnmt3a and Dnmt3b (Takada et al., 2009).
Cloned embryos show aberrant patterns of the global DNA methylation (Kang et al., 2001). DNA hypermethylation, which is one of the major epigenetic abnormalities observed in cloned embryos, is associated with their developmental potential (Dean et al., 2001; Santos et al., 2003). DNA demethylation, therefore, appears to be an important factor limiting the cloning efficiency. Epigenetic modulators, such as 5-aza-2′-deoxycytidine, which is a DNMT inhibitor, have been used for modifying the epigenetic status of cloned embryos by decreasing their DNA methylation (Enright et al., 2005). miR-29b has been reported to reduce DNA methylation by regulating DNMTs in preimplantation embryos in mouse (Zhang et al., 2015).
Recent studies show that DNA methylation can be altered in bovine SCNT embryos (Liang et al., 2018) and pig IVF embryos (Zhang et al., 2018) by treatment with miR-29b mimic or inhibitor. Our results agree with these studies and suggest that treatment of SCNT embryos with miR-29b mimic could be an effective approach for reducing DNA hypermethylation in SCNT embryos.
Several pluripotency-related genes, such as NANOG, OCT4, and SOX2, play an important role in embryonic development. miR-29b mimic treatment of reconstructs increased the expression level of these three genes significantly in blastocysts. Inhibition of miR-29b in mouse embryos resulted in downregulation of Nanog (Wang et al., 2016). Sox2, one of the four Yamanaka factors (OSKM), directly regulates the expression of miR-29b during production of induced pluripotent stem cells, and miR-29b expression has been found to be essential for OSKM- and OSK-mediated reprogramming (Guo et al., 2013). Moreover, the effects of miR-29b on reprogramming could be blocked by overexpressing Dnmt3a or Dnmt3b. Altogether, this suggests that in addition to reducing hypermethylation, miR-29b may be playing a role in reprogramming in SCNT embryos.
We found that miR-29b treatment of reconstructed embryos significantly increased the expression of development-related genes, GLUT1 and FGF4, in blastocysts than in untreated controls. GLUT1, which facilitates the transport of glucose and glucosamine across plasma membrane, influences preimplantation embryo development by regulating the uptake of glucose, and the embryo has innate ability to control the uptake of glucose by regulating its expression (Leppens-Luisier et al., 2001). GLUT1 expression is linked to the need of the embryos for glucose present in embryonic environment since in vitro-produced blastocysts, which are present in constantly high glucose environment, have less than half the expression of this gene than in vivo-produced blastocysts (Morita et al., 1994).
GLUT1 is present throughout embryonic development, with the peak expression at 16-cell stage when embryo metabolism switches from being glycolytic-centric to glycogenic-centric in bovine (García-Herreros et al., 2018). It would be interesting to find out if the high GLUT1 expression in miR-29b-treated embryos leads to uptake of extra glucose, to be stored as glycogen to enable the embryos better withstand the anorexic environment following transfer to recipients.
High GLUT1 expression following miR-29b treatment may have contributed to lowering of the level of apoptosis seen in the blastocysts of the treatment group since a decrease in GLUT1 expression and reduced glucose transport have been reported to trigger apoptosis in blastocysts (Chi et al., 2000). FGF-4, a mitogen, which exhibits strong action on numerous different cell types, plays a role in embryonic development, morphogenesis, and proliferation, and differentiation of embryonic stem cells and tissue stem cells (Kosaka et al., 2009). It plays an important role in some of the critical events during preimplantation embryonic development, such as segregation of the ICM cells into epiblast and hypoblast (Kuijk et al., 2012).
It is interesting to note that the effects of miR-29b mimic treatment of reconstructed embryos could be observed 8 days later in blastocyst-stage embryos. This could be due to the use of locked nucleic acid (LNA) bases for synthesizing the miR-29b mimic. LNAs are ribonucleotide analogs, which contain a methylene linkage between the 2′-oxygen and 4′-carbon of the ribose ring. miRNA mimics synthesized from LNA bases offer several advantages such as high affinity and specificity toward the target strand, high in vivo stability, lack of toxicity, and ease of transfection into cells (Kaur et al., 2007).
In conclusion, our results show that miR-29b exhibits a typical expression pattern during preimplantation development of embryos, which is similar in IVF and cloned embryos. The results of our study also suggest that treatment of reconstructed embryos with miR-29b mimic results in improvement in the quality of blastocyst-stage embryos as indicated by normal cell number ratio of ICM:TE cells, lower apoptotic index, and higher TCN. However, miR-29b treatment did not affect the blastocyst rate significantly. miR-29b mimic treatment decreases the DNA methylation and expression of DNMT3A and DNMT3B and increases the expression level of many genes associated with pluripotency and development.
Footnotes
Acknowledgments
The financial support for this work came from Science and Engineering Research Board, Department of Science & Technology, Ministry of Science and Technology, Government of India. S. Sah and S. Shyam were supported by fellowships from ICMR and UGC (India), respectively.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.