
Abstract
Abstract
Stromal mesenchymal stem cells (MSCs) have the capability to self-renew and can differentiate into multiple cell types of the mesoderm germ layer, but their properties are affected by molecular aging mechanisms. MSCs can be obtained from adipose tissue termed as adipose-derived stem/stromal cells (ASCs) representing a promising tool for studying age-related diseases in detail. ASCs from young (16 weeks) and old (>108 weeks) rabbits were successfully isolated and propagated. ASCs showed the typical morphology and stained positive for CD105, Vimentin, Collagenase 1A, and negative for CD14, CD90, and CD73, demonstrating their mesenchymal origin. ASCs expressed MSC markers, including MYC, KLF4, CHD1, REST, and KAT6A, whereas pluripotency-related genes, such as NANOG, OCT4, and SOX2, were not expressed. Aged ASCs showed altered protein and mRNA levels of APOE, ATG7, FGF2, PTEN, and SIRT1. Adipogenic differentiation of old visceral ASCs was significantly decreased compared with young visceral ASCs. We successfully established rabbit ASC cultures representing an in vitro model for the analysis of stem cell aging mechanisms. ASCs, obtained from old female rabbits, showed age- and source-specific alteration due to aging of the donor. Stem cell plasticity was altered with age as shown by reduced adipogenic differentiation capacity.
Introduction
Since 1990, the average life expectancy rose up to 6 years worldwide. In 2050, the population group of <60 years will be nearly doubled (United Nations Reports, 2015). Old age represents the most important risk factor for aged-related diseases, including type 2 diabetes, Alzheimer's, and cardiovascular diseases (Goodell and Rando, 2015; Rando, 2006). Therefore, the understanding of age-dependent tissue and organ function is a major issue of medical research in the aging society. The age-dependent loss of stem cells and their impaired regeneration may be a major reason for age-related diseases (Ho et al., 2005; Oh et al., 2014). Thus, adult stem cells are very important in proving this hypothesis. They are preserved as a pool of stem cells remaining from embryonic development for ensuring tissue growth and maintenance during the postembryonic life span.
Self-renewal and the capacity to differentiate into derivatives of the local organ tissue describe essential properties of adult stem cells defining the stem cell plasticity. The stem cell plasticity undergoes age-related changes and old adult stem cells downregulate the dividing capacity and dysregulate its differentiation (Jung and Brack, 2014). Even though this principle has been described, the molecular and cellular mechanisms behind are poorly understood.
Mesenchymal stem cells (MSCs) have been studied intensively in the context of aging (Bruna et al., 2016; Lohmann et al., 2012). Adipose-derived stem/stromal cells (ASCs) are of particular interest for analyzing aging and the development of age-related disorders, because they are easily accessible in large numbers. Recently, stem cell-based in vitro models became important for analyzing stem cells in general due to the technical advance within the last years (Jung et al., 2015). However, primary cultures create a more complex system and are closer to cellular changes by whole-body interactions.
This advantage highly recommends combining primary stem cell models and animal models to investigate time- and organ-related biological processes. Rabbits are suitable models investigating cardiometabolic diseases in humans (Niimi et al., 2016). We applied the rabbit as an in vivo model in the context of age-related diseases, especially in the context of the metabolic pathways (Fischer et al., 2017) for many years.
In the present study, we found cellular phenotypes of rabbit ASCs, which contribute to disturbed maintenance and regeneration of adipose tissue in old age. We established a scientific model combining the advantages of in vivo and in vitro models for mimicking age-related changes in the stem cell plasticity of ASCs. These data point to general stem cell aging mechanisms and may provide new insights into age-related disturbance of stem cell plasticity in healthy aging and age-related diseases.
Materials and Methods
Isolation of rabbit ASCs
ASCs were isolated from the abdominal visceral and subcutaneous adipose tissue of young (16 weeks) and old female (>108 weeks) New Zealand white rabbits (hybrid strain ZIKA). The experimental studies were approved by the Local Ethics Commission of Landesverwaltungsamt Dessau (Saxony-Anhalt, Germany) and conducted in accordance with the principles of their approved guidelines. Single caged rabbits were housed under controlled light and temperature conditions (12-h light–12-h dark cycle; 22°C ± 1°C) with free access to food and water.
Rabbit ASCs were isolated according to the human protocol of Zhu et al. (2013), adjusted, and modified for rabbit cells (Fig. 1). Around 30 g–50 g of rabbit adipose tissue was washed in sterile phosphate-buffered saline (PBS) several times and was prepared by mincing and dissociation with 0.08% Collagenase A (Sigma Aldrich) for 40 minutes (visceral tissue) or 50 minutes (subcutaneous tissue) at 37°C with gentle agitation. To inactive Collagenase A activity, we added DMEM (Gibco) culture medium (CM) supplemented with 10% fetal bovine serum (FBS) (Biochrom), 100 U/mL penicillin, and 100 μL/mL streptomycin (PAA GmbH) at equal volume to each probe for 5 minutes. Cell residues were removed by a nylon mesh with a pore size of 100 μm.

Isolation and Expansion of young and old rabbit VASCs. Mechanical and enzymatic digestion of primary ASCs
After washing and centrifugation (500 g; 10 minutes), cell pellets containing the stromal vascular fraction (SVF) were resuspended in CM and plated on gelatin-coated culture dishes. To remove any containing red blood cells, the medium was changed 24 hours after isolation and cells were washed twice with the CM.
Expansion of rabbit ASCs
Cells were maintained at 37°C/5% CO2 and the culture medium was changed three times a week. At confluence of 80%–90%, we harvested cells using 0.08% EDTA and portioned cells 1:2 into 25-cm flasks. Until the first passage, CM was supplemented with 1% gentamicin and 1% amphotericin B (Biochrom). Cells were analyzed morphologically by microscopy (Nikon) at day 1, 3, 5, and 8 (Fig. 1B). ASCs at Passage 3 and 4 were used for the characterization and adipogenic differentiation.
Adipogenic differentiation of rabbit ASCs
Undifferentiated rabbit visceral adult stem/stromal cells (VASCs) of Passage 4 were plated in six-well plates with a density of 40,000 cells/well. After 6 days undifferentiated growth adipogenic differentiation was induced by DMEM supplemented with 1 μM dexamethasone (Merck), 500 μM 3-isobutyl-1-methylxantine (IBMX) (Merck), 200 μM indomethacin (Merck), 10 μg/mL insulin (Merck), 10 ng/mL BMP4 (PeproTech), 10% FBS, 100 U/mL penicillin, and 100 μL/mL streptomycin for 14 days. Medium was changed every3 days. Undifferentiated VASCs in DMEM without adipogenic supplements were used as negative controls. After 14 days, cells were washed with PBS and analyzed by flow cytometry as described previously (Biemann et al., 2012). For microscopy analysis, cells were fixed with 4% PFA for 10 minutes, and stained with 0.5% Oil Red O solution for 2 hours. Oil Red O stains lipids and is an indicator of neutral lipid droplets in adipocytes.
Flow cytometry of cell surface markers of undifferentiated VASCs
VASCs from young and old rabbits were cultured in CM 48 hours before flow cytometry analysis. Detached cells were centrifuged (1200 g; 5 minutes). Cell aliquots (5 × 105) were suspended in 100 μL PBS. Then cells were incubated with an antibody cocktail containing the monoclonal antibodies: CD14-Alexa Fluor, CD44-APC, CD45-PE-Y, CD73-Alexa Fluor 700, CD90-Alexa Pacific Blue, and CD105-PE-Cy7 (Miltenyi Biotech). The incubation with the antibodies was 10 minutes at 2°C–8°C in the dark.
After incubation and washing with PBS, cells were analyzed by flow cytometry (LSR II Fortessa; BD biosciences). The leader of the core facility flow cytometry performed the gating and measurement unknowing the type of samples. Data were analyzed with the BD FACSDiva Version 6.1.3 and the FlowJo 10.1 software (BD Biosciences). All antibodies used in flow cytometry analysis were first tested for binding epitopes in rabbit cells. Antibodies used against rabbit cells in this study analyzed for proteins by flow cytometry, so far as we know, are not described in the literature.
Immunofluorescence imaging
After fixation in 4% paraformaldehyde/PBS, cells on coverslips were permeabilized with 0.2% TritonX-100 and 3.5% horse serum for 30 minutes, blocked in PBS with 3.5% horse serum for 30 minutes, then incubated with primary antibodies Vimentin (Merck Millipore) or Collagen 1A1 (Sigma Aldrich) at 4°C overnight. Coverslips were washed in PBS and then incubated with the second antibody goat Alexa Fluor 488 anti-mouse IgG (Thermo Fisher Scientific) at 4°C overnight. The nuclei were stained with Hoechst 33342 (Thermo Fisher Scientific). At once cells were rinsed with PBS and at the end washed with ultrapure water. Fluorescent images were obtained with Keyence Biozero fluorescence microscope.
Proliferation assay and population doubling time
VASCs of passage 3–4 were used for the proliferation assay. Cells were seeded in triplicates in six-well-plates with a density of 25,000 cells per well and maintained with DMEM and 10% FBS. VASCs were harvested and counted with a Neubauer counting chamber on day 1, 3, 5, 7, 9, 11, and day 13. The resulting growth curve was logarithmized and the slope factor was calculated. The population doubling time (PDT) was than calculated according to the equation: N (t) = N0 * eμ*t (N0-initial quantity, N-quantity after time (t), t-time in hours, μ-slope factor).
RNA extraction and reverse transcription reaction
Undifferentiated (Passage 3) VASCs and subcutaneous adipose-derived stem/stromal cells (SASCs) and adipogenic differentiated VASCs (Passage 4) were homogenized by using Precellys (Bertin Technologies) two times for 20 seconds. Total RNA extraction was performed by using TRI Reagent (Merck). Protocols for total RNA extraction and cDNA synthesis were used as described before (Navarrete Santos et al., 2000).
The RNA expression of the genes of interest (MYC, CHD1L, REST, KLF4, KAT6A, OCT4, SOX2, NANOG, ADIPONECTIN, FABP4, and PPARγ) was detected by reverse transcription (RT)–PCR, using specific oligonucleotide primers (Table 1) on cDNA from the cells investigated. RT-PCR was performed by 40 cycles (94°C for 45 seconds, 60°C for 45 seconds, and 72°C for 60 seconds). The amplification of the housekeeping gene GAPDH was used to verify quality of the sample cDNA. Resulting RT-PCR products were visualized in a 2% agarose gel.
Primers Used for Reverse Polymerase Transcription–Polymerase Chain Reaction and Quantitative Real-Time Reverse Polymerase Transcription–Polymerase Chain Reaction
Quantitative real-time PCR
We quantified the RNA expression of specific target genes in VASC and SASC samples by quantitative RT-PCR (qRT-PCR) using StepOnePlus™ System (Applied Biosystems) and SYBR green (Eurogentec, Köln). The nucleotide sequences of the used primers are listed in Table 1. Each assay included duplicates of each cDNA sample and a “no template” control. Absolute mRNA were calculated as amounts (molecules per molecule-specific standard DNA) of primer-specific plasmid standards, using dilutions in a range from 107 to 102. The amount of the housekeeping gene GAPDH was used to normalize samples for cDNA used as per reaction. To confirm the amplification, the resulting qRT-PCR products were analyzed by dissociation curves.
Protein preparation and immunoblotting
Protein preparation, quantification, and western blot were performed with 30 μg protein. After blotting, the nylon membrane was stained with Ponceau S. The immunoreactive signals were visualized by enhanced chemiluminescence (Millipore, Schwalbach, Germany) and quantified by Fusion Fx7 imaging system (Peqlab, Erlangen, Germany). The primary antibodies were incubated at 4°C overnight The following antibodies were used: anti-APOE (mouse Ab, 1:500, #NB110-60531; Biotechne), anti-ATG7 (mouse Ab,1:1000,#MAB6608; Biotechne), anti-FGF2 (mouse Ab 1:100,#; Santa Cruz), anti-PTEN (mouse Ab, 1:200,#; Santa Cruz), anti-SIRT1 (mouse Ab,1:1000,NBP1-51641SS; Biotechne), anti-β-Actin (mouse Ab, 1:20000, A-5441; Sigma-Aldrich), anti-mouse IgG conjugated to HRP (1:10,000; Dianova, Hamburg). Protein amounts were calculated as ratio band intensities (protein of interest vs. β-Actin) in the same blot.
Statistical analysis
Statistical differences between groups (young ASCs vs. old ASCs) were calculated using Students t test after proven normal distribution (SigmaPlot v.12.0). Differences between groups were considered as statistically significant if p < 0.05. Data are presented as mean ± SEM. Controls were taken as reference group.
Results
Isolation of rabbit VASCs
Rabbit VASCs were isolated from 30 g to 50 g visceral adipose tissue from young and old female New Zealand white rabbits (Fig. 1A). After mincing and washing with PBS, the tissue was digested by collagenase A (Fig. 1A, I–IV). After centrifugation and washing steps, the resulting SVF was counted with Neubauer counting chamber and plated on 25 cm flasks. Isolation of cells from young and old donors did not show a significant difference between cell number per 1 g of adipose tissue (young: 3442 ± 1131 cells/g; old: 3940 ± 422 cells/g; n = 4 per group).
Expansion of rabbit VASCs
After reaching confluence of 80%–90%, isolated VASCs from young and old donors showed MSC characteristics, such as bipolar and spindle-shaped morphology and growth (Fig. 1B). The analysis of the PDT of young and old undifferentiated VASCs revealed no significant differences between groups (young: 33.7 ± 6.9 hours; old: 34.7 ± 6.9 hours; n = 4 per group, Fig. 1C).
No morphologic differences could be observed during culture between young and old VASCs and between young and old SASCs (data not shown).
Characterization of undifferentiated rabbit VASCs
To verify mesenchymal stem/stromal cell properties, we analyzed the expression of surface antigens CD14, CD45, CD44, CD73, CD90, and CD105 in young and old VASCs by flow cytometry. Moreover, the expression of specific mesenchymal markers, Vimentin and Collagen 1A1, was detected by immunofluorescent staining in VASCs (Fig. 2A).

The MSC phenotype characterization of rabbit VASCs. Immunofluorescence detection of COL1A1 and Vimentin of young VASCs (
Surface antigens, CD14, CD73, and CD90, were not expressed, whereas two hematopoietic surface markers, CD45 and CD44, were slightly expressed in young and old rabbit VASCs (Fig. 2B and Table 2). Also CD 105 was present in young and old rabbit VASCs (young: 89% ± 3.3%; old: 74% ± 8.1%) (Fig. 2B and Table 2). The expression levels of surface antigens did not differ in old rabbit VASCs (Table 2).
Expression of Surface Antigens from Rabbit Visceral Adipose-Derived Stem/Stromal Cells
Data are presented as mean ± SEM from three independent experiments (n = 4).
The multipotency and pluripotency potential of the isolated cells was investigated by RT-PCR analysis. Transcripts of multilineage genes, CHD1, KAT6A, KLF4, MYC, and REST, were detected in young and old rabbit VASCs. The pluripotency-related genes, OCT4, NANOG, and SOX2, were not expressed in young and old rabbit VASCs (Fig. 2C; Supplementary Data).
Significant decrease of adipogenic differentiation of old VASCs
To investigate adipogenic differentiation potential of young and old rabbit VASCs, we analyzed adipogenic differentiated cells by flow cytometry analysis, morphology, and expression level of adipogenic marker genes. During adipogenic culture, cell morphology of old and young ASCs changed. Two days after adipogenic induction, cell morphology changed from spindle shaped to star shaped. After 7 days of adipogenic induction, first adipocytes could be observed (data not shown). Within the next 7 days of adipogenic culture, both the total number of adipocytes and the amount of lipid droplets inside the cells increased in young and old differentiating cells (Fig. 3A).
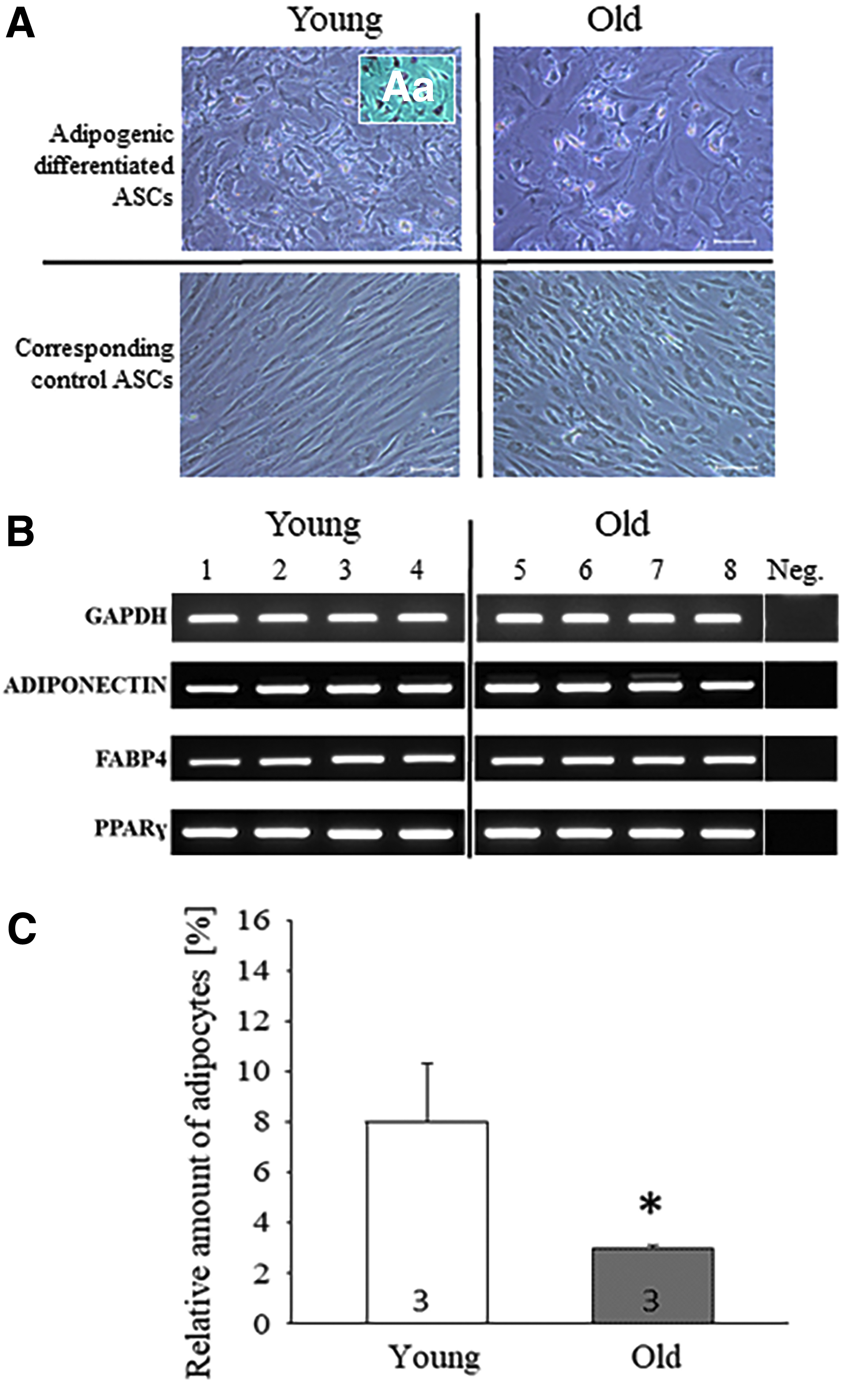
Adipogenic differentiation of young and old VASCs. Representative images of adipogenic differentiated young and old VASCs, [
Corresponding control VASCs without induction medium did not show morphological changes (Fig. 3A). We used Oil Red O, a fat-soluble dye, to stain neutral triglycerides and lipids in adipocytes (Fig. 3Aa). Adipogenic differentiated old VASCs had decreased average mean of adipocytes (3% ± 0.1%) compared with young adipogenic differentiated VASCs (8% ± 2.3%; p ≤ 0.05) as observed by flow cytometry analysis (Fig. 3B). The expression of adipogenic markers PPARγ, FABP4, and ADIPONECTIN, was present in differentiated cells (Fig. 3C).
Changes of marker gene expression in old VASCs
The mRNA expression of ATG7, an autophagy marker in tissues and MSCs was reduced in VASCs of old individuals compared with young VASCs. The mRNA expression of all other investigated aging markers, APOE, FGF2, PTEN, and SIRT1, was not changed (Fig. 4 and Table 3). The protein amounts of APOE, ATG7, and PTEN were increased in old VASCs (Fig. 5)

Analysis of aging markers–mRNA level. Age-related effects of absolute mRNA expression of APOE, ATG7, FGF2, PTEN, SIRT1 in young and old VASCs and SASCs (white bars: young; black/gray bars: old). Results are mean ± SEM from three independent experiments with six to eight rabbits/group. *p < 0.05 compared with young VASCs or young SASCs. SASC, subcutaneous adipose stem/stromal cell.

Analysis of aging markers–protein level. Age-related effects of relative protein amount of APOE, ATG7, FGF2, PTEN, SIRT1 in young and old VASCs and SASCs (
The mRNA Expression of Young and Old Rabbit Visceral Adipose-Derived Stem/Stromal Cells and Subcutaneous Adipose Stem/Stromal Cell [per 10x GAPDH Molecules]
Data are presented as mean ± SEM from three independent experiments (n = 6–8).
p < 0.05 compared with untreated controls.
SASC, subcutaneous adipose stem/stromal cell; VASC, visceral adult stem/stromal cell.
Changes of marker gene expression in old SASCs
The mRNA concentration of the aging makers, APOE, FGF2, and SIRT1, were significantly decreased in old SASCs (Fig. 4). No significant changes of mRNA expression in old SASCs could be observed for PTEN and FGF2 (Fig. 4 and Table 3). In contrast, protein amount of all aging markers was not changed in old SACS significantly (Fig. 5). The low molecule-isoform of FGF2/18 was slightly, but not significantly reduced in old SACSs (Fig. 5).
Discussion
Adult stem cells in almost all tissues operate as regenerators and health keepers against extrinsic and intrinsic influences over lifetime in humans and animals. During life, they undergo aging processes, which can result in loss of stem cell function. Aging mechanisms include changes of adult stem cells, their stem cell niche, and their surrounding environment leading to lesser stem cell power with declined potential of tissue regeneration and tissue repair (Rando, 2006). To investigate underlying mechanisms and age-related diseases, adult stem cells mimic an auspicious instrument. MSCs have the capability to differentiate into adipocytes, chondrocytes, and osteocytes (Pittenger et al., 1999; Zhu et al., 2009).
Many studies on human MSCs had demonstrated the effect of donor age on lineage-specific differentiation. In 2009, Zhu and colleagues showed that osteogenic differentiation potential depends on donor age. However, mechanisms behind aging are complex, controversially discussed, and until now not fully understood.
Rabbits are phylogenetically more similar to humans than to rodents like rats and mice (Graur et al., 1996). The rabbit is a proven animal model for research on age-related metabolic diseases like hyperglycemia and hyperlipidemia (Fischer et al., 2017; Roos et al., 2005; Weekers et al., 2003).
Isolation and characterization of young and old rabbit VASCs
Based on the isolation protocol of Zhu et al. (2013) for human ASCs, the isolation and primary culture of ASCs from young and old female NZW rabbits was established. To prove the MSC properties, we analyzed the cell morphology, expression level of surface antigens, and multi/pluripotency markers and differentiated cells into adipocytes as mentioned and compared it with human data (Baer and Geiger, 2012; Dominici et al., 2006; Zhu et al., 2013). The International Society for Cellular Therapy (ISCT) defined human MSCs as plastic adherent under normal cell culture conditions, multipotent, and with a cell surface immunophenotype positive for CD105, CD90, and CD73 and negative for CD34, CD45, and CD14 (Dominici et al., 2006).
The expression of the multipotency markers, CHD1, KAT6A, Klf4, MYC, and REST (Bernard et al., 1983; Gaspar-Maia et al., 2009; Li et al., 2005; Ooi and Wood, 2007; Sheikh et al., 2015), and no expression of the pluripotency genes, NANOG, SOX2, and OCT4 (Chambers et al., 2003; Schöler et al., 1989; Zhang and Cui, 2014), in rabbit VASCs confirm the multipotency potential of the isolated VASCs from young and old rabbits (Fig. 2C).
Our data are in accordance to observations in human ASCs. Zhu et al. (2009) isolated and cultured human ASCs. These cells showed a typical MSC morphology with no differences between young (20 years) and old (58 years) donors. Rabbit VASCs also showed an MSC characteristic morphology with no visual differences between young and old rabbits (Fig. 1B). In addition, VASCs bear Vimentin and Colagenase1A1 as typical MSC markers (Fig. 2A).
In most studies, PDT of human ASCs did not change with higher age (Chen et al., 2012; Ding et al., 2013; Zhu et al., 2009). Furthermore, human SASCs from omental adipose tissue proliferate faster than VASCs (Van Harmelen et al., 2004). In accordance to human data, the PDT of old rabbit VASCs was not different to young VASCs (Fig. 1C).
The negative surface markers, CD14, CD73, and CD90 of rabbit VASCs fits into the literature of human ASCs (Zhu et al., 2013). Bakhtina et al. (2014) confirmed CD73 and CD90 as negative markers in rabbits (Table 4). The expression profile of positive markers in our rabbit model was strong positive for CD105, which correlates with the literature of human ASCs (Dominici et al., 2006; Zhu et al., 2013). In humans, CD44 is a positive marker and CD45 a negative marker (Zhu et al., 2013). In contrast, both markers are slightly expressed in rabbits (30%). Therefore, these markers are not suitable for the characterization of rabbit ASCs. Taken together, the isolated rabbit ASCs showed typical MSC characteristics with no significant differences between young and old donors.
Expression Profile of Surface Antigens of Mesenchymal Stem Cells
Reduced adipogenic potential of old VASCs
It has been postulated that a decline in the capability of preadipocytes to differentiate into mature adipocytes is responsible for body fat reduction in the elderly (Cartwright et al., 2007; Kirkland and Dobson, 1997; Kirkland et al., 1990). In the present study, adipogenic differentiation of young and old VASCs was achieved by insulin, dexamethasone, indomethacin, BMP4, and IBMX. Both young and old VASCs could be differentiated into adipocytes successfully (Fig. 3). Expression of the adipogenic markers, adiponectin, FABP4, and PPARγ, was detected by RT-PCR in all probes of young and old VASCs (Fig. 3B). Flow cytometry analysis of adipogenic differentiated cells revealed a significant decrease of adipocytes in the old group (8%) compared with young donors (3%, Fig. 3C).
Xu et al. made a similar observation in 2015. They showed a decrease of adipogenesis rate of in vitro aged human fat progenitor cells. In contrast, Zhu et al. (2009) observed no differences in adipogenic differentiation capacity of in vivo aged human ASCs (young: 20 years, old: 58 years). However, they showed a high variability of adipogenic differentiation rate (from 58% to 4%). Nevertheless, new techniques as the 3D culture system may be enhanced and align the yield of differentiated adipocytes (Miyamoto et al., 2016), and standardized methods on ASC research and cell culture systems reveal new light inside the process of MSC aging.
Significant changes of aging markers in old VASCs and SASCs
Throughout lifetime, tissue-specific stem cells undergo aged-related changes in many intrinsic and extrinsic pathways. To further investigate this complex interrelationship during aging, we analyzed a marker panel, including APOE, ATG7, FGF2, PTEN, and SIRT1. This panel is reviewed mainly in Oh et al. (2014), and encompasses molecular pathways of autophagy (ATG7), cell proliferation (FGF2), cell cycle, self-renewal (PTEN), cellular senescence (SIRT1) (Oh et al., 2014), and additionally APOE, a metabolic sensor for longevity (Noordam et al., 2016).
APOE: metabolic sensor for longevity
The protein amount of APOE showed significant increase in old VASCs, but not significant in SASCs. APOE, a lipid-binding Apolipoprotein plays a pivotal role in lipid metabolism, atherosclerotic cardiovascular disease, Alzheimer diseases, and longevity (Ang et al. 2008; Beekman et al., 2013; Corella and Ordovás, 2013; Liu et al., 2014). Furthermore, APOE acts as a transcription factor for regulating programmed cell death, sirtuins, and aging (Theenadakara et al., 2016). APOE−/− mice showed enhanced hypercholesterolemia-induced oxidative stress, which goes along with accelerated aging of HSCs (Tie et al., 2014).
It might be possible that in old rabbit VASCs the upregulation of APOE protects cells from accelerated aging mechanisms. To clarify APOE function and regulation in old VASCs and SASCs, additional experiments are needed. Particularly the mRNA of APOE was significantly reduced in old SASCs, indicating a possible negative feedback regulation. In contrast to old VASCs, we could not detect significant changes of APOE protein in SASCs, demonstrating their fat tissue function and metabolic activity on different fat depot sites.
ATG7: one key regulator of autophagy
The protein amount of ATG7 was significantly increased in old VASCs, but not in SASCs. The process of autophagy is essential to degrade old, defective cell fragments to maintain proteostasis (Lapierre et al., 2015; Mortensen et al., 2011). Several experiments indicate that changes in autophagy activity go along with age-dependent pathologies (Kroemer and Levine, 2008). Zheng et al. (2014) showed in in vitro aged, senescent rat MSCs, an activated autophagy and elevated autophagy-related proteins, such as ATG7. Similar observations were made in senescent endothelial cells (Kurz et al., 2000) and fibroblasts (Gerland et al., 2003).
In contrast, a deletion of ATG7 gene induces reactive oxygen species (ROS) levels and depletes hematopoietic stem cells (HSCs) (Mortensen et al., 2011). However, an elevation of ATG7 protein in old VASCs may provide a survival effect under aging. The inverse correlation of ATG7-RNA and protein levels may appear by posttranscriptional and posttranslational mechanisms. In stress conditions ATG7-RNA expression has been shown directly regulated by heat shock protein 1 (Desai et al., 2013) and inversely correlated with microRNA-17 expression (Comincini et al., 2013). The protein expression was downregulated by miRNA-137 (Zeng et al., 2015). We suppose that ATG7 protein stabilization could affect ATG7-RNA availability by a direct or indirect mechanism.
FGF2: modulator of self-renewal
The mRNA expression of FGF2 was significantly decreased in old SASCs, but not in VASCs. Hurley et al. (2016) postulate that a decrease in FGF2 expression could contribute to age-related cell function of human mesenchymal-derived progenitor cells (hMDPCs). In vitro FGF2 stimulation of young human, mice, and rabbit bone marrow-derived MSCs result in an elevated cell proliferation and an expanded lifetime (Choi et al., 2008; Eom et al., 2014; Tsutsumi et al., 2001).
Compared with the decrease of FGF2 expression in old SASCs, a slight, (p = 0.07) decrease of protein amount in old SASCs was observed for the low molecular weight FGF2/18 form. In contrast, significant changes in protein and mRNA level could not be detected in old VASCs. FGF2 signaling in aging ASCs depends on donor sites, and the use of FGF2 as a strong aging marker in old rabbit ASCs has to be investigated further in detail.
PTEN: an inhibitor of PI3K/AKT cascade
The protein amount of PTEN in old VASCs was increased, but not in SASCs. PTEN acts as an inhibitor of the PI3K/AKT pathway, regulates cell proliferation, survival, and migration, and is a tumor suppressor (Panigrahi et al., 2004; Zhang et al., 2006). Confirming to our date Rocha et al. (2014) documented an age-related increase of PTEN in healthy rats. In addition, PI3K/AKT inhibition blocks adipogenic differentiation of brown preadipocytes (Fasshauer et al., 2000) and 3T3L1 preadipocytes (Zhang et al., 2009). Thus, the elevated PTEN protein we observed maybe one possible reason for the decrease in adipogenic differentiation in old VASCs.
SIRT1: delays cellular senescence
The mRNA expression of SIRT1 was significantly decreased in old SACSs, but not in old VASCs. Protein levels were not affected by age. In general, SIRT1 levels are decreased during aging (Yuan et al., 2016). Recently, Khanh et al. (2018) showed that a reduction of SIRT1 is responsible for the induction of senescence in old human ASCs and for a lesser adipogenic differentiation of human beige adipocytes, too. In contrast, they demonstrate that an overexpression of SIRT1 prevents old ASCs for starting cellular senescence. Nevertheless, we only detect a decrease in mRNA of old SASCs and protein levels were unaffected by age.
Are rabbit ASCs able to struggle for longevity to maintain stem cell plasticity?
We showed that aged rabbit ASCs changed the expression of markers that are crucial for the biology and plasticity of stem cell aging. We postulate that ASCs are able to compensate extrinsic and intrinsic aging to maintain their stem cell plasticity for balancing normal tissue and organ functions. However, this capability seems to be limited. For example, aged ASCs had a lower adipogenic differentiation rate. To clarify the impact of healthy aging on adipocyte tissue function in detail, more in vitro experiments are needed.
VASCs versus SASCs: why do they act differently by age?
Table 5 shows that the mRNA level and protein amounts depend on the fat depot site. Cells from both locations showed significant changes in stem cell aging markers, but in a different manner. Old VASCs showed significant changes of APOE, ATG7, and PTEN levels, whereas old SASCs showed changes of APOE, FGF2, and SIRT1 levels. Aging causes a decrease of total body fat and a redistribution from fat depots to other sites (Kirkland et al., 2002).
Expression Profile of Old Rabbit Visceral Adipose-Derived Stem/Stromal Cells and Subcutaneous Adipose Stem/Stromal Cells
P, Protein; R, mRNA; +, elevated; −, decreased; n.c., not changed.
In addition, the amount of subcutaneous fat mass decreases first, followed much later by a loss of visceral fat mass. This finally results in an accumulation of visceral fat and ectopic fat in bone marrow, muscle, and liver (Cartwright et al., 2007). The principal mechanism behind is until now not fully understood. Taken together, our data indicate that the observed differences in the expression profile of the aging markers in old ASCs are one reason for the obviously different age-related shift of subcutaneous to visceral fat depot.
In summary, we showed a decrease of adipogenic differentiation rate in ASCs of visceral adipose tissue from old female NZW rabbits. The loss of adipogenic differentiation plasticity is age related and it is associated with altered mRNA and protein levels of specific aging markers in the undifferentiated stage in ASCs of visceral adipose tissue from old female NZW rabbits. The findings suggest that the reason for the differentiation dysfunction of old adult stem cells may appear already in the undifferentiated stage of old visceral rabbit ASCs and this may contribute to a less tissue maintenance, regeneration, and cause typical disorders later in life.
Footnotes
Acknowledgments
This work was supported by DFG GRK 2155 ProMoAge and Wilhelm Roux Program, MLU Faculty of Medicine. The authors would like to thank Sabine Schroetter, Michaela Kirstein, and Bernadette Harwardt.
Major Findings
Rabbit ASCs showed age- and source-specific alterations due to the age of the donor. Stem cell plasticity was altered with age as shown by reduced visceral adipogenic differentiation capacity.
Author Disclosure Statement
The authors declare they have no financial conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.