
Abstract
To evaluate age-related changes in T cells and stem cell-related genes in mice and the changes in T cells and stem cell-related genes after allogenic bone marrow mesenchymal stem cell (BMSC) transplantation, BALB/c mice were divided into young (2 months, n = 5) and old (20 months, n = 5) groups and 1 × 107 BMSCs from 3-week-old C57BL/6J mice were injected into the old mice (n = 5). T lymphocytes including CD3+, CD8+, CD8+CD28+, and CD8+CD44lowCD62LhighSca-1+ stem cell-like memory T cells from spleens were analyzed by flow cytometry. mRNA transcriptions of the tumor suppressor p16INK4A and the senescence inhibiting AUF 1 and stem cell-related ADAM12, GIL3, c-MYC, NANOG, Wnt, HOX11, Sox2, Oct3/4, and KLF4 genes were analyzed by quantitative reverse transcription–polymerase chain reaction for comparison between young and old mice and old mice after BMSC application. Stem cell-related genes were reduced transcribed in old mice, an action that could be partly or completely reversed for some genes by BMSC injections. The proportion of CD8+CD28+ T cells in the spleens of old mice was significantly reduced (p < 0.01), indicating advanced proliferative T cell senescence. The CD8+CD44lowCD62Lhigh cell fraction was significantly reduced and that of CD8+CD44lowCD62LhighSca-1+ increased in splenic CD8+ cells of old mice, both actions of which were reversed by BMSC injections. p16INK4A transcription was enhanced and AUF1 transcription was reduced in old mice, the latter effect partly reversed by BMSC injections. BMSC injections led to recovery of stem cell-related gene activation or BMSC stem cell-related gene expression tolerance in spleens of old mice.
Introduction
Aging has been defined as a decline of intrinsic physiological functions, resulting in an increase of age-specific mortality rates (Flatt, 2012). Cellular senescence has been attributed to oncogene-induced senescence (Chandeck and Mooi, 2010), DNA damage-induced senescence (Minieri et al., 2015), and replicative senescence (Cristofalo et al., 2004). Since Hayflick (1965) postulated a limited number of cell division for diploid cells, the question arose whether cell division limitations also exist for immune system cells and it has been noted that replicative senescence is reached after 25–40 divisions for both CD4/helper and CD8/cytotoxic T cell subpopulations (Effros, 2004). In line with T cell division restriction is the finding that even in younger people with autoimmune diseases or chronic viral infections, high levels of senescent T cells have been detected by their lost CD28 expression (Vallejo et al., 2004).
However, naive T cells have stem cell characteristics and after their transformation into memory T cells, some subsets were proposed to maintain these stem cell-like properties (Stemberger et al., 2009). In a previous study, Zhang et al. (2005) reported the discovery of candidate memory stem cells in a mouse graft-versus-host disease model, which consisted of a postmitotic CD44low CD62Lhigh CD8 (+) subset of T cells. These cells were able to self renew in addition to generate and sustain central memory, effector memory, and effector CD8 (+) T cells (Zhang et al., 2005). Later studies revealed that a CD44low CD62Lhigh CD8 (+) subset of T cells with high stem cells antigen-1 (Sca-1), a marker for hematopoietic stem cells, CD122 and antiapoptotic protein B cell lymphoma 2 (Bcl-2) expressions could be generated by Wnt signaling (Gattinoni et al., 2009).
However, reports about CD44low CD62Lhigh CD8 (+) on T cells are sparse. In this study we have investigated age-related changes in T cells focusing on CD44low CD62Lhigh CD8 (+) T cells and related gene expression in a mouse aging model. We hypothesized that bone marrow mesenchymal stem cell (BMSC) application to old mice might have a rejuvenation effect on their CD44low CD62Lhigh CD8 (+) T cell population.
Materials and Methods
Animals
Three-week-old female C57BL/6J mice were purchased from the Institute of Zoology, Chinese Academy of Military Medical Sciences (Permission no. SCXK [Jing] 2012-0001). Female BALB/c mice were divided into two age groups with five animals in each group (2 and 20 months). The 2-month-old mice were purchased from the Institute of Zoology of the China Academy of Military Medical Sciences (License no. SCXK [Beijing] 2012-0001). The 20-month-old mice were purchased from the Sichuan Dashuo Laboratory Animal Company (License no. SCXK [Sichuan] 2008-24) at the age of 17 months, and fed in the special pathogen-free animal laboratory belonging to the Institute of Zoology of the China Academy of Military Medical Sciences (the center meets the national standards for laboratory animal facilities [GB14925-2001]).
All procedures involving animals were performed in accordance with the ethical standards of the participating institution and the guidelines for the Humane Treatment of Laboratory Animals (Ministry of Science and Technology of the People's Republic of China, Policy no. 2006 398). The mice were anesthetized by intraperitoneal injection of 0.2% pentobarbital sodium at a dose of 40 mg/kg. With mice in the supine position, the skin was cut in the middle of the abdomen, and the spleen was found in the left upper quadrant and completely resected.
Isolation of splenocytes
First, 5 mL of Dulbecco's modified Eagle medium (DMEM) containing 2% fetal bovine serum (FBS) was added to a 50 mL centrifuge tube. Then, a 200-mesh sieve was put on the top of a 50 mL tube and the spleen was grinded on the sieve using the plunger of a 10 mL glass syringe, followed by centrifugation at 1200 rpm/min for 5 minutes. After centrifugation, the supernatant was discarded and 3 mL red blood cell lysis buffer 1 × working solution was added and mixed with a pipette tip for ∼3 minutes after which 10 mL DMEM containing 2% FBS was added for neutralization. After centrifugation at 1200 rpm/min for 5 minutes the cell number was counted and adjusted to 2 × 107/mL. The cell suspension was divided in 100 μL aliquots and the respective antibodies were added according to the recommended instructions.
After shaking, the suspension was incubated at 4°C in the dark for 30 minutes and 1 mL phosphate-buffered saline (PBS) was added followed by centrifugation at 1000 rpm/min for 5 minutes, after which the supernatant was discarded and the cells were suspended in 200 μL PBS for flow cytometry analysis.
Flow cytometry analysis
The antibodies used for cytometry APC/Cy7-conjugated mAbs to CD3ɛ (145-2C11), PE/Cy7-conjugated mAbs to CD4 (GK1.5), V450-conjugated mAbs to CD8a (53-6.7), APC/Cy7-conjugated mAbs to CD8a (53-6.7), PerCP/Cy5.5-conjugated mAbs to CD44 (IM7), and APC-conjugated mAbs to CD62L (MEL/14) were purchased from BD Biosciences (San Jose, CA) and fluorescein isothiocyanate (FITC)-conjugated mAbs to CD28 (E18), Alexa Fluor 647-conjugated mAbs CD11B (M1/70), Alexa Fluor 488-conjugated mAbs CD29 (TS2/16), PE-conjugated mAbs CD31 (390), PE/Cy5-conjugated mAbs CD34 (HM34), PE/Cy7-conjugated mAbs CD45 (30-F11), FITC-conjugated mAbs CD86 (GL-1), PE/Cy7 conjugated mAbs to Sca-1 (D7), and Alexa Fluor 488-conjugated mAbs to Sca-1 (D7) were purchased from BioLegend (San Diego, CA).
Cells were measured with a Gallios flow cytometer (Beckman Coulter, Krefeld, Germany) and analyzed using Kaluza (Beckman Coulter) software.
RNA isolation and cDNA synthesis of spleen tissue
Spleen tissue sample of 50–100 mg was added to 1 mL of TRIzol solution (Thermo Fischer Scientific) and homogenized at 50 Hz for 3–4 minutes. The sample volume did not exceed 10% of the TRIzol volume. After homogenizing twice, the isolation of RNA was performed according to the instruction of the manufacturer and the integrity of RNA was detected by Nanodrop. Subsequently, cDNA was synthesized using 0.5 mM Oligo(dT)12–18 Primer and 200 U M-MLV RT (Invitrogen). Reverse transcription was performed according to the instruction of Invitrogen. All cDNA samples were diluted with DNAase/RNAase-free water and stored at −80°C.
Primers were designed for p16INK4A, AUF1, ADAM12, GIL3, NANOG, c-MYC, HOX11, Wnt, Sox2, Oct3/4, and KLF4 by Primer 3.0 software and synthesized by the Shanghai Biological Engineering Co., Ltd. The sequences of forward and reverse primers used are given in Supplementary Table S1. The melting curves of the genes showed only one peak, indicating good primer amplification specificity. Components of the real-time polymerase chain reaction (PCR) system were as follows: 0.6 μL forward primers, 0.6 μL reverse primers, 10 μL SYBR-green, 6.8 μL ultrapure water, and 2 μL cDNA. Specific reaction procedures were as follows: predenaturation at 95°C for 5 minutes, denaturation at 95°C for 10 seconds, and renaturation at 60°C for 30 seconds (45 cycles) and 72°C for 10 seconds.
The instrument used was a Lightcycler 480 real-time PCR instrument (Roche Molecular Diagnostics) with GAPDH as control. The expression level of each target gene was standardized using the 2−(ΔΔCt) method.
Isolation, culture, identification, and amplification of mouse BMSCs
When we designed the study, we chose BALB/c as recipients and C57BL/6J as donor mice, because it mimics allotransplantations, which might be more suitable for a clinical study. BMSCs were collected from 3-week-old female C57BL/6J mice. The animals were killed by cervical dislocation and sterilized with 75% alcohol for 5 minutes. Femur and tibia were taken out sterile in a super clean bench, placed in the specimen collection solution (2 mL FBS, 2 mL penicillin [100 U/mL], 2 mL streptomycin [100 μg/mL] in 94 mL PBS), and cooled on ice. Then bones were placed in 10 mL culture dishes with bone marrow cavity rinse solution (2 mL FBS in 98 mL PBS).
After cutting through the femur and tibia, each bone was rinsed for 10 times until the bone and marrow cavity turned white. A 1 mL syringe was used to collect the rinse solution. Then bones were transferred into 3.5 cm Petri dishes containing 2 mL of collagenase (0.25% type I) and placed in an incubator at 37°C for softening. After ∼5 minutes, the bones were cut into pieces with a scissor in super clean bench and transferred into 5 mL circular bottom flow tubes with collagenase. The flow tube was fastened covered with sealing film, and was then transferred to a 37°C thermostat shaker at 200 rpm for 45 minutes.
Then, the digested bone pieces and collagenase were transferred into a 50 mL centrifuge tube containing 25 mL of bone marrow cavity rinse solution. After allowing to be settled for 3 minutes, the supernatant was filtered through a 70 μm filter into a new 50 mL centrifuge tube. Then 10 mL of bone marrow cavity rinse solution was added to the settled bone pieces and thoroughly mixed. After 3-minute settling time, the supernatant was filtered through a 70 μm filter and combined with the former filtered fluid.
After centrifugation at 1200 rpm for 10 minutes, bone marrow cells were resuspended in 5 mL of BMSC complete medium (100 mL BMSC medium supplement; stem cell catalog no. 05502). Then the suspension was added into 400 mL of BMSC basic medium (catalog no. 05501; STEMCELL Technologies, Beijing) and 3% methylene blue acetate was added to dilute for counting. Cells were then inoculated into six-well plates at 7 × 105 cells per well containing 2 mL of medium per well. After 10-day culture, cells reached 80% confluence and were harvested.
After centrifugation and discarding the supernatant, cells were resuspended in complete medium and subcultured at a 2:1 ratio, marked as P1. In the following cell passages the ratio was improved to 1:2 or 1:3 and P5 led to a cell culture, which exhibited high expressions of CD29 and Sca-1 and low expressions of CD119, CD31, CD34, CD45, and CD86. In addition, P5 aliquots differentiated into osteogenic (C57BL/6J mice BMSC osteogenic induction differentiation medium, catalog no. MUBMX-90021; Cyagen Biosciences, Guangzhou, China) and adipogenic (C57BL/6J mice BMSC adipogenic induction differentiation medium, catalog no. MUCMX-90031; Cyagen Biosciences) cells in their respective selective media (Supplementary Fig. S1) (Zhu et al., 2010).
P6 cells were selected for injection of 1 × 107 cells after resuspension into 200 μL Dulbecco's phosphate-buffered saline into old BALB/c mice’ caudal veins with a 1 mL syringe. The mice were killed 1 month after the treatments. An aliquot of CM-Dil-stained BMSCs was used to follow their distribution in mice spleens after injection (Supplementary Fig. S2).
Statistical analysis
Statistical analyses were performed using SPSS for Windows (version 16.0; SPSS, Inc., Chicago, IL). Data are expressed as mean ± standard deviation and comparisons among different age groups were made using one-way analysis of variance, with p < 0.05 considered significant.
Results
Changes of different T cell proportions in the spleens related to aging and after the administration of BMSC to old mice
From the results of flow cytometry analysis of T cells in different age mice, we found that the proportion of CD8+CD28+ cells significantly decreased in the spleens of old mice (13.1% vs. 6%; Fig. 1A).

Comparison of the proportion of different T cells in young and old mice and in old mice 1 month after caudal vein BMSC injection.
The number of CD3+ T cells in total splenocytes was 47.8% in young and 36.3% in the elderly group. The proportion of CD4+ T cells in total splenocytes was 24.2% in young and 21.0% in aged mice, whereas the proportion of CD8+ T cells in total splenocytes was 17.8% in young and 9.8% in the elderly group (Fig. 1B). The proportion of CD8+CD44lowCD62Lhigh T cells in total splenocytes was 5.1% in young and 3.7% in the elderly group, whereas the proportion of CD8+CD44low CD62Lhigh Sca-1+ T cells in total splenocytes was 1.4% in young and 3.2% in old mice (Fig. 1C). The proportions of CD8+CD44lowCD62Lhigh T cells and CD8+CD44low CD62Lhigh Sca-1+ T cells in CD8+ T cells were 24.5% and 14.8% in young and 16.5% and 26.8% in the elderly group, respectively (Fig. 1D).
BMSC injections reduced the proportions of CD4+, CD8+CD44lowCD62Lhigh and CD8+CD44lowCD62Lhigh Sca-1+ T cells in the total splenocytes compared with noninjected old mice, but enhanced the proportion of CD8+CD44lowCD62Lhigh T cells in the total CD8+ cell population. In contrast, the percentage of CD8+CD44lowCD62LhighSca-1 expressing T cells in CD8+ cells was reduced after BMSC injections in old mice.
Changes of mRNA transcription of genes related to aging and stem cell properties
Spleen tissue real-time PCR measurement showed that 1 month after caudal BMSC application AUF1, ADAM12, GIL3, c-MYC, Wnt, HOX11, Sox2, Oct3/4, and KLF4 transcription was decreased and that of p16INK4A increased with mouse age. BMSC injections had no effect on p16INK4A, ADAM12, c-MYC, HOX11, and KLF4 transcriptions, whereas age-related transcriptional reductions of AUF1 were complete and that of GIL3, Wnt, Sox2, and Oct3/4 were partly reversed by BMSC administration. NANOG was essentially activated by the administration of BMSC to a greater extent in old compared with young mice.
Discussion
T cell senescence is regulated by a number of different mechanisms. After initial exposure to an antigen, telomerase activity is enhanced in T cells and gradually declines to undetectable levels for ∼3 weeks. However, from the fifth exposure to antigens, telomerase activity is completely absent in CD8+ T cells, which is paralleled with a loss of CD28 expression, the first T cell-specific change indicating replicative senescence (Valenzuela and Effros, 2002). In our study, the percentage of CD8+CD28+ T cells in the spleen was significantly reduced in old mice, indicating that during aging a high proportion of their T cells became senescent (Fig. 1). However, a small subpopulation of memory CD8+CD44lowCD62LhighSca-1+ T cells might escape the fate of reproductive senescence because they are self-maintaining owing to their remaining stem cell properties.
The percentages of CD8+CD44lowCD62Lhigh T cells of all CD8+ cells were 25% in young mice and dropped to 15% in old mice, indicating a loss or shift into different subgroups of these cells, which could be reversed by BMSC injection into the old mice to levels of young mice. In contrast the percentages of CD8+CD44lowCD62LhighSca-1+ T cells of all CD8+ cells were 14% in young mice and increased to 25% in old mice, indicating that during the lifespan of mice the memory T stem cell reservoir is increased in older mice, which requires Wnt signaling to induce Sca-1 expression (Gattinoni et al., 2009).
As given in Figure 2, Wnt expression in young mice was high, but fell to undetectable levels in old mice, which might have led to a constant increase of CD8+CD44lowCD62LhighSca-1+ T cells over time with a fixed final pool in older age mice with inactivated Wnt expression. However, BMCS injections led to a reduction of the CD8+CD44lowCD62LhighSca-1+ T cell pool in old mice and Wnt signaling was partly recovered, thereby restoring conditions similar to young mice. p16INK4A is a tumor suppressor gene, which arrest cells in the G1 phase and is believed to be a marker of aging (Krishnamurthy et al., 2004).
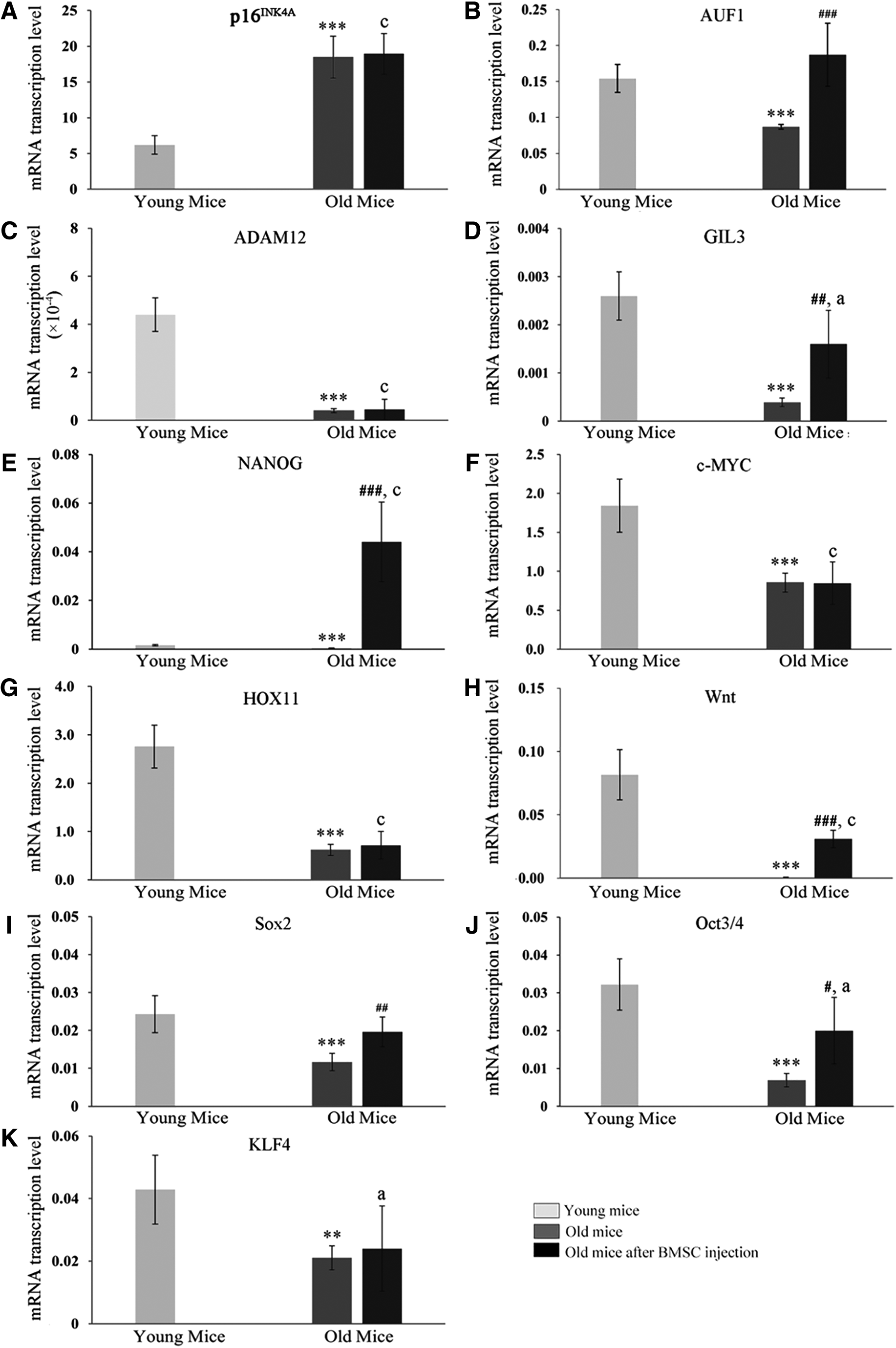
Indicated gene transcription rates in spleens of young, old mice and old mice 1 month after caudal vein BMSC injection. Transcription levels of
p16INK4A expression was enhanced in aged mice and was not influenced by the administration of BMSC, but in contrast AUF1, which suppresses senescence by activating telomerase transcription (Pont et al., 2012) was downregulated in old mice, but restored by BMSC injections, indicating that BMSC changed the splenic aging pattern. Apart from T cell activation (Langeveld et al., 2006), the spleen has been recognized as a stem cell-producing organ with activity of induced pluripotent stem cell-specific genes Oct3/4, Sox2, KLF4, c-MYC, and NANOG and embryonic stem cell-specific genes HOX11, GIL3, Wnt, and ADAM12. The splenic stem cells can differentiate into T cells (Dieguez-Acuna et al., 2010).
With the exception of NANOG, all stem cell-related genes are downregulated in the spleens of old mice, indicating reduced stem cell productivity during an advanced lifespan. However, BMSC leads to upregulation of NANOG and recovery of splenic GIL3, Wnt, Sox2, Oct3/4, and KLF4 activities in old mice spleens. These data indicated that BMSC can trigger stem cell-related gene activity in the spleen of old mice or that BMSC could maintain stem cell-related gene expression in the spleens of old mice.
In summary, although the proportion of CD8+CD28-T cells in the spleens of old mice was significantly reduced, indicating advanced proliferative T cell senescence, the stem cell-like CD8+CD44lowCD62LhighSca-1+ memory T cells accumulated to a higher level in old compared with young mice spleens. All stem cell-related gene transcriptions were reduced in old mice, which could be partly or completely reversed for some genes provided by BMSC injections, which indicates that BMSC injections led to recovery of stem cell-related gene activation or BMSC stem cell-related gene expression tolerance and concomitant maintenance in spleens of old mice.
Footnotes
Authors' Contributions
B.Z., Z.Z., and X.Z. were responsible for the conception and design of the study. B.Z., Z.Z., X.Z., W.D., and Y.L. were responsible for acquisition and analysis of data; B.Z., Z.Z., X.Z., and W.D. were in charge of statistical analysis. B.Z. drafted the article; B.Z., and Z.Z. revised and commented the draft, and all authors read and approved the final version of the article.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
No funding was received for this work.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.