
Abstract
The genetic diversity of Neotropical deer is increasingly jeopardized, owing to declining population size. Thus, the formation of cryobanking of somatic cells is important for the preservation of these species using cloning. The transformation of these cells into viable embryos has been hampered by a lack of endangered species oocytes. Accordingly, the aim of this study was to produce brown brocket deer embryos by interspecific somatic cell nuclear transfer (iSCNT), using goat or cattle oocytes as cytoplasts, and to elucidate embryo mitochondrial activity by measuring the expression levels of ATP6, COX3, and ND5. Cattle embryos produced by in vitro fertilization (IVF) were used as a control. There were no differences in the development of embryos produced by traditional SCNT and iSCNT when using either the goat cytoplasts (38.4% vs. 25.0% cleaved and 40.0% vs. 50.0% morula rates, respectively) or cattle cytoplast (72.8% vs. 65.5% cleaved and 11.3% vs. 5.9% blastocyst rates, respectively). Concerning the gene expression, no significant difference was observed when goat oocytes were used as cytoplasts. However, when using cattle oocytes and 16S as a reference gene, the iSCNT upregulated COX3, when compared with SCNT group. In contrast, when GAPDH was used as a reference gene, all the evaluated genes were upregulated in the iSCNT group, when compared with the IVF group. When compared with the SCNT group, only the expression of ATP6 was statistically different. In conclusion, it was demonstrated that interspecific nuclear transfer is a potentially useful tool for conservation programs of endangered similar deer species.
Introduction
According to the IUCN (International Union for Conservation of Nature) (2016), of 10 known species of brown brocket deer (Mazama spp.), 6 are categorized as vulnerable, and 2 are categorized as least concern (LC), whereas insufficient data are available to assess the remaining two. Brocket deer are widely used as a food resource for populations in Central and South America and are likely to suffer from strong predation pressures (Taber et al., 2016). Furthermore, the description of new species in the genus is expected, owing to the observation of chromosome polymorphisms that function as efficient mechanisms of postzygotic reproductive isolation (Abril et al., 2010; Duarte et al., 2008; Salviano et al., 2017), and many of the new species will be categorized as endangered (Duarte and Jorge, 2003).
The brown brocket deer (Mazama gouazoubira) exhibits impressive ecological plasticity, occurring in all the biomes of Brazil, except for the Amazon basin (Black-Décima et al., 2010). The species is classified, by the IUCN (2016), as a species that does not suffer immediate threats to survival (LC) and has been used as a model deer species for many procedures (Zanetti et al., 2010).
Historically, threatened species have been preserved through the protection of their habitat. However, this strategy is currently insufficient, owing to the high rate of human population growth. Therefore, assisted reproductive technologies (ARTs), such as artificial insemination and in vitro fertilization (IVF), followed by embryo transfer, have been widely applied to preserve species and have yielded encouraging results (Locatelli et al., 2006; Oliveira et al., 2016). Among the different ARTs, somatic cell nuclear transfer (SCNT) is worth highlighting and has been significantly improved (Hansen, 2014) since the birth of Dolly (Wilmut et al., 1997). Somatic cells, such as skin biopsies, are easy to store and can be used to produce viable cell lines for nuclear donation (Magalhães et al., 2017).
Cryogenic banks of such cells already exist for some endangered Neotropical deer species. For example, the Deer Research and Conservation Center (NUPECE) of São Paulo State University (UNESP) have established a cell bank of >1000 individuals of 10 Neotropical deer species. Such resources can function as a repository of genetic diversity for small populations. However, to produce viable embryos from these cells, oocytes (cytoplast) will be needed to receive the fibroblast (karyoplast) nuclei, and the current captive populations of Neotropical deer are small and unavailable for invasive procedures, such as laparoscopy (Cursino et al., 2014).
Considering the difficulty of obtaining oocytes from threatened animals, interspecific SCNT (iSCNT) is especially useful once the embryo is reconstructed using karyoplasts from the endangered species and a cytoplast from farm animals (Lagutina et al., 2013). Indeed, several studies have demonstrated the utility iSCNT in endangered species (Lanza et al., 2000; Loi et al., 2001).
Despite these achievements, the success rate of iSCNT remains low, and even though many factors contribute to the method's inefficiency, it seems that failures in nucleus–mitochondria communications (St John et al., 2004) and the basis of nuclear transfer itself (Niemann et al., 2008) play the greatest roles. In addition, because mitochondria perform a broad range of critical functions related to cellular energy supply and programmed cell death, it is possible that inadequate or perturbed mitochondrial functions may adversely affect the success of interspecific cloning (Hiendleder and Wolf, 2003).
Even though the brown brocket deer is not an endangered species, the iSCNT procedure can be optimized in this species and then used as a model for performing nuclear transfer in other deer species. Accordingly, the aim of this study was to evaluate the in vitro development of brown brocket deer cloned embryos produced by iSCNT using cattle and goat cytoplasts and to assess embryo mitochondrial activity (MA) by measuring the expression levels of some mitochondrial genes (ATP6, COX3, and ND5).
Materials and Methods
Chemicals and ethics
All chemicals were purchased from Sigma Chemicals Company (St. Louis, MO, USA) unless otherwise indicated, and the study was approved by the Ethics Committee on Animal Use of Ceará State University (6552020/2016) and the Chico Mendes Institute for Biodiversity (45727-1).
Oocyte collection and in vitro maturation
Cattle and goat ovaries were obtained from a local abattoir and transported to the laboratory in warm isotonic saline (32°C–37°C) within 2 hours. Cumulus-oocyte complexes from follicles of >2 mm in diameter were aspirated using an 18-gauge needle (Hasler, 1998). The medium used for in vitro maturation (IVM) was TCM199 with 10 ng/mL epidermal growth factor, 100 μM cysteamine, 0.2 mM pyruvate, 1 μg/mL estradiol, 100 U penicillin, 100 μg/mL streptomycin, 0.25 μg/mL amphotericin, 20 μg/mL FSH/LH (Pluset; Calier, Barcelona, Spain), and 10% estrus goat serum when using goat oocytes, or 10% fetal bovine serum (FBS; Thermo Fisher Sci., Waltham, MA, USA) for cattle oocytes. The IVM conditions were 5% CO2 in humidified air at 38.5°C for 20–21 hours (Cavalcanti et al., 2018).
Establishment and culture of donor fibroblasts
Cell culture was prepared from skin biopsies of experimental animals (deer, cattle, and goat). The samples were cultured in Dulbecco's Modified Eagle's Medium (DMEM) (Thermo Fisher Sci.) supplemented with 10% FBS, 100 U penicillin, 100 g/mL streptomycin, and 0.25 g/mL amphotericin at 38.5°C and 5% CO2 in air. After 7–14 days of incubation, monolayers with fibroblast-like morphology were disaggregated with Trypsin-EDTA (Thermo Fisher Sci.), then regrown and passaged 3–7 times, before being resuspended in DMEM with 10% FBS and 10% dimethyl sulfoxide, and, finally, stored in liquid nitrogen (Magalhães et al., 2017). The quiescence of donor cells was induced by growth to confluence for 5–6 days before the scheduled experiment. At 30 minutes before SCNT or iSCNT, the cells were prepared by trypsinization (Trypsin-EDTA, 0.25%), washed, and then resuspended in DMEM.
Cytoplast preparation
After IVM, cattle and goat oocytes were denuded of cumulus cells by repeatedly pipetting with hyaluronidase solution (1 mg/mL TCM199) for 2 minutes and then washing the cells in TCM199 for 1 minute. The resulting denuded oocytes were stained with Hoechst 33342 (5 μg/mL) for 10 minutes, and evaluated for maturation under inverted microscope (TE2000; Nikon, Tokyo, Japan). Enucleation was performed by aspirating the first polar body and ∼10% of the underlying cytoplasm using a beveled enucleation micropipette (15–20 μm internal diameter) and a holding pipette (40 μm internal diameter, 120 μm external diameter) to support the oocyte during enucleation under inverted microscope equipped with a micromanipulator (NT-88-V3; Narishige, Tokyo, Japan). Enucleation was confirmed by observing the stained metaphase plate inside the pipette under UV light.
Nuclear transfer
Embryo reconstruction was achieved using fibroblast/cytoplast, respectively, for the following groups: deer/goat, goat/goat, deer/cattle, and cattle/cattle. The nuclear transfer and oocyte activation procedures were those described by Ross and Cibelli (2010). Fibroblast with a smooth plasma membrane was injected into perivitelline space of enucleated oocyte using an injection pipette. The couplets were placed in fusion medium (0.3 M mannitol, 0.1 mM MgSO4, 0.05 mM CaCl2, and 1 mg/mL polyvinyl alcohol) for 30 seconds and then transferred to a 0.2-mm microfusion chamber (Micro chamber; Eppendorf, Hamburg, Germany) that contained 1 mL fusion medium. Membrane fusion was performed using an electroporation system (Multiporator, Eppendorf) with two 25 μseconds DC pulses of 2.0 kV/cm, and the couplets were placed in synthetic oviduct fluid (SOF) medium.
The activation of reconstructed embryos was induced by exposure to 5 mM ionomycin in TALP-H for 4 minutes and immediate incubation in 1.9 mM 6-dimethylaminopurine in SOF medium for 3 hours.
Cattle IVF
A group of IVF-derived embryos was designed only for cattle to serve as a control. In brief, proven fertility semen was thawed and separated using a 4-mL Percoll gradient (55%/90%) and centrifugation at 700 g for 25 minutes. The Percoll-separated semen was washed twice with 2 mL of Brackett–Oliphant (BO) medium that was supplemented with 5 mM caffeine and 20 IU/mL heparin (Calbiochem; Merck, Darmstadt, Germany). After washing, the semen was diluted with BO added with 10 mg/mL bovine serum albumin (BSA) to achieve a final concentration of 20 × 106 spermatozoa/mL.
Finally, the IVM oocytes were washed in manipulation medium that included TCM-199 supplemented with 1% (v:v) antibiotic–antimycotic solution, 0.1 mM sodium pyruvate, 10% (v:v) FBS, and 25 mM HEPES, and then coincubated with the semen suspension in 100-μL droplets covered with mineral oil under a humidified 5% CO2 atmosphere at 38.5°C for 6 hours. After incubation, presumptive zygotes were denuded by vortexing in manipulation medium.
Culture of in vitro fertilized and reconstructed embryos
Presumptive zygotes were cultured in vitro in four-well cell culture plates, containing 500 μL SOF supplemented with 2.5% FBS and 0.4% BSA, under mineral oil. The culture conditions were a humidified gas mixture of 5% CO2, 5% O2, and 90% N2 at 38.5°C (Cavalcanti et al., 2018). Cleavage and morula/blastocyst rates were assessed on day 3 and 7 of culture, respectively. The morula and blastocyst rates were determined on the number of cleaved embryos.
Gene expression analyses
Total RNA extraction
Media was removed from the cultured embryos by washing with phosphate-buffered saline, and the embryo samples were frozen at −80°C. Total RNA was extracted using the RNeasy micro kit (Qiagen, Inc., Hilden, Germany) according to the manufacturer's instructions. In brief, each frozen sample was resuspended in 75 μL RLT Plus buffer and mixed with an equal volume of 70% ethanol. The mixture was then transferred to an RNeasy MinElute spin column from the RNeasy Micro kit for RNA to bind to the column. RNA purification included an RNAse-free DNase treatment for 15 minutes at room temperature. After three washes, the RNA was eluted with 12 μL RNAse-free water. Fibroblasts known to express each of the target genes were also used as positive control.
Reverse transcription
For each experimental group, a total RNA corresponding to three blastocysts (IVF group, cattle/cattle, and deer/cattle) and two or three morulae (goat/goat and deer/goat) were used. The procedure was performed by adding 1 μL Random Hexamers (Promega, Madison, WI, USA) to each sample, followed by heating to 70°C for 5 minutes and then cooled for 2 minutes. After, it was added 4 μL Improm II Reaction Buffer (Promega); 1 μL Improm II Reverse Transcriptase (Promega); 0.5 mM dNTPs (Promega); 2 U/μL Recombinant RNasin (Promega); 3mM MgCl2 (Promega) and RNase-free water for a final reaction volume of 20 μL.
Reverse transcription was performed at 42°C for 60 minutes and then at 70°C for 15 minutes. The cDNA products were then stored at −80°C for later use as templates for real-time polymerase chain reaction (PCR). Negative controls or RT blanks were prepared under the same conditions but without the inclusion of reverse transcriptase.
Real-time quantitative PCR
The expression levels of mitochondrial genes that encoded electron transporters (COX3, ND5) and a subunit of the ATP synthase enzyme (ATP6) were analyzed. Specific primers pairs for amplifying each gene (Table 1) were designed using Primer-BLAST (Ye et al., 2012) and by multiple sequence alignment of deer, goat, and cattle sequences that were constructed using ClustalW2 (Larkin et al., 2007). Target gene expressions were normalized for nuclear (GAPDH) or mitochondrial (16S) housekeeping genes (Fig. 1). MA was assessed using 16S normalization, since 16S expression is inherent to each organelle.
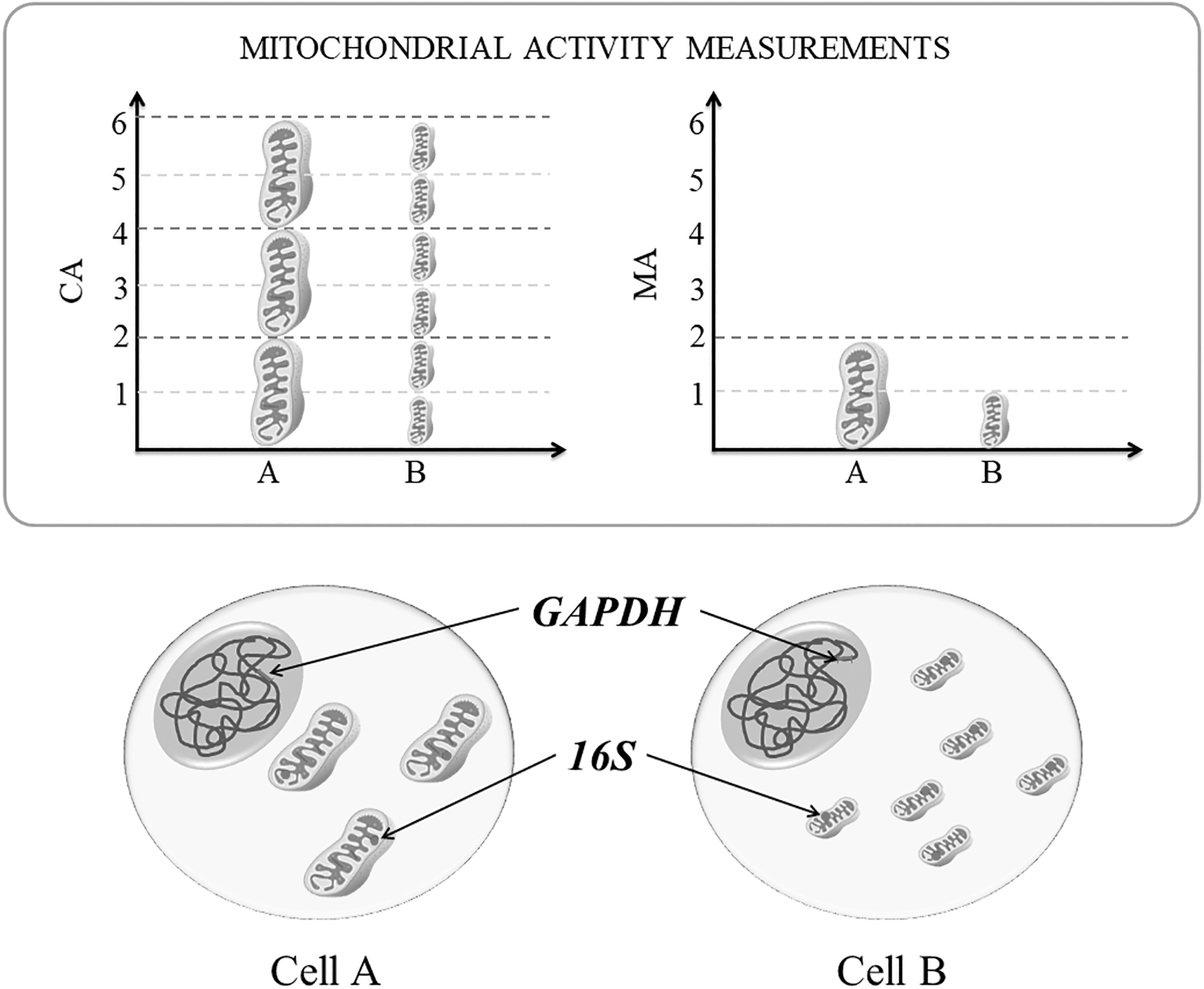
Schematic representation of measurements and interpretations of MA patterns in embryo cells. The size of each mitochondria represents the amount of MA of the organelle relative to other. The mitochondria in Cell A have twice MA than in Cell B (bottom right graphic). In contrast, the CA depends on the total amount of mitochondrial organelles and is measured by the sum of all MA (bottom left graphic). Thus, the Cell A presents CA similar to the Cell B. To access MA and CA by real-time qPCR, the signal of mitochondrial genes (ATP6, COX3, and ND5) was normalized for mitochondrial 16S or nuclear GAPDH housekeeping genes, respectively. CA, cellular activity; MA, mitochondrial activity; qPCR, quantitative polymerase chain reaction.
Details of Primers Used for the Quantification of Real-Time Polymerase Chain Reaction
In contrast, the total cellular activity from mitochondria was accessed using GAPDH normalization, since the GAPDH expression is inherent to each embryo cell. Relative quantification was performed in technical triplicate using a QuantStudio 3 Real-Time PCR System (Thermo Fisher Sci.). Reactions (15 μL total volume) contained the following: 2 × Fast SYBR Green Master Mix (7.5 μL; Applied Biosystems, Foster City, CA, USA), specific quantities of each primer pair (0.1 μM for 16S, 0.3 μM for GAPDH and ATP6, and 0.8 μM for COX and ND5), and cDNA. Reactions were heated at 95°C for 10 minutes, followed by 40 cycles of an amplification program of 95°C for 15 seconds and 60°C for 30 seconds.
Fluorescence signal was acquired during the 60°C extension steps. Specificity was ascertained after completion of the amplification in the presence of SYBR Green by cDNA melting curve analysis (55°C–95°C, starting fluorescence acquisition at 55°C and taking measurements at 10 seconds intervals until the temperature reached 95°C). Threshold and threshold cycle (Ct) values were automatically determined by Realplex 2.2 software (Eppendorf, Hamburg, Germany) using default parameters.
Statistical analyses
GraphPad InStat 7.0 (GraphPad Software, Inc., La Jolla, CA, USA) was used for all statistical analysis. Cleavage and blastocyst rates were compared using Fisher's exact test. The relative quantification of gene expression was performed using the 2−ΔΔCt method (Livak and Schmittgen, 2001). Expression profiles were expressed as mean (±standard deviation) of relative mRNA abundance and were compared using Kruskal–Wallis or Mann–Whitney test, when convenient. A value of p < 0.05 was set as the limit for statistical significance.
Results
In this study, a total of 124 and 777 oocytes from goats and cattle, respectively, were used for cloning. More cattle oocytes were used, owing to the greater local availability of cattle ovaries from local abattoirs. However, despite the paucity of goat ovaries, a total of 37 embryos were reconstructed. Furthermore, no significant differences (p > 0.05) were observed in the embryo development of the goat/goat and deer/goat groups (Table 2). However, it should be noted that the embryos did not reach the blastocyst stage (Fig. 2A, B).
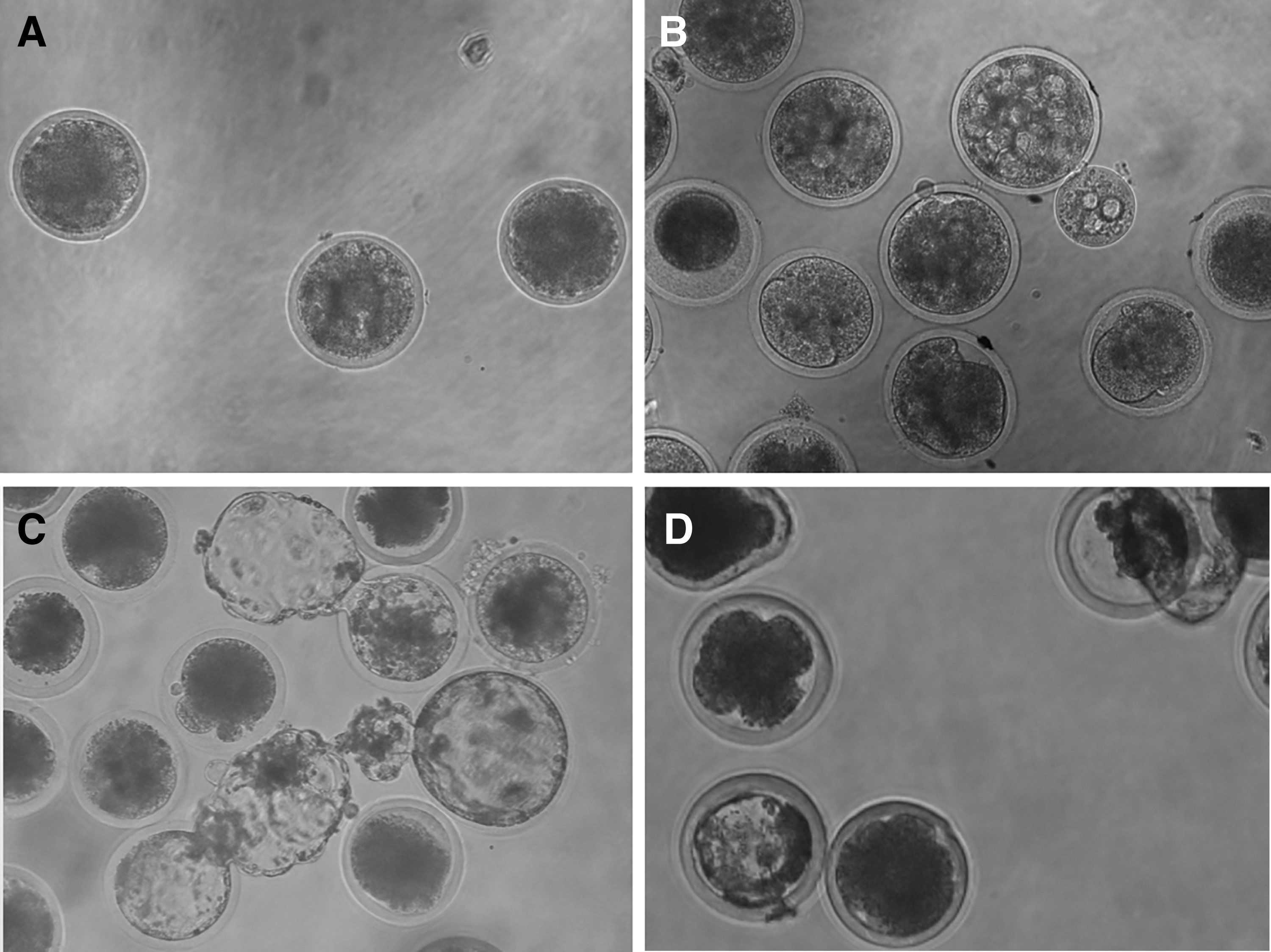
In vitro development of cloned embryos for the couples (fibroblast/cytoplast): goat/goat
In Vitro Development of Cloned Embryos Using Goat Cytoplast
Values with different superscripts in the same column are significantly different (p < 0.05). The data were obtained from one replicate.
In contrast, a total of 438 embryos were reconstructed using cattle oocytes. During the cleavage stage, there were no significant differences between the IVF and cattle/cattle groups (p > 0.05). However, the blastocyst rate of the IVF group was significantly greater than that of either the cattle/cattle or deer/cattle groups (p < 0.05; Table 3). When using cattle cytoplasts, reconstructed embryos reached the blastocyst stage (Fig. 2C, D).
In vitro Development of In Vitro Fertilized and Cloned Embryos Using Cattle Cytoplast
Values with different superscripts in the same column are significantly different (p < 0.05). The data were obtained from 2 (IVF) and 10 (cattle/cattle and deer/cattle) replicates.
IVF, in vitro fertilized.
Concerning the validation of qPCR amplification, parameters as efficiency and linearity, confirmed that the qPCR conditions were suitable for subsequent quantitative analysis (Table 4). Using goat cytoplast, no significant differences (p > 0.05) were observed in the ATP6 expression level of cloned embryos (goat/goat and deer/goat), irrespective of which reference gene was used (GAPDH or 16S; Fig. 3).

Relative quantification (±SD) of ATP6 in cloned embryos reconstructed with couples (fibroblast/cytoplast) of goat/goat and deer/goat, respectively. Relative quantification was observed separately for the reference genes (GAPDH and 16S). Different letters within a graph indicate a significant difference (p < 0.05). SD, standard deviation.
Quantitative Reverse Transcription Polymerase Chain Reaction Amplification Parameters for Gene Expression Analysis
SD, standard deviation.
When analyzing the MA per mitochondria, significantly greater COX3 expression was observed in the deer/cattle group than in the cattle/cattle group (p < 0.05). However, when analyzing MA per cell, the expression of all the evaluated genes was significantly greater in the deer/cattle group than in the IVF group (p < 0.05). Nevertheless, this difference was not observed (p > 0.05) when comparing deer/cattle and cattle/cattle groups for COX3 and ND5 (Fig. 4).
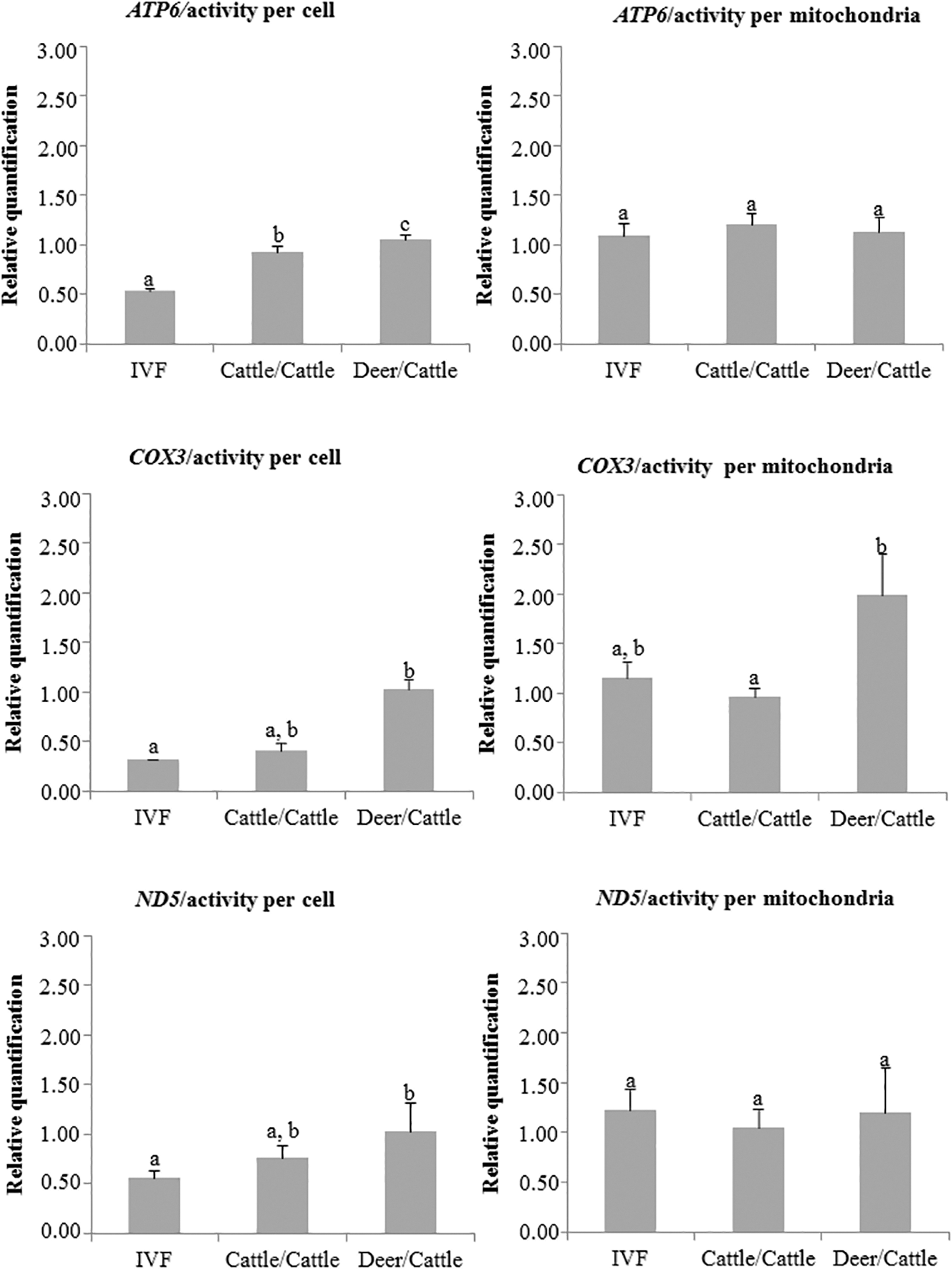
Relative quantification (±SD) of ATP6, COX3, and ND5 in in vitro fertilized (IVF) and cloned embryos reconstructed with couples (fibroblast/cytoplast) of cattle/cattle and deer/cattle, respectively. Relative quantification was observed separately for the reference genes (GAPDH and 16S). Different letters within a graph indicate a significant difference (p < 0.05).
Discussion
Interspecies cloning has been used successfully for a number of animals, including mouflon (Loi et al., 2001), wild cat (Gómez et al., 2004), wolf (Kim et al., 2007), sand cat (Gómez et al., 2008), bucardo (Folch et al., 2009), coyote (Hwang et al., 2012), gaur (Srirattana et al., 2012), and, more recently, Bactrian camel (Wani et al., 2017).
One of the main interests of interspecies cloning research involves the nuclear reprogramming of somatic cells, rather than simply addressing incompatibilities between the nucleus and receptor cytoplasm (Lagutina et al., 2011; Wang et al., 2011). More specifically, the hypothesis is that with less divergence between the nucleus of the donor species and the oocyte of the recipient species, there should be better communication between the nucleus and cytoplasmic elements, such as the mitochondrial genome (Bowles et al., 2007; Narbonne et al., 2011).
In this study, goat and cattle oocytes were chosen as cytoplasts to promote nuclear reprogramming in brown brocket deer. However, the goat/goat and deer/goat embryos only reached the morula stage. This may have been the result of the relatively low number of repetitions used in the experiment. However, Abdullah et al. (2011), who attempted to produce cloned goat embryos by SCNT and iSCNT, also reported that the in vitro development failed to overcome the morula stage. In fact, cloned goat embryos generally exhibit low in vitro developmental potential toward the late stage of preimplantation development (Das et al., 2003).
Similar to the previous study by Abdullah et al. (2011), the in vitro development rates of the SCNT and iSCNT goat embryos in this study did not differ at any of the cell stages, which suggests that goat cytoplasts could be used reconstruct embryos that develop beyond the morula if the embryos are transferred at earlier stages. Since the SCNT or iSCNT goat embryos produced in this study also presented similar patterns of mitochondrial gene expression, regardless of normalization method.
The results obtained with the cattle cytoplasts are also in agreement with previous studies (Lanza et al., 2000; Li et al., 2006; Matshikiza et al., 2004), in that the embryos reached the blastocyst stage. The reduced blastocyst rate observed in this study, when compared with embryos produced by IVF, could be related to the inability of maternal hereditary factors to activate the embryonic genome, inadequate demethylation of the donor genome, or nucleomitochondrial incompatibilities that could contribute to the early death of iSCNT embryos (Wani et al., 2017).
Evidence has shown that incompatibilities between the nuclear and mitochondrial genomes in iSCNT embryos negatively impact embryonic development (Mastromonaco et al., 2014). It has been proposed that proteins encoded by the nuclear genome are unable to recognize specific mitochondrial DNA sequences, owing to differences in taxonomic origin (Smith et al., 2005). Therefore, understanding the interactions between nuclear and ooplasmic compartments is essential to elucidating the mechanisms negatively affect the development or paresis of iSCNT embryos during early cleavage stages (González-Grajales et al., 2015).
The transfer of somatic cells from a deer to the cytoplast of other species has not been reported previously. The results of this study are consistent or better than those reported for other iSCNT embryos. For example, the blastocyst rates of this study are similar to those reported for cattle (7.5%; Uhm et al., 2007), panda (10.9%; Wen et al., 2005), and cat (5.8%; Wen et al., 2005) and numerically superior to those reported for tiger (1.0%; Hashem et al., 2007) and rhesus monkey (3.3%; Zhu et al., 2017).
The role of mitochondria in energy production is one of the critical factors in the normal development of gametes and embryos (Ramalho-Santos et al., 2009), especially cloned embryos, since mitochondria are mainly inherited from the recipient ooplasm, with only a small number provided by the donor cell (Mastromonaco et al., 2007). In this study, the goat/goat and deer/goat embryos exhibited similar patterns of gene expression, which demonstrated the occurrence of efficient nuclear reprogramming. However, when using cattle cytoplasts, the MA per cell differed among the groups, with greater gene expression observed in the deer/cattle group than in the IVF group. Meanwhile, the evaluation of MA per mitochondria verified that the gene expression patterns of the cattle/cattle and deer/cattle embryos were similar to those of the IVF group.
Similar findings concerning mitochondrial gene expression have been reported previously (González-Grajales et al., 2015). Based on these findings, two possible mechanisms were proposed to explain this higher expression. First, the improper silencing of genes by the oocyte soon after the transfer of the somatic cell might favor the propagation of somatic mitochondria during a stage when only oocyte-derived mitochondria should be maintained (Sutovsky et al., 2004). Second, it is possible that the expression of mitochondrial transcripts is a compensatory response to inadequate ATP reserves that result in premature mitochondrial biogenesis (González-Grajales et al., 2015). Therefore, it is clear that the production of ATP required for in vitro development was achieved and that efficient nuclear reprogramming occurred.
This study is the first to demonstrate the successful production of iSCNT embryos using karyoplasts from brown brocket deer. The results of the study indicate that both goat and cattle oocytes are capable of reprogramming the somatic cells of brown brocket deer and that the resulting embryos are able to develop until the morula and blastocyst stage, respectively. The success of iSCNT demonstrates the potential for its use in the preservation and dissemination of genetic diversity from small and fragmented populations of rare similar deer species. In this context, somatic cell banks should be promoted, and roadkill, confiscated animals, and hunted animals should all be used as sources of samples for embryo production.
Footnotes
Acknowledgments
The authors are very grateful to Fazenda Haras Claro (Caucaia, Brazil), which allowed the collection of samples from their animals. L.C.M. received a scholarship from CAPES, and V.J.F.F. and J.M.B.D. are CNPq senior investigators.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This study was supported by CNPq (grant no. 461330/2014-8) and CAPES/COFECUB (grant no. 88881.142966/2017-01).