
Abstract
Abstract
Maturation conditions and oocytes quality have substantial roles on developmental competence of unreconstructed or reconstructed oocytes. Cloning has been reported successfully with low efficiency through embryonic or somatic nuclear transfer into enucleated metaphase II oocytes. It has been suggested that introducing embryonic or somatic nucleus to cytoplast at earlier stage might improve reprogramming of the introduced nucleus. In addition, the synchronization between the donor nucleus and recipient cytoplasts might effect on reprogramming and further embryonic development. Therefore, the question arises; does the cytoplast of germinal vesicle (GV) oocyte containing nuclear sap improve developmental competence upon reconstruction with embryonic/somatic nucleus compared with MII cytoplast. It has been indicated that GV material is essential for remodeling of sperm or embryonic or somatic nucleus in GV oocyte cytoplast and their further embryonic developmental competence. GV cytoplasts could be obtained through micromanipulation. Different micromanipulation techniques of immature oocytes at different stages were adapted in addition to introducing donor nuclei at G0/G1, S and G2/M phase, and enucleolation technique as well. Upon micromanipulation, it could obtain GV cytoplasts; cumulus-free and without GV material, cumulus-complexes and without GV material, cumulus-free and with GV material, and cumulus-complexes and with GV material in addition to enucleolated GV oocytes. Therefore, this short review will address briefly the importance of maturation conditions, cumulus cells, oocyte quality, the techniques of enucleation GV oocyte, the cell cycle stage of the introduced donor cell, or nucleus for oocyte maturation and embryo development.
Introduction
It is well-known that developmental competence of in vivo MII oocytes is better than those produced in vitro due to maturation conditions and quality of oocytes (Egashira et al., 2019). In vivo matured oocytes were ovulated from large and ovulatory follicle, whereas in vitro matured oocytes were aspirated from follicles that vary in diameters and quality and mature in culture conditions (Mohammed et al., 2005). Therefore, the effects of in vitro maturation or culture conditions and quality of oocytes were presented first because of their substantial roles on developmental competence of oocytes and resulting embryos.
Production of embryos in vitro includes in vitro maturation, in vitro fertilization, and in vitro culture of the fertilized oocytes. In addition, the success in assisted reproductive technology (ART) depends on several factors concerning maturation of germinal vesicle (GV) oocyte cytoplast (Wani et al., 2018a; Zhang and Liu, 2015), GV material (Bui et al., 2008; Mohammed et al., 2008, 2010), cumulus cells surrounding the GV oocytes, and cell cycle stage of donor nuclei (Mohammed et al., 2010). Wright (2018) suggested that cloning animals in the year 2070 will be used for conservation, education, and tourism.
The success in embryo and fetal development and pregnancy depends on maturation of both cytoplast and nucleus of oocyte (Kyogoku et al., 2018). For obtaining artificial gamete in ARTs, recipient cytoplast and donor nucleus is required. The interaction between the recipient cytoplast and the donor nucleus is considered one of the fundamental factors controlling development of the resulting embryos (Mohammed, 2006; Mohammed et al., 2008, 2010; Wani et al., 2018a). The resulting competent artificial gamete, which has the ability to develop to embryo and fetus, can be used for treatment of reproductive disorders or embryonic or somatic cloning.
Development of reconstructed MII oocytes is too low due to poor reprogramming of the introduced nucleus/cell. Hence, the suggestion was to introduce the donor nucleus/cell to oocyte cytoplast at earlier stage that is, GV cytoplast to help in embryonic/somatic nuclear reprogramming. Such reconstructed GV cytoplasts or oocytes are expected to solve the abnormalities of male sperm. In case of disability of sperm to fertilize oocytes or abnormalities, primary or secondary spermatogonia can be used for fertilizing the oocytes (Ogura et al., 1998). One and two meiotic divisions of primary and secondary spermatogonia, respectively, are expected in GV cytoplasts.
Therefore, obtaining GV cytoplast is required in basic research for the investigation of cell biology and reprogramming of donor nucleus or cell. Trials were carried for enucleation of immature oocytes at different stages in addition to introducing donor cells at stages of G0/G1 or S or G2/M phase (Grabarek et al., 2004; Mohammed, 2006; Mohammed et al., 2008, 2010) for exploring meiotic division and further developmental competence of embryos (Table 1 and Fig. 1). The enucleation techniques of GV oocytes include removal of the whole GV nucleus (complete enucleation [CE] technique) or enucleation with removal of only the nucleus membrane and chromatin, but leaving in cytoplasm the nuclear sap and nucleolus (selective enucleation [SE] technique). Complete and SE techniques of GV carry out without or with the presence of cumulus cells.

Factors affecting developmental competence of unreconstructed and reconstructed germinal vesicle oocytes.
Effects of Types of Recipient Cytoplasts and Donor Nuclei and Their Cell Cycle Stages Used in Reconstructed Artificial Oocytes
Therefore, the effects of cumulus cells as well as nuclear components can be tested through such techniques. In addition, enucleolation technique is adapted in some species as pig and mice (Fulka et al., 2003; Mohammed, 2006; Mohammed et al., 2008) to explore the importance of nucleolus on oocyte maturation and embryo development. It has been indicated that the nucleolus is not necessary for maturation of oocyte; however, it is necessary for pronuclear nucleoli formation after fertilization or activation and is essential for further embryonic development (Mohammed et al., 2008; Ogushi et al., 2008). This enables to study nucleolus transfer from donor fully grown GV oocyte to recipient GV oocyte containing abnormal nucleolus to rescue infertility due to abnormal nucleolus.
The application of such ARTs might enable to produce more offspring of prepubertal parents and treat infertility in animals and human as well. The success of technologies might shorten the interval between generations, treat of reproductive diseases and conserve of endangered mammalian species. Therefore, the technologies have beneficial and potential roles in reproduction and expected full-term deliveries upon embryo transfer. Because the developmental competence of either unreconstructed or reconstructed oocytes is governed by in vitro culture conditions and characteristics of GV oocytes, they were included in this article ahead of oocytes micromanipulation.
Therefore, this short review will address briefly factors affecting in vitro developmental competence of unreconstructed or reconstructed GV oocytes as maturation conditions, cumulus cells, oocytes quality, micromanipulation techniques GV oocyte, the cell cycle stage of the introduced donor cell or nucleus, and their importance for fertility and reprogramming.
Maturation Conditions
There are several factors as maturation conditions affecting in vitro developmental competence of unreconstructed (intact) or reconstructed GV oocytes. In general, as the time of oocyte maturation in vitro increases the developmental competence of oocyte decreases as in swine and camel oocytes. Duration of oocyte maturation in vitro from GV to metaphase stage lasts in most species from 17 to 48 hours (Luo et al., 2018; Mohammed et al., 2005, 2008, 2010; Moulavi and Hosseini, 2018). Several factors of maturation conditions in vitro such as temperature, humidity, oxygen tension, and maturation medium have been confirmed to effect on oocyte maturation and further embryo development of either unreconstructed or reconstructed oocytes (Kovacovicova et al., 2016; Mohammed et al., 2005, 2008, 2010; Zheng et al., 2005).
Trials were undertaken to study the effects of undefined and defined media on oocyte maturation and subsequent developmental competence of embryos in different species (Abdoon et al., 2018; Madkour et al., 2018; Mohammed et al., 2005; Spacek and Carnevale, 2018).
Oocyte maturation in vitro is influenced by the addition of fetal calf serum (Moulavi and Hosseini, 2018) or follicular fluid to maturation media (Valckx et al., 2015). The effect of FF is dependent on the size of follicle from which the follicular fluid was obtained (Oberlender et al., 2013), timing of follicular fluid collection either before or after LH surge (Romero-Arredondo and Seidel, 1994), and percentage of follicular fluid added to maturation media (Mohammed et al., 2005) in different species. Coelho et al. (2014) investigated maturation and embryo development upon adding bovine follicular fluid to maturation medium (0%, 25%, 50%, 75%, and 100%), which collected from follicle with diameter 8–15 mm.
The authors found that higher follicular fluid concentration (75%–100% FF) slowed meiotic progression and cortical granule migration. It could be concluded that follicular fluid obtained from smaller follicles has inhibitory effect on oocytes' maturation than that obtained from larger ones. Other supplements added to maturation medium might affect oocyte maturation and developmental competence as FSH, LH, and estrogen hormones (Moulavi and Hosseini, 2018), others (Akar et al., 2018). Therefore, the trials to improve culture conditions for in vitro embryo production are still required as quality of oocytes (Maylinda et al., 2018; Mohammed, 2018; Raj et al., 2018).
Cumulus Cells
Cumulus cells are a group of cells that surround the oocyte and communicate through gap junctions. It has been thought that gap junctions between cumulus cells and oocytes are essential for oocyte growth and maturation and further embryonic development. Presence of cumulus cells around the GV oocytes (cumulus oocytes complexes, COCs) or inclusion of cumulus cells to maturation medium (Lee et al., 2018; Mohammed, 2006; 2008; Mohammed et al., 2008, 2010) during oocyte maturation in vitro effects on maturation rate and development of the resulting embryos. Cumulus cells are essential for transfer of some nutrients to the oocytes such as amino acids.
Amino acid uptake is first by cumulus cells and transferred thereafter to the oocyte via gap junctions. There are several roles of amino acids in oocytes and the resulting embryos as protein synthesis, energy sources (Rieger, 1992), intracellular buffers (Edwards et al., 1998), osmolytes (Dawson et al., 1998), heavy metal chelators (Bavister, 1995), and antioxidant compounds (Guérin et al., 2001).
Follicular fluid that surrounds the oocyte is synthesized from secretions of cumulus cells and from a transudate of blood plasma. Amino acid composition of follicular fluid was used as predictors of in vitro embryo development. The amino acid composition of follicular fluid was associated with morphological quality of COC and with postfertilization embryo development to the blastocyst stage (Sinclair et al., 2008). It has been found that inclusion of specific amino acids in culture media facilitates embryo hatching of embryos in some species (Liu and Foote, 1995; Pinyopummintr and Bavister, 1996), helping to alleviate cultured-induced arrest. Therefore, cumulus cells or their secretions (FF) can improve cytoplasmic maturation of oocytes (Ikeda and Yamada, 2014).
Oocyte Quality
The success in oocyte maturation and further embryo development depends on oocyte quality. There are several factors affecting oocyte quality, including follicle size (Mohammed et al., 2019a; Shabankareh et al., 2014; Zhou et al., 2005) and nutrition (Mohammed and Al-Suwaiegh, 2019; Mohammed et al., 2011, 2019b; Mohammed, 2018). In addition, developmental competence of in vivo matured oocytes is higher than in vitro matured oocytes. This might be related to nucleus and/or cytoplasmic factors. There are several factors affecting on oocytes maturation in vitro and their further developmental competence; such as culture and coculture conditions (Hoseini et al., 2016; Mohammed et al., 2005; Zhou et al., 2005), cumulus cells surrounding the oocytes (Mohammed et al., 2005; 2008; 2010; Mohammed, 2006), and diameter of ovarian follicles from which the oocytes were collected (Shabankareh et al., 2014; Zhou et al., 2005).
In addition, the components of maturation medium (Mohammed et al., 2005) were occasionally effected on maturation where segregation of the chromatids and polar body extrusion seemed to be impaired in the presence of urea (De wit et al., 2001; Kowsar et al., 2018). Our work (Mohammed et al., 2005) indicated change in timing of embryo cleavage and developmental competence upon supplementation of follicular fluid to the maturation medium during oocytes maturation. Studies have indicated the increase of some genes expression during the stages of oocyte maturity from GV to meiosis II (MII) stage (Assou et al., 2006; Wells et al., 2005).
Oocyte nucleus/cytoplasm transfer techniques have been used for treatment of abnormal oocytes due to cytoplasmic and/or nucleus reasons or to increase their developmental competence (Hoseini et al., 2016; Mohammed et al., 2008, 2010). Hoseini et al. (2016) concluded that the technique of cytoplasm transfer is not effective in cytoplasmic maturity of the recipient GV oocytes, whereas the presence of cumulus cells during oocyte maturation or GV nucleus transfer is effective in increasing the developmental competence of the resulting embryos (Mohammed et al., 2005, 2006, 2008, 2010).
Manipulation of Oocytes and Reconstruction
Obtaining the recipient cytoplasts of oocytes necessitates the removal of the nuclear material from oocytes. The recipient cytoplasts at earlier stages of maturation (GV, ProMI & MI) are a useful material for basic research in cell biology and reprogramming of the introduced nucleus or cell (Van Heyman, 2005). It may solve the problem of infertility with advancing maternal age (Hassold and Chiu, 1985) or ensure the developmental competence in case of scarce resources. It is well known that there is a clear inverse relationship of fertility with advancing maternal age. In addition, oocytes' developmental competence of young animals were lower than that of adult ones. Therefore, transfer of nuclei from nongrowing, growing, or aging GV oocytes into fully grown GV cytoplasts might rescue the developmental incompetency due to ooplasmic or nuclear nature (Bao et al., 2003).
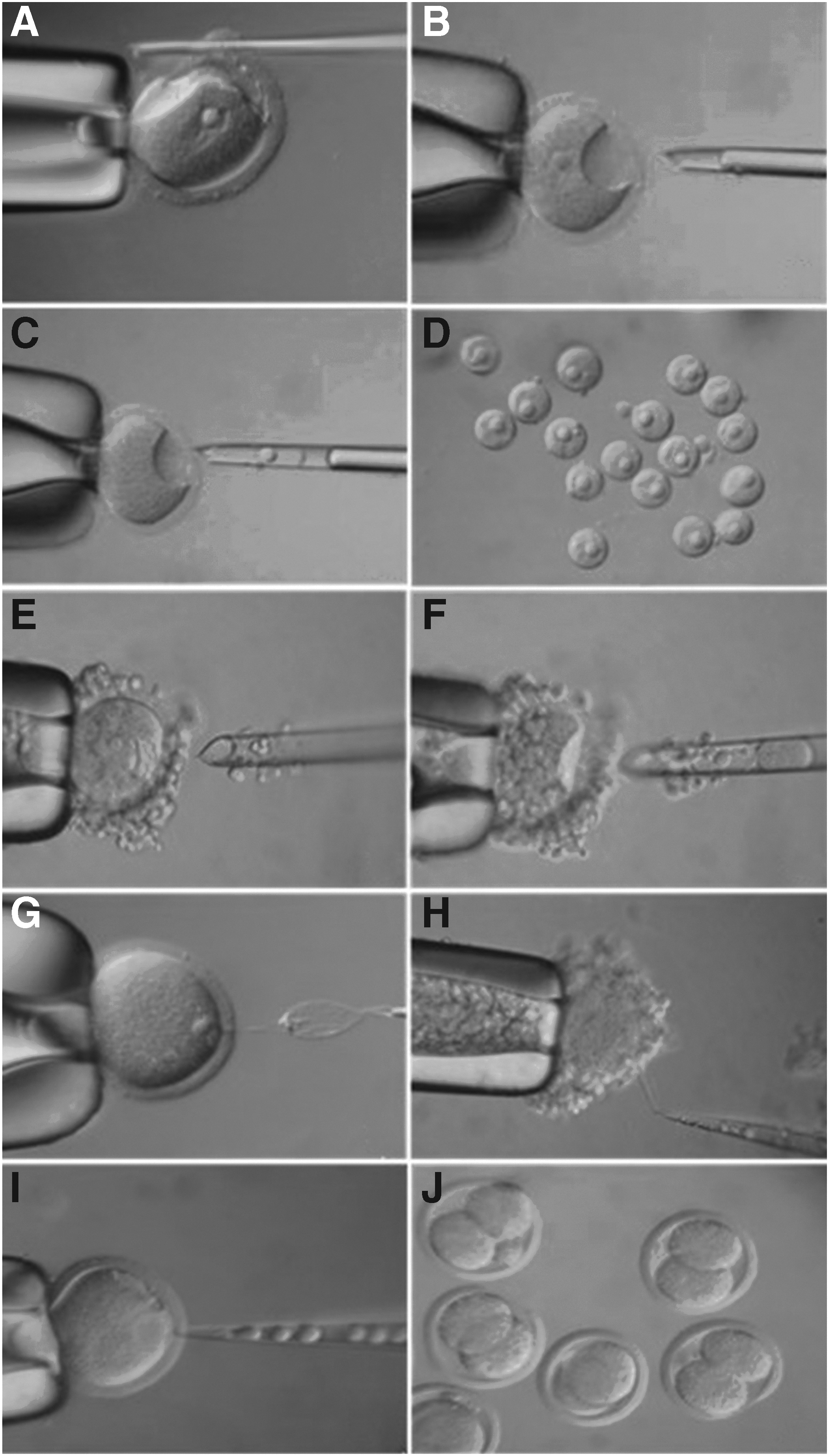
The nucleus of GV oocytes of mammals is visible in some species as rodents, rabbit, and human or invisible as ruminants due to lipid droplets. The GV oocytes with visible nucleus are directly enucleated, whereas the GV oocytes with invisible nucleus required visualizing the nucleus through staining (Hoechst 33342) or ultracentrifugation at 10,000–12,000 rpm for 10 minutes followed by enucleation. Four techniques were carried out on GV oocytes to explore the meiotic maturation and further developmental competence or embryos; CE of denuded or cumulus-enclosed GV oocytes and SE of denuded or cumulus-enclosed GV oocytes (Fig. 2). In addition, enucleolation technique was adapted in some species (Fulka et al., 2003; Mohammed, 2006; Mohammed et al., 2008, 2010). Such techniques enable to explore the effects of cumulus cells, nuclear sap, and nucleolus on oocytes maturation and developmental competence of embryos.
Enucleation and enucleolation of GV oocytes
Indirect enucleation of GV oocyte
Enucleation of GV oocytes is difficult due to the sensitivity of the plasma membrane, the large diameter of GV nucleus, and the close contact of zona pellucida (ZP) and vitelline membrane. Therefore, indirect enucleation of GV oocytes has been adapted earlier (Li et al., 2001) by cutting the ZP and aspiration of GV nucleus from the oocyte through the slit in the ZP (Fig. 2A).
Direct CE of GV oocyte
A direct enucleation method of GV oocytes was adapted by preincubation and enucleation in M2 medium containing 16 mM glucose, 1 μg CD, 0.25 μg nocodazole, and 0.2 mM dbcAMP (Grabarek et al., 2004). This enucleation solution enlarged the space between ZP and perivitelline membrane of GV oocytes, which in turn allowed to insert the enucleation pipette without the need to cut the ZP prior (Fig. 2B, C). Our protocol (Mohammed, 2006; Mohammed et al., 2008, 2010), in which the GV oocytes were preincubated and enucleated in M2 medium supplemented with CB (10 μg), nocodazole (0.25 μg), and dbcAMP (0.2 mM), resulted in comparable survival rate upon enucleation and higher survival rate after fusing the introduced cells compared with the previous study.
In addition, the technique of CE was performed for cumulus-enclosed GV oocytes as the same way in cumulus-free GV oocytes (Fig. 2E, F). This enables to investigate the potential role of cumulus cell on oocyte maturation and developmental competence. The obtained GV nuclei (Fig. 2D) could be transferred to other cytoplasts for different purposes as treatment of infertility.
Direct SE of GV oocyte
The technique of SE is adapted to leave the nucleoli and nuclear sap in the cytoplast and remove only the nuclear envelope with the attached chromatin (Fig. 2G, H). The technique of SE was performed in the same way of cumulus-free and cumulus-enclosed GV oocytes (Mohammed, 2006; Mohammed et al., 2008) with visible GV nucleus. This technique enables to explore the potential role of GV material on oocyte maturation and developmental competence of embryos. The developmental competence of embryos was cleaved further to the blastocyst stage compared to those blocked at two-cell stage without GV material.
Enucleation of ProMI and MI oocytes
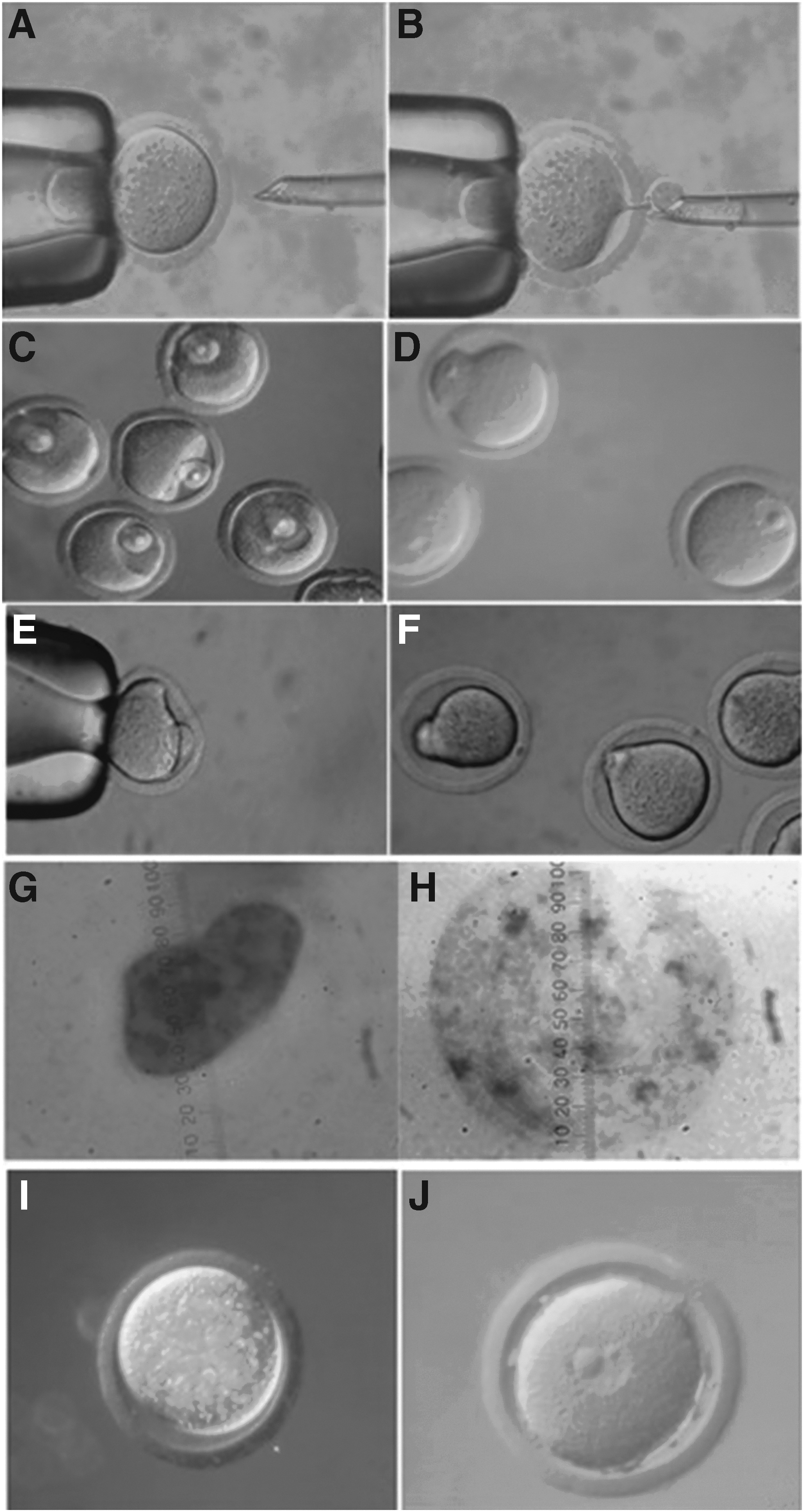
The ProMI and MI oocytes were incubated for 30 minutes in the enucleation medium supplemented with 2% sucrose to visualize the localization of nuclear material (Wang et al., 2001). The ProMI nuclear material or MI spindle after this treatment was visible as translucent spot in the cytoplast (Fig. 3A, B) and could be removed using enucleation pipette (10–15 μm). The cytoplasts of ProMI and MI oocytes contain unknown quantity of GV material compared with SE techniques in addition to the timing of maturation, which process in few hours.
Reconstruction of oocytes
Fusion of the introduced GV nucleus or cell
The GV nucleus or cell was introduced under the ZP (Fig. 3C, E). The GV nucleus-cytoplast or cell-cytoplast complexes were exposed to electrofusion. Fused pairs (Fig. 3D, F) were cultured for maturation and further developmental competence.
Enucleolation GV oocytes
Enucleolation of GV oocytes enables to explore the role of nucleolus on oocytes maturation and further developmental competence of the resulting embryos. The technique requires preincubation of GV oocytes for 30 minutes in M2 medium supplemented with dbcAMP (0.2 mM) to prevent GV breakdown. The tip diameter of the enucleolation micropipette used was ∼3 μm (Fig. 2I). The result indicated that nucleolus is dispensable for oocyte maturation, but is essential for further embryonic development where the embryos blocked at the two-cell stage in case the nucleolus removed from oocytes (Fig. 2J).
Embryonic Developmental Competence After Nuclear Transfer
The results indicated that GV material is essential for remodeling of sperm or embryonic or somatic nucleus in oocyte cytoplast and their further embryonic developmental competence (Mohammed 2006; Mohammed et al., 2008, 2010). The formed pronucleus in GV cytoplast without GV material seemed smaller and irregular in shape with no visible nucleolar material (Fig. 3G) versus that formed in GV cytoplast with GV material, which seemed regular in shape, larger, and contain nucleolar material (Fig. 3H). In addition, the sperm could not form paternal pronucleus (Fig. 3I) in matured GV cytoplast without GV material versus formation of paternal pronucleus in matured GV cytoplast with GV material (Fig. 3J).
Furthermore, presence of cumulus cells potentially increased the developmental competence of the resulting embryos (Mohammed 2006; Mohammed et al., 2008, 2010) where the cumulus cells is essential for passing nutrients to the oocyte cytoplasm as some of amino acids (L-alanine) and glucose. Numerous projections from the innermost cumulus cells penetrate the ZP and end on the oolemma with gap junctions. Although cumulus-enclosed oocytes, when cultured in medium containing only glucose as an energy source, resume meiosis, cumulus-free oocytes could not. This indicates that cumulus cells take up glucose and metabolize it to products necessary to support meiotic maturation (Leese and Barton, 1985). No pregnancies or offspring were confirmed up till now over reconstruction of GV cytoplasts either with embryonic or somatic nuclei.
On the contrary, cloned animals were obtained in different species over reconstruction of MII cytoplasts either with embryonic or somatic nuclei with efficiency lower than ten percent (Matoba and Zhang, 2018; Westhusin and Long, 2018; Xu et al., 2018) in addition to transgenic animals (Houdebine, 2018).
Donor Nuclei
The source, type, and treatment of donor cells influence in vivo and in vitro development of embryos cloned by somatic cell nuclear transfer in different species (Liu et al., 2018; Wani et al., 2018b). In addition, the interaction between the recipient cytoplasts and the donor nuclei is considered one of the fundamental questions in the technologies of assisted reproduction. Our work (Mohammed, 2006; Mohammed et al., 2008, 2010) indicated the synchronous between the donor nucleus and the recipient cytoplast is essential for proper maturation and developmental competence. Transferring donor nuclei at S-phase of cell cycle to GV cytoplast accelerated timing of polar body extrusion and disorganized the chromosomes in the spindle versus transferring donor nuclei at G0/G1 or M-phase of the cell cycle.
Kimura and Yanagimachi (1995) indicated delivery of normal mouse from oocyte injected with secondary spermatocyte nucleus. In addition, Ogura et al. (1998) indicated obtaining normal mice from metaphase I oocyte fertilized with primary spermatocyte. Furthermore, Sasagawa et al. (1998) indicated two meiotic divisions within the oocyte cytoplast of mouse primary spermatocytes. In our conclusion, the GV cytoplast with nuclear material might give the introduced growing GV nucleus or primary and secondary spermatocytes the full time for maturation and consequently organization of chromosomes on the spindle. In addition, the presence of nuclear material, including nuclear sap and nucleolus, will potentially help the formation of proper nucleus and support the further developmental competence of embryos.
Conclusion
GV materials, including nuclear sap and nucleolus, are essential for remodeling of sperm or germ or somatic or embryonic nucleus in oocyte cytoplast. In addition, cumulus cells improved the developmental competence of matured oocytes. The techniques of CE, enucleolation, SE of denuded, and cumulus-enclosed GV oocytes provide materials for studying reprogramming of germ or embryonic or somatic donor nuclei and improving the developmental competence of reconstructed oocytes or artificial gametes. SE technique yielded cytoplasts with high survival rate and developmental competence.
Transfer of male or female germ nuclei ‘at least’ into selective GV cytoplasts might enable creating new oocytes containing the haploid and introduced nuclear genome through meiotic maturation. Such new oocytes could subsequently be activated or fertilized. SE of GV oocytes has provided a new alternative technique to leave nuclear sap and nucleolus in the cytoplast compared to CE, which support maturation of oocytes, preimplantation, and probably postimplantation development of embryos. Such techniques might be helpful in epigenetic reprogramming and treatment of infertility in human.
Footnotes
Acknowledgment
The authors thank and acknowledge Deanship of Scientific Research, King Faisal University, Saudi Arabia for the financial support under Nasher track (Grant No. 186153).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.