
Abstract
Since no study has explored whether exercise could improve impaired proliferation, migration, and angiogenesis of endothelial progenitor cells (EPCs) in animal models or humans with type 2 diabetes, we aimed to explore the effect of different models of exercise on EPC function and expression of caveolin-1, PI3K, and AKT in mice with type 2 diabetes. Male db/db mice (age: 8 weeks) with type 2 diabetes were subjected to aerobic training (AT), resistance training (RT), or combined aerobic and resistance training (AT+RT) 3 or 4 days/week. Mice in the control group remained sedentary with no specific training requirement. Bone marrow-derived EPCs were isolated, and the protein concentrations of caveolin-1, Pi3k, and AKT, and EPC function, were identified in the 1st, 4th, 8th, and 12th weeks of the intervention. Greater increases in proliferation, migration, and angiogenesis were observed in the AT, RT, and AT+RT groups than in the control group. AT+RT was more effective than AT or RT in improving the migratory and angiogenesis function of EPCs in mice with type 2 diabetes and achieved maximum improvement after 8 weeks of intervention. Western blot analysis showed that caveolin-1, p-PI3k, and p-Akt levels were obviously increased in the AT, RT, and AT+RT groups compared with the control group. The expression level of these proteins in the AT+RT group was higher than that in the AT and RT groups. AT+RT may be a helpful reference when choosing exercise methods for the prevention of diabetes-related cardiovascular diseases.
Introduction
High blood glucose induces decreases in endothelial progenitor cell (EPC) proliferation, migration, and angiogenesis, resulting in impaired EPC ability to regenerate new vessels and accelerate atherosclerosis, which may contribute to a higher risk of cardiovascular complications in patients with type 2 diabetes (Rigato et al., 2015; Schmidt-Lucke et al., 2005).
Although aerobic exercise was reported to improve the proliferation, migration, and angiogenesis of EPCs in healthy individuals and patients with metabolic syndrome, congestive heart failure, coronary heart disease, and prediabetes (Hoellriegel et al., 2011; Ribeiro et al., 2013; Sandri et al., 2016; Sen et al., 2014; Sonnenschein et al., 2011; Tsai et al., 2016; Volaklis et al., 2013), to the best of our knowledge, no study has explored whether exercise could improve impaired EPC proliferation, migration, and angiogenesis in animal models or humans with type 2 diabetes.
Caveolin-1 was found to be highly abundant in endothelial cells (ECs) and EPCs. A previous study documented that knockdown of caveolin-1 contributed to impaired EC proliferation, migration, and angiogenesis (Feng et al., 2012). Improvement of EPC function was detected when the PI3K/AKT pathway was active (Cheng et al., 2010).
In conclusion, it is well established that chronic exercise training has the ability to enhance EPC function. For patients with type 2 diabetes, this is of clinical importance since EPCs have been implicated in vascular repair and revascularization. However, studies are needed to identify the best mode of exercise training that will upregulate circulating EPCs as well as clarify the mechanism by which exercise improves EPCs.
We aimed to explore the influence of exercise models on EPC function and protein expression of caveolin-1, PI3K, and AKT in mice with type 2 diabetes and provide a helpful reference for choosing exercise methods for the EPC treatment of diabetes-related cardiovascular diseases.
Materials and Methods
Animals and experimental design
Ethical approval was granted by the Guangxi Medical University of Institutional Animal Care and Use Committee. Eight-week-old male type 2 diabetic db/db (BKS-Dock Leprem2Cd479/Nju) littermates (Nanjing Biomedical Research Institute of Nanjing University) were randomly divided into the control group, aerobic training (AT) group, resistance training (RT) group, and combined aerobic and resistance training (AT+RT) group (n = 20/each group). Five mice from each group (20 mice in total) were killed at each assessment point (1st, 4th, 8th, and 12th weeks) for measurement of EPC function and protein concentration.
During the intervention, a tail blood sample was obtained for glucose measurement with a glucose meter (Johnson & Johnson, Milpitas, CA) before starting the experiment and 24 hours after training, which was repeated every week. Type 2 diabetes was diagnosed when the blood glucose level of a mouse was above 11.1 mmol/L twice. EPCs were isolated in the 1st, 4th, 8th, and 12th weeks of the intervention. Five mice (selected by a random number table) in the four groups were intraperitoneally euthanized with a dose of 100 mg/kg sodium pentobarbital in accordance with the U.K. Animals (Scientific Procedures) Act, 1986, and the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 8023, revised 1978).
Exercise training
Mice were fed ordinary chow and maintained under a 12-hour light/12-hour dark cycle. Mice in the control group were kept sedentary with no specific training requirement, and the exercise groups performed AT, RT, or AT+RT. AT was performed on a treadmill (WDW-1; Beijing North Rui Future Analytical Instrument Co., Ltd., Beijing, China) specifically designed for mice.
Mice in the AT group were initially trained at a speed of 10 m/min for 10 minutes 3 days/week in the first week. Beginning the second week, the exercising mice performed AT for 50 minutes at a speed of 13–17 m/min 3 days in the second week and 4 days for the next 10 weeks. The mice in the AT group were trained for a total of 12 weeks (the mice ran 3–4 days a week) (Lin et al., 2015).
The mice in the RT group climbed a 1-m ladder composed of a 2-cm grid inclined at 85°. The mice in the RT group climbed from the bottom of the ladder to the top while carrying the appropriate weights attached to their tails. The weight was 50% of the mouse's body weight in the first week and gradually increased throughout the training period: 1st and 2nd week, 50%; 3rd and 4th week, 60%; 5th and 6th week, 80%; 7th through 12th week, 100%. All mouse body weights were measured every day. The mice were motivated to finish three sets of five repetitions of climbing 3 days/week in the first 2 weeks and 4 days/week over the next 10 weeks. The mice were allowed to rest for 1 minute between repetitions and 2 minutes between the sets (Lee and Farrar, 2003).
The mice in the AT+RT group performed AT following the protocol of the AT group and RT following the protocol of the RT group.
Isolation and culture of bone marrow-derived EPCs
The medullar channels of the tibiae and femurs of each mouse were bluntly dissected and flushed with 10 mL of phosphate-buffered saline (PBS). Cell suspensions were collected by filtration and added to the lymphocyte-separating solution at a ratio of 1:1. Cell suspensions were centrifuged at 400 g for 30 minutes. The precipitates were subsequently resuspended in EC basal medium-2 containing 10% fetal bovine serum (Gibco) and vascular endothelial growth factor (VEGF; R&D Systems).
Definition of EPCs
Characterization of EPCs
Cells were observed under an inverted microscope. Spindle-shaped cells observed under an inverted microscope were defined as bone marrow-derived EPCs.
Cellular staining
Cells were first incubated with acetylated low-density lipoprotein (Molecular Probes) (10 μg/mL) at 37°C for 4–6 hours and then fixed with 4% paraformaldehyde for 10 minutes. Subsequently, the washed cells were incubated with Ulex europaeus agglutinin-1 (Sigma Chemical Co., St. Louis) (10 mg/mL) for 1.5 hours. The number of EPCs was determined according to the number of cells in five randomly selected high-power fields (200 × ) via an inverted fluorescence microscope (Olympus Corporation).
Immunofluorescence staining analysis
Cells were immunofluorescently stained overnight at 4°C with CD31 (Proteintech Group), CD34 (Beijing Bioss Molecular Co., Ltd.), CD133 (Proteintech Group), CD144 (Beijing Bioss Molecular Co., Ltd.), and VEGFR2 (Proteintech Group) antibodies diluted to 100 mL. The cells were incubated with CoraLite488-conjugated secondary antibody (Proteintech Group) and Alexa Fluor 594-conjugated secondary antibody (Proteintech Group) in the dark at room temperature for 2 hours. The cellular DNA was stained with DAPI (Beijing Solarbio Science & Technology Co., Ltd.), and cells were examined by fluorescence microscopy (Olympus Corporation).
Proliferation assay
Suspended EPCs were plated on a collagen-coated 96-well plate (3.6–4.0 × 103 cells/well) and cultured for 24 hours. Subsequently, suspended EPCs were incubated for another 4 hours under lucifugal conditions with 10 μL of CCK-8 solution (Dojindo Molecular Technologies, Inc.) in each well. Then, the plate was agitated for 10 seconds, and the optical density was measured at a wavelength of 450-nm with a microplate reader.
Migration assays
A straight line was drawn with a marker on the back of a 6-well plate; next, 1–3 × 106 EPCs were seeded per well on the plate and cultured for 24 hours. The 6-well plate was washed three times with 1 × PBS buffer. The cells were added to serum-free medium and maintained in a 37°C, 5% CO2 incubator (Thermo Fisher Scientific, Inc.). Images were obtained using an inverted microscope (magnification 100 × ) at 0 and 48 hours and were analyzed with ImageJ (National Institutes of Health).
In vitro angiogenesis assay
A total of 1–2 × 105 EPCs were seeded on a plate coated with Matrigel matrix (R&D Systems) and incubated for 18 hours in a 37°C, 5% CO2 incubator. Then, images of tube morphology were obtained with an inverted microscope. A capillary tube was defined as a tubular structure with a length that was four times the width. The mean number of capillary tubes was determined, and the tubes were photographed via an inverted microscope (magnification 100 × ) and analyzed by ImageJ.
Western blot
Primary antibodies against the following proteins were used for Western blotting: caveolin-1 (Cell Signaling Technology), total PI3K p85 (t-p-PI3Kp85; Abcam), phosphor-PI3K p85 (p-PI3Kp85; Abcam), total AKT (t-Akt; Cell Signaling Technology), and phosphor-Akt (p-Akt; Cell Signaling Technology). Membranes were washed with Tris-buffered saline tween (TBST) buffer (Beijing Leagene Biotechnology Co., Ltd), and they were probed with HRP-conjugated secondary antibodies (goat anti-rabbit or goat anti-mouse IgG). Protein bands were visualized, and densitometric analyses were performed using enhanced chemiluminescence (LI-COR Biosciences) to quantify protein expression concentrations.
Statistical analysis
The Shapiro–Wilk test was performed to check the normality of the continous data. Equality of variance was tested by Levene's test. If the residual variables of each group and subgroup were homogeneous and the equality of variance was satisfied, a two-way analysis of variance (ANOVA) was used to detect the difference in the concentrations of proteins (caveolin-1 and Pi3k/Akt) and the function of EPCs (proliferation, migration, and angiogenesis in vitro) among the four interventions and time points in mice with type 2 diabetes. The Bonferroni test was used for adjustment of multiple comparisons. All the data were analyzed with SPSS software 22.0 (SPSS, Inc., Chicago, IL), and p < 0.05 (two-tailed test) was considered indicative of a significant difference.
Results
The effect of exercise on EPC proliferation
As shown in Figure 1D, the proliferative activities of EPCs were significantly lower in the control group than in the AT group, RT group, and AT+RT group. No difference in proliferative activity was found between the AT, RT, and AT+RT groups.
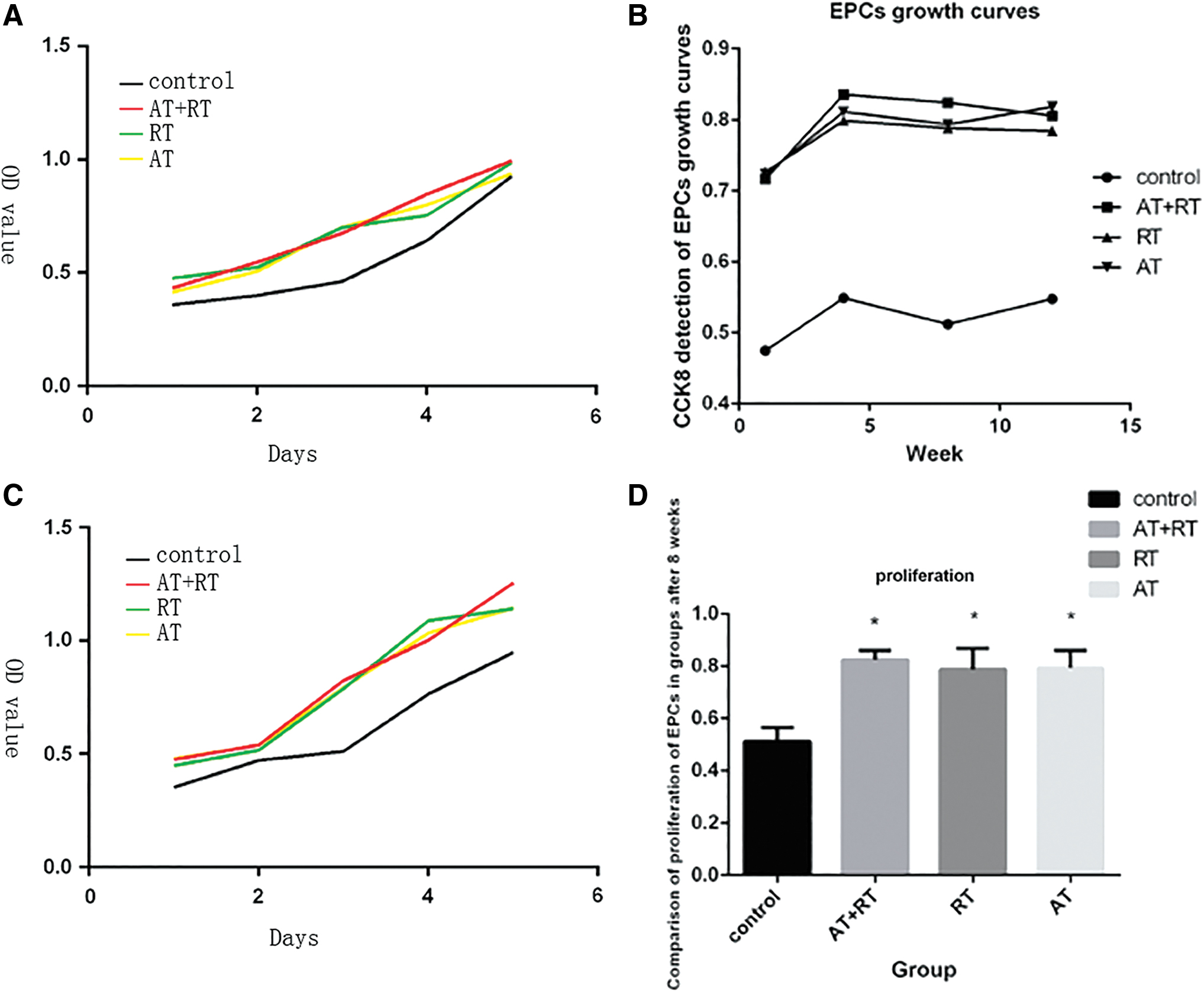
Proliferative function of EPC
The effect of exercise on EPC migration
As shown in Figure 2B, the EPCs in both the AT and AT+RT groups achieved greater migratory ability in the 8th week than in the 1st, 4th, and 12th week. The RT group showed a significant increase in EPC migration in the 8th week compared with that in the 1st and 12th week. As shown in Figure 2C, compared with the control group, the AT, RT, and AT+RT groups exhibited significant increases in EPC migratory ability, and an increase in migration was detected in the AT+RT group compared with the AT or RT group at 8 weeks of intervention.

Migratory function of EPC.
The effect of exercise on EPC angiogenesis in vitro
As shown in Figure 3B, the EPCs in both the AT and AT+RT groups achieved greater angiogenic function in the 8th week than in the 1st, 4th, and 12th week. EPC angiogenesis in the RT group was significantly increased in the 8th week compared with that in the 1st and 12th week. As shown in Figure 3C, compared with the control group, the AT, RT, and AT+RT groups exhibited significant increases in angiogenic ability. Higher angiogenesis was presented in the AT+RT group than that in the AT or RT group at 12 weeks of intervention.

Angiogenic function of EPC in vitro.
The concentrations of caveolin-1 and Pi3k/Akt determined by Western blot analysis
As shown in Figure 4, there were significant differences in the concentrations of caveolin-1, p-PI3Kp85, and p-AKT between the control group and the AT, RT, and AT+RT groups at 4 weeks of intervention. AT+RT was more effective than AT, RT, and the control regimen in increasing the concentration of caveolin-1 and Pi3k/Akt at 8 weeks of intervention.
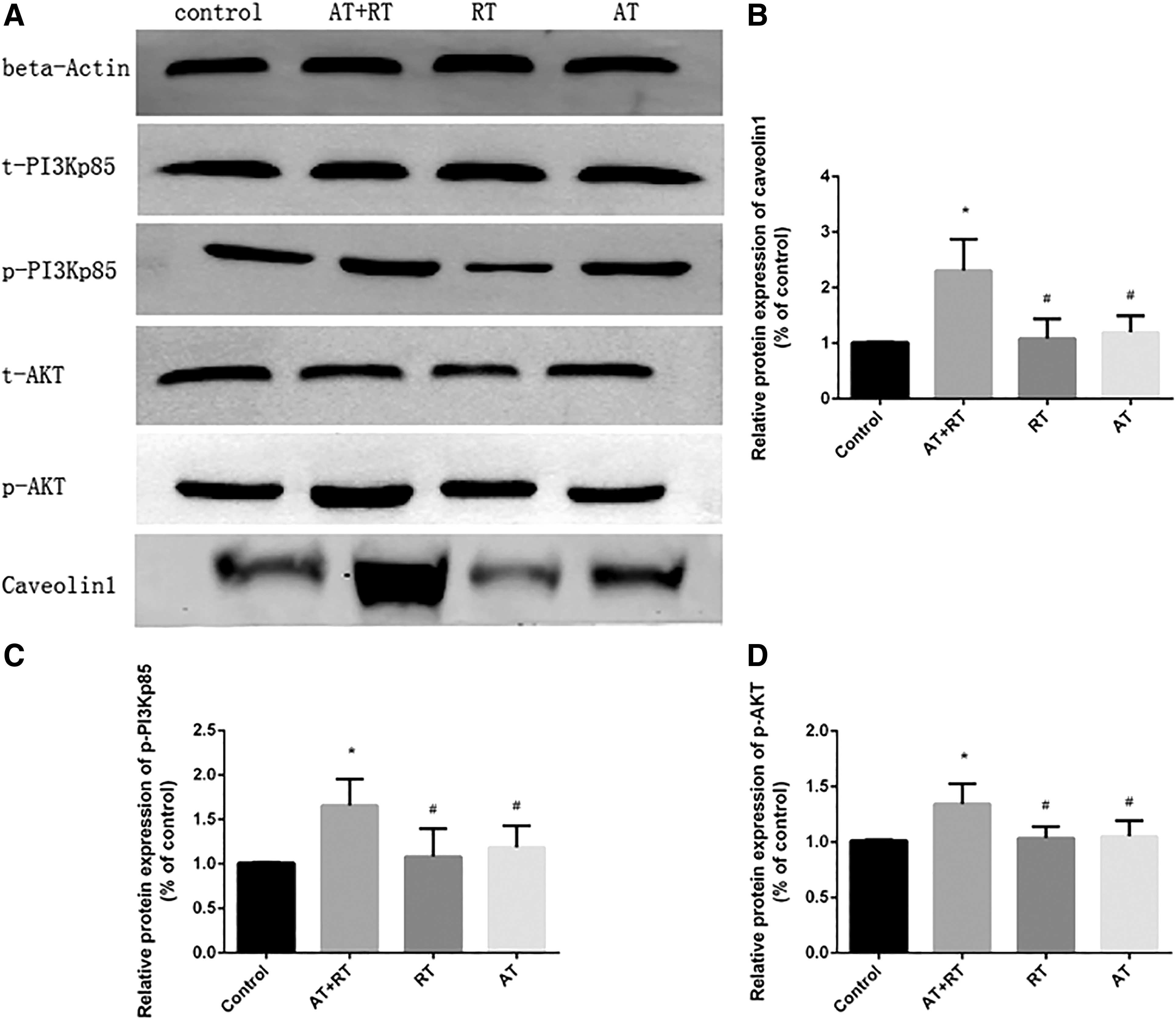
The results from caveolin-1 and Pi3k/Akt proteins in all groups at the 8th week of intervention.
Changes in glucose between the groups with different interventions
After 12 weeks of intervention, the blood glucose levels of mice in the AT, RT, and AT+RT groups were significantly lower than those in the control group (see Fig. 5 for more details).
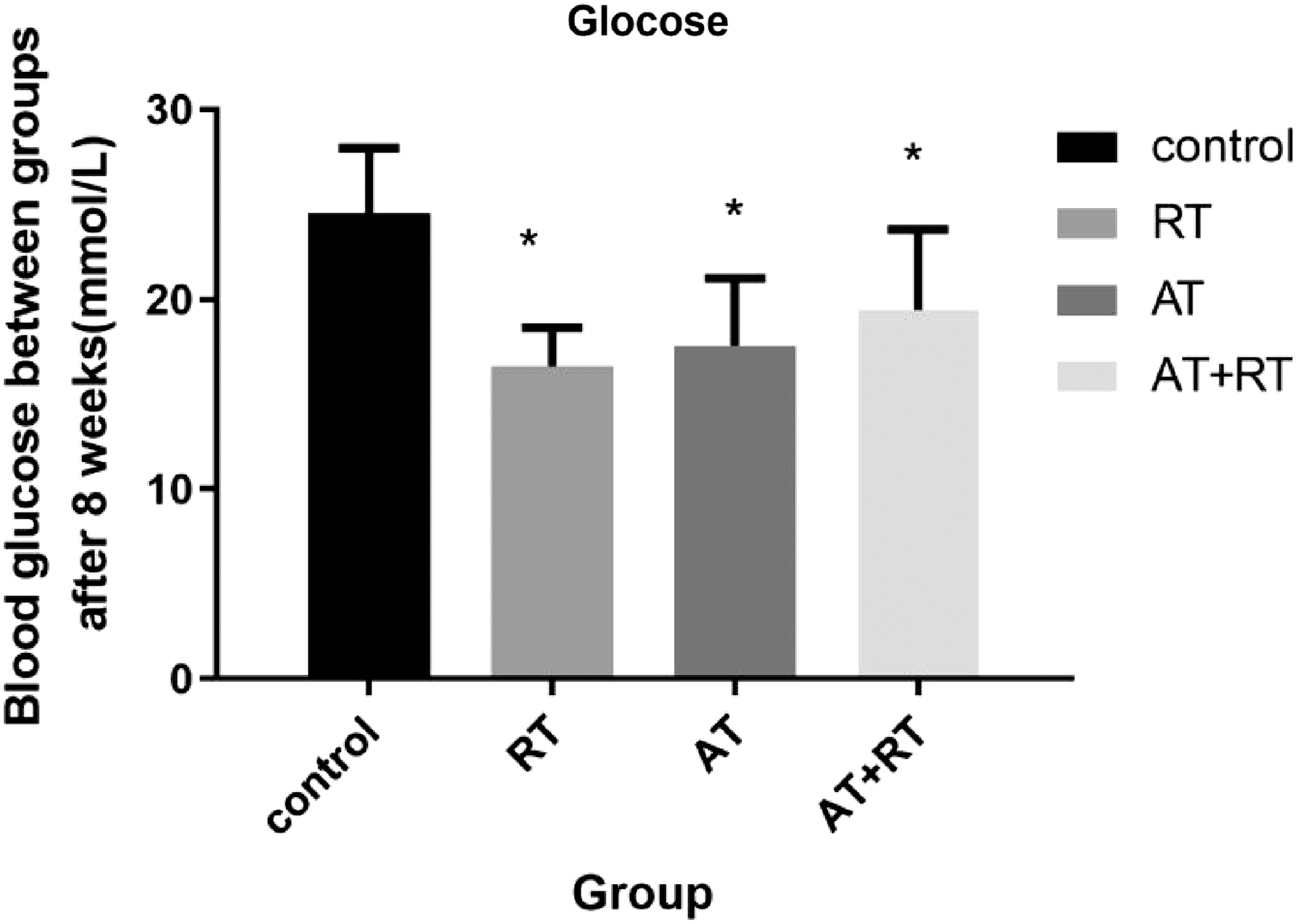
Blood glucose levels of mice following the exercise intervention. Control represents the control group, RT represents the RT group, AT represents the AT group, and AT+RT represents the AT+RT group. *p < 0.05 versus the control group.
Discussion
This study revealed that (1) RT, AT, and a combination of both can significantly improve the proliferative, migratory, and angiogenic functions of EPCs in mice with type 2 diabetes and that AT+RT induced the greatest improvement in angiogenesis of EPCs; (2) the effect of exercise on the functions of EPCs may be related to the increase in caveolin-1 and PI3KAKT; and (3) exercise attenuated the age-related decline in EPC function.
The most innovative finding of our study is that we first demonstrated the effect of RT, AT, and a combination of both interventions on the proliferative, migratory, and angiogenic functions of EPCs in mice with type 2 diabetes. The most important result obtained in this study is that AT+RT is more effective than either isolated AT or isolated RT in enhancing the proliferation, migration, and angiogenesis of EPCs in mice with type 2 diabetes. Exercise has been shown to enhance the proliferative, migratory, and angiogenic abilities of EPCs (Hoellriegel et al., 2011; Sandri et al., 2016; Sen et al., 2014; Sonnenschein et al., 2011; Tsai et al., 2016), but the effect differs based on the type of exercise (De Biase et al., 2014; Ribeiro et al., 2017; Waclawovsky et al., 2015).
AT has been well studied and shown to enhance the function of EPCs in patients with hypertension, obesity, metabolic syndrome, coronary artery disease, and chronic heart failure (Hoellriegel et al., 2011; Sandri et al., 2016; Sen et al., 2014; Sonnenschein et al., 2011; Tsai et al., 2016). After a single bout of RT, a significant increase in some important factors of EPCs was identified, including the angiogenic factors stromal cell-derived factor 1α (SDF-1α), VEGF, hypoxia-inducible factor 1-α, SDF-1α, and matrix metalloproteinase-9 (De Biase et al., 2014; Ribeiro et al., 2017; Waclawovsky et al., 2015), which were reported to be able to enhance the proliferative, migratory, and angiogenic abilities of EPCs (Chang et al., 2015; Cheng et al., 2010; De Biase et al., 2014; Huang et al., 2009). This study confirmed the benefit of AT+RT in the proliferation, migration, and angiogenesis of EPCs, forming the basis of research on therapy for some diseases related to EPCs.
The results of two-way ANOVA in our study showed that EPC migration and angiogenesis were lower in the AT, RT, and AT+RT groups after 12 weeks of intervention than after 1, 4, and 8 weeks of intervention, which was unexpected. We speculated that with progression and at the highest intensity of the training, 12 weeks of exercise should have stimulated a higher increase in migration and angiogenesis. Moreover, a similar decrease in EPC migration and angiogenesis existed in the control group after 12 weeks of intervention and then after 1, 4, and 8 weeks of intervention. Therefore, we hypothesized that age may have a negative impact on EPC function in mice.
A study provided support for our results by reporting that the function of EPCs starts to decline when humans reach middle age since all diabetic mice, with a maximum life span of 40 weeks, were 20 weeks old (equivalent to middle age in humans) at the end of the 12-week intervention in the study (Rauscher et al., 2003).
Our study showed higher proliferative, migratory, and angiogenic activities of EPCs in the AT, RT, and AT+RT groups than in the control group, although an age-related decline was observed in all groups after 12 weeks of intervention. This finding indicated that AT, RT, and AT+RT were able to attenuate the decrease in EPC function caused by aging. Studies documented that age-related decline in EPC function could be attenuated effectively by exercise in animal and human models with or without chronic heart failure (Cheng et al., 2010; Sandri et al., 2016; Xia et al., 2012; Yang et al., 2013). These studies supported our study.
The most significant result of this study was that the upregulation of caveolin-1, PI3K, and AKT is involved in the mechanism by which AT, RT, and AT+RT effectively enhanced EPC function in mice with type 2 diabetes. Previous studies have documented that an inhibited PI3K/AKT signal reduced EPC capabilities to proliferate, migrate, and generate new blood vessels (Ankeny et al., 2012) and eliminated improvement on EPC functions in mice of advanced age by exercise (Cheng et al., 2010). Knockdown caveolin-1 decreased PI3K, AKT, and EPC proliferative, migrative, and angiogenic functions in mice with multiple organ dysfunction syndrome (Luo et al., 2019). Therefore, previous studies reported that caveolin-1/PI3K/AKT plays an important role in EPC proliferative, migrative, and angiogenic functions.
Another study showed that AT increased PI3K/AKT expression and improved EPC proliferation, migration, and angiogenesis in mice with myocardial infarction (Guo et al., 2016), which was consistent with our study. A study showed that AT promoted caveolin-1 expression and enhanced neural stem cells (Zhao et al., 2017). Based on the aforementioned studies, we deduced that AT, RT, and a combination of both trigger increases in the expression of caveolin-1 and PI3K/AKT, resulting in the promotion of EPC proliferative, migratory, and angiogenic capacities in mice with type 2 diabetes.
Studies (Cheng et al., 2010; Luo et al., 2019; Obi et al., 2012) proved that the expressions of caveolin-1, Pi3k, and Akt were closely related to improvement of EPC functions. The expressions of caveolin-1, Pi3k, and Akt, as well as EPC migration and angiogenesis, were lower in the AT, RT, and AT+RT groups at 12 weeks of intervention than that at 1, 4, and 8 weeks of intervention, which may be a result of advanced age mice in our study after 12 weeks of intervention. A study gave support to our study by reporting that the function of EPCs starts to decline when humans reach middle age (Rauscher et al., 2003).
The limitations of this study included in vitro observations that could not reflect the impact on the entire body and a lack of baseline variables regarding EPC function. However, a control group that did not participate in a specific exercise intervention was used to identify the effects of exercise and remaining sedentary on EPC function in this randomized-controlled study. Our study is limited to observations of the relationship between exercise and caveolin-1 and PI3K/AKT expression. Future studies are needed to elucidate the mechanism.
Conclusion
In conclusion, the results of this study showed that AT+RT can be expected to improve EPC function in mice with type 2 diabetes by increasing the protein concentrations of caveolin-1 and Pi3k/Akt, which may be a helpful reference when choosing exercise methods for the prevention of diabetes-related cardiovascular diseases. The effect of exercise on EPC function is suggested to be closely related to the upregulation of caveolin-1, PI3K, and AKT.
Footnotes
Authors' Contributions
All authors made contributions to this study by conceptualization (X.D., Y.P.Y.), methodology (X.D., L.Z., Y.P.Y., Q.S., B.B.L., C.W.), investigation (X.D., L.Z., Y.H.L., Y.F.H., C.M.), formal analysis and data curation (X.D., L.Z.), writing—original draft preparation (X.D., L.Z.), and writing—review and editing (X.D., Y.P.Y.). All authors provided a critical review of the article and read and approved the final version.
Acknowledgment
This study was carried out at the First Affiliated Hospital of Guangxi Medical University, Guangxi, China.
Ethics Approval
The present study was approved by the Ethics Committee of the First Affiliated Hospital of Guangxi Medical University (Guangxi, China).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
Funding Information
This work was supported by the National Natural and Scientific Foundation (81660147).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.