
Abstract
This study aims to understand the possible effects of TET2 (ten–eleven translocation 2) on the osteogenic differentiation of human adipose-derived mesenchymal stem cells (ADSCs). The human ADSCs were transfected with TET2 siRNAs. The osteogenesis-related genes were detected by quantitative real-time reverse transcription PCR (qRT-PCR), and the osteogenic differentiation was evaluated by alkaline phosphatase (ALP) staining and Alizarin Red staining. TET2 and 5-hydroxymethylcytosine (5hmC) expressions were determined by western blotting and immunofluorescence staining. Meanwhile, wild-type (WT) and TET2-deficient (TET2−/−) mice were selected to observe the alteration of biological characteristics in vivo. TET2 was significantly upregulated along with the osteogenic differentiation of human ADSCs. Compared with Blank group, TET2 siRNA-3 group showed apparent reductions in TET2, 5hmC, and osteogenesis-related genes, as well as decreases in mineralized nodules, ALP activity, and cell growth (all p < 0.05). Besides, Tet2−/− mice had shorter femoral length, lower bone mineral density, and reduced bone volume to total volume (BV/TV) ratio relevant to WT mice; and meanwhile, the percentage of TUNEL-positive chondrocytes increased significantly with the decreased total collagen-positive area, and the distance between two markers of calcein narrowed with declined bone formation rate (BFR) and mineral apposition rate (all p < 0.05). Furthermore, Toluidine Blue staining presented the appreciable decrease of BFR/bone surface (BS) ratio, BFR/BV ratio, osteoblast number over bone perimeter (N.Oc/B.Pm), and osteoblast surface (Ob.S)/BS in Tet2−/− mice (all p < 0.05). Taken together, TET2 upregulation was observed during the osteogenic differentiation of ADSCs, whereas TET2 inhibition may lead to reductions of osteogenesis-related genes and downexpression of 5hmC, which eventually plays a negative role in osteoporosis.
Introduction
Osteoporosis, a systemic bone disease featured by decreased bone mass and damaged bone microstructures, is greatly affecting the life quality of patients because of its high incidence rate and serious complications (Lee et al., 2016). The osteoporotic fracture would usually lead to the decreases in bone healing and bone regeneration, ultimately resulting in delayed healing, bone nonunion, and bone defect, which is a difficult medical problem puzzling the clinical work (Kolios et al., 2010). According to the findings by some previous studies, bone transplantation does have certain effects on repairing bone defect caused by osteoporosis, but this method still has some great limitations (Cao et al., 2012; Diwan et al., 2013).
As a prominent strategy in regenerative medicine, tissue engineering has become one of the most promising therapy for the bone defect repairing in the recent years (Keeney et al., 2016; Lappalainen et al., 2016), whereas adipose-derived mesenchymal stem cells (ADSCs), the multipotent cells with high self-renewal ability routinely isolated from adipose tissue, hold great promise in potential treatments for tissue regeneration (Narai et al., 2015; Zidi and Allaire, 2015). As a matter of fact, ADSCs can be induced to differentiate into osteoblasts under the stimulation of certain culture conditions (Liu et al., 2011). Owing to the features of abundant sources, simple sampling and multidirectional differentiation, ADSCs have become the ideal seed cells for bone tissue engineering (Zuk et al., 2001).
Therefore, knowing the molecular action of ADSC osteogenic differentiation provides a better understanding of the pathogenesis of bone-related diseases, such as osteoporosis, and may lead to the development of new strategies for therapies (Li et al., 2015).
It has been revealed that the biological processes of ADSCs would be controlled by various genes and epigenetic regulations (Ge et al., 2014). DNA methylation is an important epigenetic regulator, especially in the form of 5-methylcytosine (5mC), which has a critical role to play in the regulation of gene expression (Dan and Chen, 2016; Marinus and Casadesus, 2009). Of note, ten–eleven translocation (TET) protein can promote the conversion of 5mC to 5-hydroxymethylcytosine (5hmC), thereby regulating genome transcription and facilitating the normal development of mammals (Ko et al., 2013). In terms of TET2, a member of the TET family, exerts vital effects on numerous pathophysiological processes, including malignant tumors, neurodegenerative diseases, and aging (Mi et al., 2015; Zhang et al., 2014).
There was recent evidence demonstrating a close relation between TET2 and cell differentiation (Hon et al., 2014). For example, Chu et al. (2018) reported that mice with TET2 deletion led to the mild osteoporosis, which could regulate osteoclast differentiation by altering the expression of certain related genes. Besides, TET2 deletion could do harm to the self-renewal and differentiation of bone marrow mesenchymal stem cells, and eventually result in the significant reduction of bone mass, as reported in the study by Yang et al. (2018). More importantly, TET2 loss could also exert regulatory effect on the proliferation and osteogenic differentiation of bone marrow mesenchymal stromal cells (BMSCs) by negatively regulating the expression of 5mC (Li et al., 2018).
A previous finding also indicates that TET2 may play an important role during osteoblast differentiation (Silva et al., 2019). But the importance of TET2 in ADSCs has not been delineated. Therefore, we put forward a hypothesis that there may be some significant correlation between TET2 and the osteogenic differentiation of ADSCs. After isolating and culturing the human ADSCs, we investigated whether and how TET2 can affect the osteogenic differentiation of human ADSCs, with the purpose of providing an alternative perspective for the clinical treatment of osteoporosis-related bone defect.
Materials and Methods
Ethics statement
This study was approved by the Ethics Committee of Jining No. 1 People's Hospital. All animal experiments in this article were approved by the Ethics Committee of Laboratory Animals and in accordance with the Guide for the Care and Use of Laboratory Animals published by National Institute of Health (NIH) (Bayne, 1996). In addition, all tissue samples were collected from subjects after they signed the informed consent form before the study.
ADSCs isolation and culture
Adipose tissues were obtained from healthy female donators (with the age range of 25–35 years) without malignant tumors or metabolic diseases who underwent selective liposuction. After careful removal of fibrous tissues and blood vessels under sterile conditions, 10 g adipose tissue was obtained and cleaned with sterile phosphate-buffered saline (PBS) to remove contaminated debris and red blood cells. Then, the human ADSCs were isolated from adipose tissue samples and resuspended in 12 mL regular culture medium at a cell density of 2 × 106 cells/L, which was placed in a 75-cm2culture flask until cells were subcultured to the third generation for subsequent experiments.
Next, ADSCs were inoculated into a six-well plate by 2 × 105 cells/cm2, and high-glucose Dulbecco's modified Eagle's medium (H-DMEM) was added for culturing. When cell confluence reached ∼80%, the osteogenic induction medium containing DMEM, 10% (v/v) fetal bovine serum, 1% (v/v) antibiotics, 100 nM dexamethasone, 10 mM β-glycerophosphate, and 0.2 mM
Transfection of siRNA
We performed siRNA knockdown experiments of the TET2 gene. Human ADSCs were transfected with three different predesigned siRNA oligonucleotides (TET2 siRNA-1, TET2 siRNA-2, TET2 siRNA-3) targeting TET2 or a nontargeting siRNA (control siRNA). The TET2 siRNAs and the nontargeting siRNA were all purchased from Dharmacon. Before performing transfection according to the instructions on the Lipofectamine™ 2000 Kit (Invitrogen), the density of human ADSCs was adjusted to 5 × 105 cells/mL. After transfection of ADSCs lasted 24 hours, the transfection working solution was removed and then cells were washed with PBS buffer three times. The culture medium was changed into osteogenic induction medium for differentiation. Besides, the nontransfected cells were used as the Blank group.
Quantitative real-time reverse transcription PCR (qRT-PCR)
The cellular total RNA was extracted with TRIzol (Invitrogen) and the concentration of RNA was detected with a UV spectrophotometer. Then, RNA was reversely transcribed into cDNA using the PrimeScript RT Kit (RR014A; TaKaRa Biomedical Technology Co., Ltd., Beijing, China). An appropriate amount of cDNA was used as the template for polymerase chain reaction (PCR), and primers needed were designed with the software Primer5.0 (Table 1) and then synthesized by GenScript Biotech Corp. (Nanjing, China).
Sequences of Primers Used in Quantitative Real-Time Reverse Transcription PCR (qRT-PCR) in the Current Study
The reaction of qRT-PCR was performed by following the procedures on the PCR Kit (KR011A1; Tiangen Biotech Co., Ltd., Beijing, China). The expression of mRNA was calculated by using 2−ΔΔCt method and ΔCt = Cttarget gene − Ctinternal reference gene; ΔΔCt = ΔCt experiment group − ΔCt control group (Ayuk et al., 2016). The relative expression level of target genes was normalized to the expression of GAPDH.
Western blotting
Cells were washed with PBS three times, the whole protein lysate was added, and cells were split on ice for 10 minutes to obtain total proteins. Bicinchoninic acid was used for protein quantification and SDS-PAGE (sodium dodecyl sulfate–polyacrylamide gel electrophoresis) was used to transfer proteins to the nitrocellulose membrane. The proteins on the membrane were blocked for 60 minutes in 5% bovine serum albumin/tris-buffered saline with Tween-20 (TBST). Next, the membrane was incubated with primary antibodies TET1 (ab191698; Abcam), TET2 (ab124297; Abcam), and β-actin (ab8226; Abcam) at 4°C overnight.
The following day, horseradish peroxidase-labeled secondary antibodies were added for 2 hours of incubation at room temperature. Subsequently, the blots were visualized with an enhanced chemiluminescence reagent (ECL detection kit; Amersham Pharmacia Biotech). For graph analysis, ImageJ software was used to analyze the gray value of bands. β-Actin was used as a loading control to quantify the relative protein level.
Alkaline phosphatase staining and Alizarin Red staining
The Alkaline Phosphatase (ALP) Kit (Institute of Hematology, Chinese Academy of Medical Sciences) was utilized to perform ALP staining in strict compliance with the manufacturer's instructions. The activity of ALP was measured using the ALP Detection Kit (Nanjing Jiancheng Bioengineering Institute). Alizarin Red staining (all kits purchased from Nanjing Jiancheng Bioengineering Institute) was conducted as follows: The culture dish was rinsed with PBS three times, ethanol (95% v/v) was used for 10 minutes of fixation, and deionized water was used to rinse the dish three times. Then, the cells were treated with 0.1% Alizarin Red S (prepared with Tris-HCl, pH 8.3) for 30 minutes at 37°C, and positive cells were stained orange red. Distilled water was used two times to rinse and glycerol was used for sealing. A microscope was used for observation and picture taking.
Cell growth curve
The human ADSCs in each transfection group were digested and suspended in culture medium, which were seeded into a six-well plate by 1 × 105 cell/well. At the 24 hours (first day), second day, third day, and fourth day after inoculation, cells were collected and counted to draw the cell growth curve.
Immunofluorescence staining
The culture medium was removed, cells were washed with PBS buffer three times (5 min/time), and fixed in 4% paraformaldehyde for 15 minutes at 37°C. Next, cells were washed again with PBS three times, and treated for 10 minutes in precooled methanol, which was placed in a refrigerator at −20°C for pretreatment in advance.
Thereafter, cells were incubated with primary antibody 5hmC (1:500, ab214728; Abcam) overnight at 4°C, then incubated in the presence of secondary antibody for 60 minutes at room temperature. The two antibodies were diluted 1:800 for AlexaFluor 594 and 1:400 for AlexaFluor 488, respectively. Finally, cells were washed again with PBS buffer and sealed with 50 glycerol per PBS, which were prepared for subsequent observation under a Nikon fluorescence microscope. Images were acquired and analyzed with the software Image ProPlus Version 4.0 and fluorescence intensity was measured.
Experimental animals
The TET2 knockout (TET2−/−) mice were generated according to protocols reported previously (Li et al., 2011). Wild-type (WT) mice and TET2−/− mice were anesthetized and killed by cervical vertebra dislocation and long bones (femur and tibia) were taken and fixed in 75% ethanol for X-ray and microcomputed tomography (CT) scanning. The two-dimensional images were used for three-dimensional reconstruction with the software Reconstruction Utility and the ratio of bone volume to total volume (BV/TV) ratio was calculated. Next, the bilateral femur and tibia were separated and fixed in periodate/lysine/paraformaldehyde (PLP) for 24 hours, followed by 2 weeks of decalcification in EDTA, dehydration with gradient alcohol, hyalinization in xylene, embedment with paraffin, and slicing of transverse sections (5 μm in thickness).
Sections of each group were used for total collagen staining by incubating with prepared supersaturated picric acid staining solution for 1 hour, followed by 1 minute of counterstaining with Hematoxylin, 10 seconds of differentiation in 1% hydrochloric ethanol, dehydration with gradient alcohol, hyalinization in xylene, and section mounting with neutral resin. At last, sections were observed under a fluorescence microscope (Olympus, Japan) and photographs were taken with DP70 CCD (Olympus). The quantitative analysis of images was conducted by using the software Northern Eclipse (Empix Imaging). Accordingly, TUNEL staining was performed based according to the manufacturer's instructions on the TUNEL Kit.
Calcein-AM/PI double staining and Toluidine blue staining
On the first day, mice were given calcein (10 μg/g body weight) by intraperitoneal injection. On the fifth day, those mice were given the second shot of calcein. On the seventh day, mice were killed by cervical dislocation and bilateral femur and tibia were separated for 24 hours of fixation in PLP. Hard tissue embedment and super-thin section slicing were performed. Sections were observed under a fluorescence microscope (Olympus) and photographed with DP70 CCD (Olympus). The images were quantified by using the software Northern Eclipse. Quantitative parameters included mineral apposition rate (MAR), which was calculated according to the formula: MAR = The widths of two calcein deposition lines/Interval of days between two injections (Shono et al., 2010). Sections were then stained with Toluidine Blue and analyzed with the software OsteoMeasure (OsteoMetrics, Inc.).
Statistical methods
Data were analyzed by using the statistical software SPSS 21.0. Measurement data were presented by mean ± standard deviation. The comparison among multiple groups was conducted by using one-way ANOVA and the intergroup comparison (post hoc test) among multiple groups was analyzed with Turkey's test. The difference between two groups was tested by Student's t-test. p < 0.05 indicated the statistical significance of differences.
Results
TET1 and TET2 expressions during osteogenic differentiation of human ADSCs
Under the optical microscope, the human ADSCs were spindle shaped (Fig. 1A) and confirmed to be the osteoblasts after differentiation induction at day 4, as shown by the analysis results of ALP staining (Fig. 1B). Alizarin Red staining demonstrated the existence of matrix mineralization at day 15 (Fig. 1C). Meanwhile, we detected TET1 and TET2 expressions on the 0, 3rd, 5th, 7th, 10th, and 14th day of osteogenic differentiation. qRT-PCR and western blotting analysis revealed that TET1 mRNA and protein levels were relatively unchanged during ADSCs differentiation. Meanwhile, TET2 mRNA and protein levels significantly increased during the osteogenic differentiation of human ADSCs in a time-dependent manner (all p < 0.05, Fig. 1D–H).
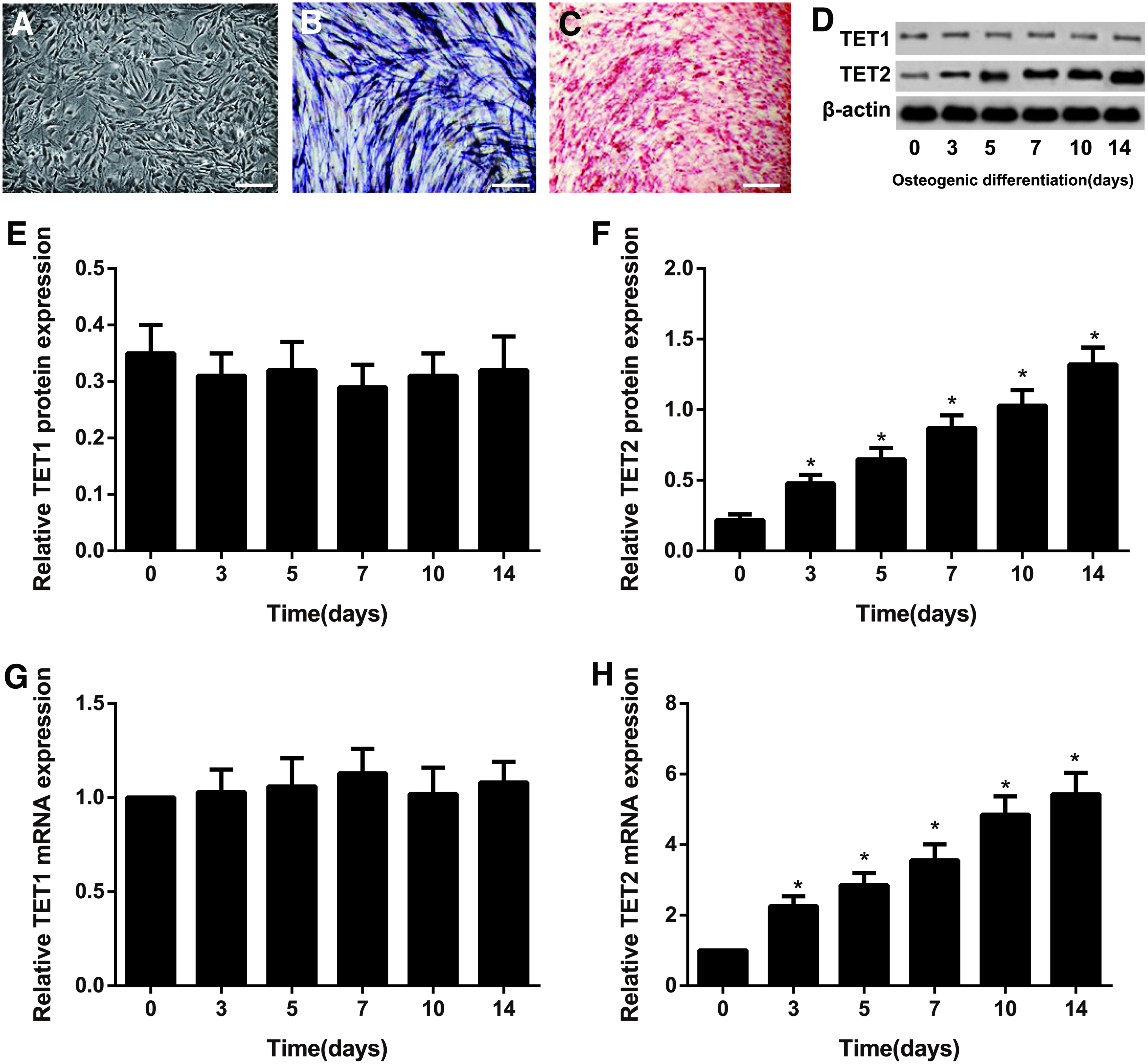
TET1 and TET2 expressions during osteogenic differentiation of human ADSCs.
Expressions of TET2 and 5hmC in the human ADSCs after transfection
As shown in Figure 2, cells in the TET2 siRNA-1, TET2 siRNA-2, and TET2 siRNA-3 groups had lower expressions of TET2 and 5hmC than Blank group (all p < 0.05), although there was no obvious difference between Control siRNA group and Blank group with respect to TET2 and 5hmC expressions (all p > 0.05). Furthermore, TET2 siRNA-3 showed the most silencing efficiency on human ADSCs among three TET2 siRNAs (p < 0.05). Then, TET2 siRNA-3 was used to transfect human ADSCs for further experiments.
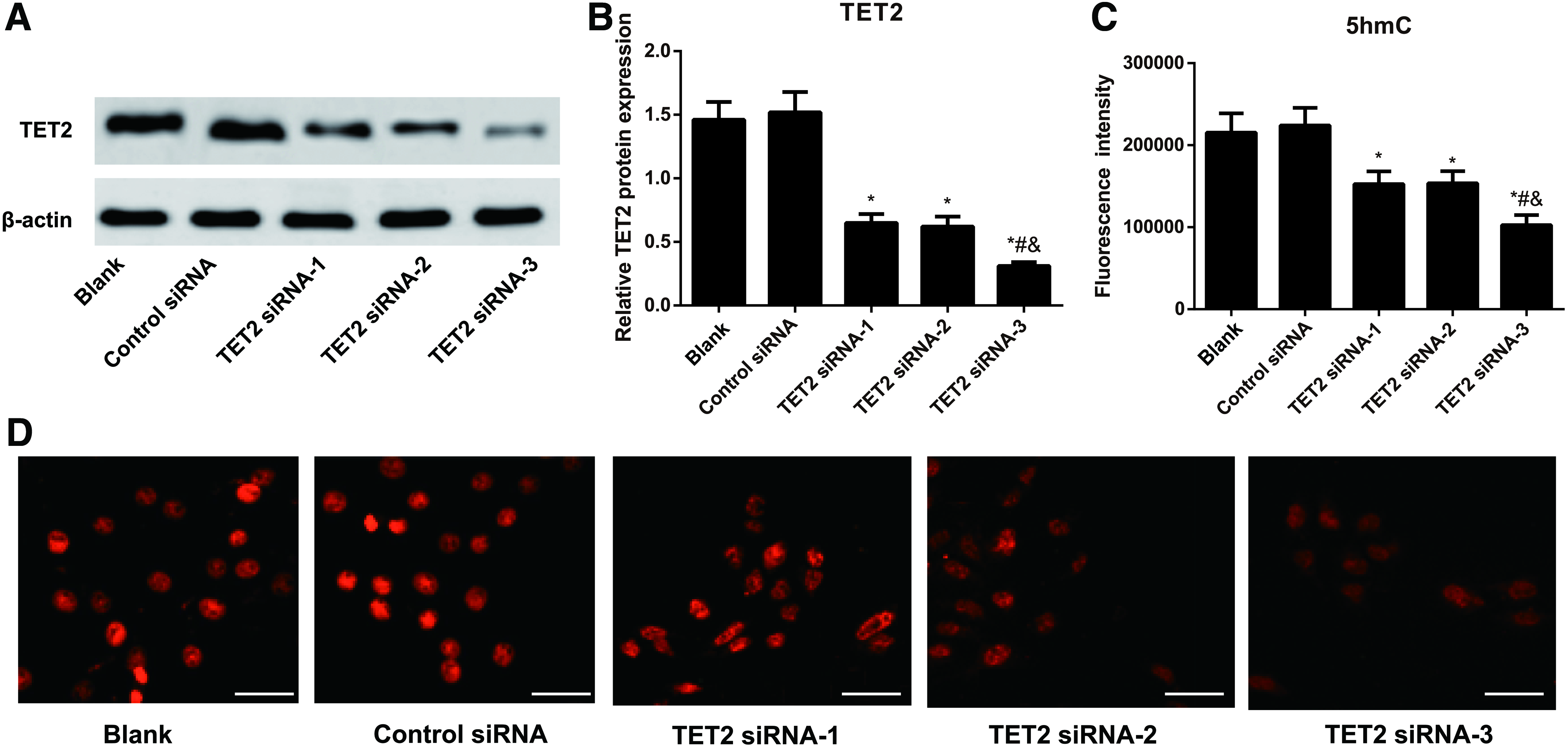
The expressions of TET2 and 5hmC in human ADSCs after TET2 siRNAs transfection.
Expressions of osteogenesis-related genes in human ADSCs after transfection
The technology of qRT-PCR was applied to detect the expression levels of osteogenesis-related genes in human ADSCs of different transfection groups. The results showed that the mRNA levels of ALP, OCN, Runx2, Col1, OSX, and OPN (osteopontin) were significantly decreased in the TET2 siRNA-3 group, but no observable differences were found regarding these osteogenesis-related genes from the Control siRNA group, when compared with the Blank group (all p > 0.05, Fig. 3).

The mRNA levels of osteogenesis-related genes (including ALP, OCN, Runx2, Col1, OSX, and OPN) in human ADSCs of each transfection group. *p < 0.05, compared with the Blank group. Results are from three independent experiments (mean ± SD). OPN, osteopontin.
Silencing TET2 inhibits osteogenic differentiation of ADSCs
According to the cell growth curve drawn based on the cell numbers, no differences were shown in the cell growth of human ADSCs from the Control siRNA group and the Blank group (all p > 0.05), whereas the cell growth declined obviously in those cells from the TET2 siRNA-3 group (all p < 0.05, Fig. 4A). As illustrated by Figure 4B–E, the human ADSCs in the TET2 siRNA-3 group exhibited light staining after ALP staining and Alizarin Red staining, with the reduced mineralized nodules and decreased ALP activity relevant to those cells in the Blank group (all p < 0.05). However, no significant difference was observed between Control siRNA group and Blank group in these aspects (all p > 0.05).
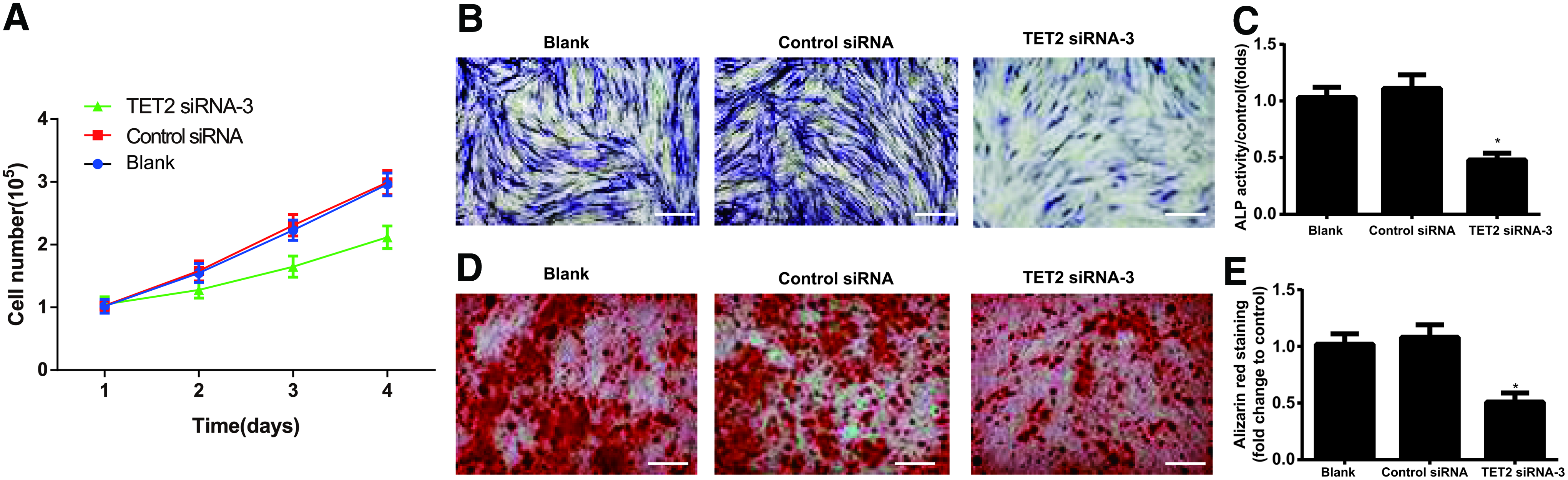
Silencing TET2 inhibits osteogenic differentiation of ADSCs.
Effects of TET2 deletion on the bone growth in vivo
The X-ray scanning and micro-CT scanning were performed to observe the changes in morphology and bone density in the left femur and tibia of WT mice and Tet2−/− mice, and the total collagen staining and TUNEL staining were conducted to detect the effect of TET2 on bone growth. As shown in Figure 5, Tet2−/− mice had shorter femur length, lower bone density, and significantly decreased BV/TV ratio than WT mice (all p < 0.05). TUNEL staining demonstrated that Tet2−/− mice had significantly higher TUNEL-positive chondrocyte percentage than WT mice (p < 0.05), which suggested that TET2 deletion induced the apoptosis of chondrocyte and inhibited intrachondral osteogenesis. Total collagen staining presented the significant reduction of total collagen-positive area in Tet2−/− mice (all p < 0.05).

Effects of TET2 deletion on the bone growth in vivo.
Effects of TET2 deletion on the osteogenesis of mouse osteoblasts
To investigate whether TET2 deletion-induced osteoporosis was associated with the reduced osteogenesis of osteoblasts, we conducted Calcein-AM/PI double staining and the results were displayed in Figure 6. Compared with WT mice, Tet2−/− mice had significantly narrowed distance between two fluorescence markers and apparently decreases in bone formation rate (BFR) and MAR (all p < 0.05). According to the results of Toluidine Blue staining of tibia, Tet2−/− mice had smaller trabecular BV and decreased osteoid formation, and also showed significant reductions in the following indexes, including BFR/bone surface (BS), BFR/BV, osteoblast number over bone perimeter (N.Oc/B.Pm), and osteoblast surface (Ob.S)/BS (all p < 0.05).
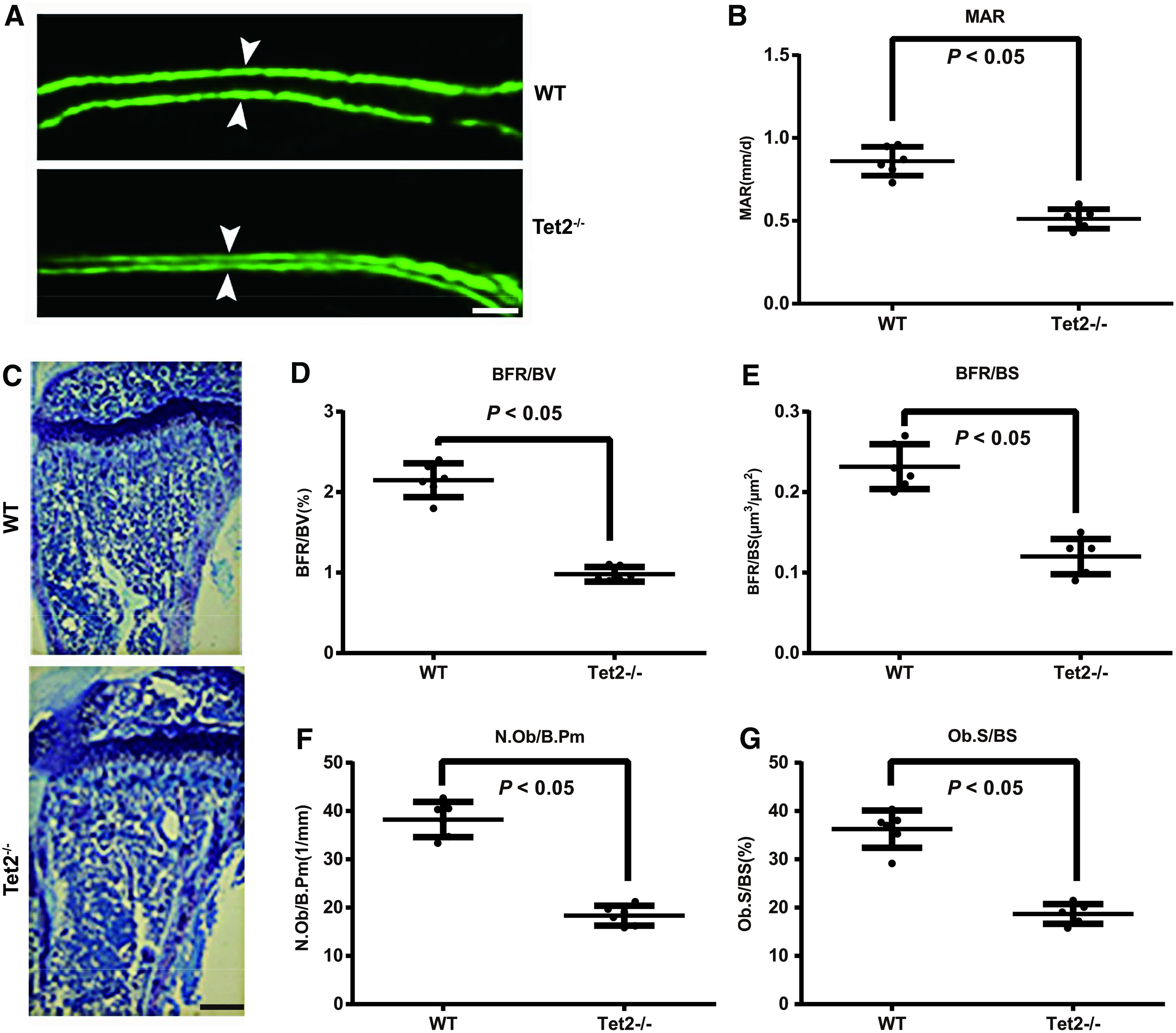
Effects of TET2 deletion on the osteogenesis of mouse osteoblasts in vivo.
Discussion
In this study, TET2 expression was found gradually increased during the process of osteogenic differentiation of human ADSCs. Zhong et al. (2017) has pointed out a similar finding as ours that TET2 expression was upregulated during myogenic differentiation, whereas TET2 silencing could reduce myogenic differentiation by downregulating the expression of myogenic genes, such as myogenin, Myf 6, and Myomaker, which indicated the involvement of TET2 in the regulation of osteogenic differentiation of human ADSCs. As far as we know, DNA demethylation is an important process affecting the differentiation of stem cells and it constitutes a mechanism of promoting osteogenic differentiation by inducing the high expression of osteogenic marker genes (Arnsdorf et al., 2010; Chen et al., 2015; Delgado-Calle et al., 2013).
5hmC was known as the intermediate leading to DNA demethylation (Feldmann et al., 2013). Evidence stated that DNA methylation-modified enhancer region has the high content of 5hmC, and TET protein family was involved in the dynamic changes of 5hmC in the enhancer region (Hon et al., 2014). As a member of DNA hydroxylase family, TET proteins have been shown to mediate DNA demethylation process by oxidizing 5mC to 5hmC, thus it has good epigenetic modification characteristics to regulate gene expression and maintain cell surface markers (Breiling and Lyko, 2015; Yang et al., 2013). Collectively, we supposed that TET2 may involve in osteogenic differentiation through regulating DNA demethylation.
Next, we conducted transfection experiments in vitro and found that TET2 siRNA could significantly reduce the expression of osteogenesis-related genes (including ALP, OCN, Runx2, Col1, OSX, and OPN). In the meantime, ALP staining and Alizarin Red staining also suggested the reduction of the human ADSC osteogenic differentiation after silencing TET2. ALP is a specific marker of a mature osteoblast phenotype and it is one of the most common indicators used for the evaluation of the secretory function of osteoblasts (Metavarayuth et al., 2015). Meanwhile, RUNX2 is a transcription factor greatly involved in the regulation of osteoblast development, osteoblast differentiation, and bone formation, and it exerts its regulatory impact by inducing the expression of osteogenic differentiation-ending proteins, including bone sialoprotein, OCN, OSX, and OPN (Lee et al., 2010).
On the other hand, as the protein framework of bone tissues, Coll is of great implication for the maintenance of biomechanical properties and structural integrity of bone tissues, and it is also a useful marker to indicate the differentiation of osteoblasts toward matrix maturation (Nyman et al., 2011). Consistent with our results, Dimitrios Cakouros and his team also reported that TET2 and 5hmC expression levels went up substantially during BMSC osteogenic differentiation, and TET2 knockout by siRNA led to the significant decrease of osteogenic potential, appreciable reductions in RUNX2, BMP-2, OPN, and OCN, and shrinking number of mineralized nodules (Cakouros et al., 2019).
These findings indicated that TET2 silencing may inhibit osteogenic differentiation of human ADSCs by blocking the transcription of osteogenic genes. Actually, TET2 mutations, including deletion, insert, and frameshift mutation, tend to be accompanied with the significant reduction of total 5hmC, as described previously (Ko et al., 2010). Thus, we detected the expression levels of 5hmC in the human ADSCs from each group, and as a result, we found TET2 siRNA can inhibit both TET2 and 5hmC expressions. During the process of osteogenic differentiation, a large number of promoter regions related to osteogenic differentiation were demethylated to facilitate the gradual acquisition of osteoblast phenotypes by osteoblasts, and these promoter regions included Runx2, OCN, OPN, ALP, and Col1 (Ling et al., 2017; Zhang et al., 2011; Zhou et al., 2009).
Therefore, silencing TET2 may inhibit 5hmC to hinder the demethylation of osteogenic genes and downregulate the expression of osteogenesis-related genes, thus eventually resulting in the inhibition of osteogenic differentiation. Moreover, Van Etten et al. (2011) revealed that TET2 was linked to activation of the Hedgehog in chronic myeloid leukemia, suggesting various possible targets for new therapies. The Hedgehog signaling was reported to be essential in osteoblast generation from preosteoblasts (Marumoto et al., 2017).
In addition, Qu et al. (2018) have shown that TET2 deficiency led to the activation of AKT and ERK and resulted in stem cell factor-dependent clonal expansion of dysfunctional erythroid progenitors. Baroncelli et al. (2019) found that inhibition of PI3K signaling significantly reduced osteoblast viability and adhesion to ECM by decreasing the AKT activity. Considering the above studies, TET2 might also regulate osteogenic differentiation of ADSCs through Hedgehog or AKT pathways. But the potential molecular mechanisms still need to be investigated in future studies.
Furthermore, the animal experiments in vivo were also performed and we found mice with TET2 deletion showed significant decreases in bone density, BV/TV, and collagen area, but apparent increases in chondrocyte percentage, suggesting that TET2 deletion may cause premature osteoporosis. In line with our observation, the study of Chu et al. (2018) also demonstrated the mild osteoporosis of mice with the reduced 5hmC and the alterations of osteogenesis-related genes induced by TET2 loss.
Moreover, the Tet2−/− mice presented osteoporosis phenotype in the work of Yang et al. (2018) and TET2 deletion could reduce the demethylation of P2rX7 promoters to inhibit the expression of osteogenesis-related genes. Additionally, Tet2−/− mice in our investigation also showed significant decreases in MAR, BFR/BS, BFR/BV, N.Oc/B.Pm and Ob.S/BS, all of which were indicators to reflect the details of osteoblast-related activity from different aspects (Allen et al., 2006; Ciria-Recasens et al., 2005; Matsushima et al., 2003; Smith et al., 2005), and further verified the findings consistent with our in vitro experiments.
In conclusion, TET2 expression was up-regulated during the osteogenic differentiation of the human ADSCs. However, TET2 deletion could down-regulate the expressions of osteogenesis-related genes to hinder the osteogenic differentiation. Hence, this study may offer some experimental and theoretical basis in human ADSCs for finding a potential therapeutic strategy for patients with osteoporosis.
Footnotes
Acknowledgment
The authors thank all members for their helpful suggestions and comments on this work.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
The authors received no funding for this study.