
Abstract

To the Editor:
A recent clinical trial reported that autologous transplantation of retinal pigment epithelium differentiated from patients' own induced pluripotent stem cells (iPSCs) halted the progression of age-related macular degeneration (Mandai et al., 2017). However, the same study also reported genomic aberration of iPSCs derived from some aged donors (A-iPSCs), which deters the use of A-iPSCs of such patients (Mandai et al., 2017). Because aged donors are most likely beneficial from cell replacement therapy, genomic instability associated with A-iPSCs poses a significant risk in clinical settings.
Using mouse embryonic stem cells (ESCs, the gold standard for pluripotency) and A-iPSCs (derived from skin fibroblasts from 1.5-year-old adults) in a genetically controlled mouse model, we discovered that excessive glutathione production in A-iPSCs contributes to genomic instability of A-iPSCs. We also examined differences in other pluripotency-specific signatures in ESCs and A-iPSCs, including microRNA expression patterns (Fig. 1A, B).
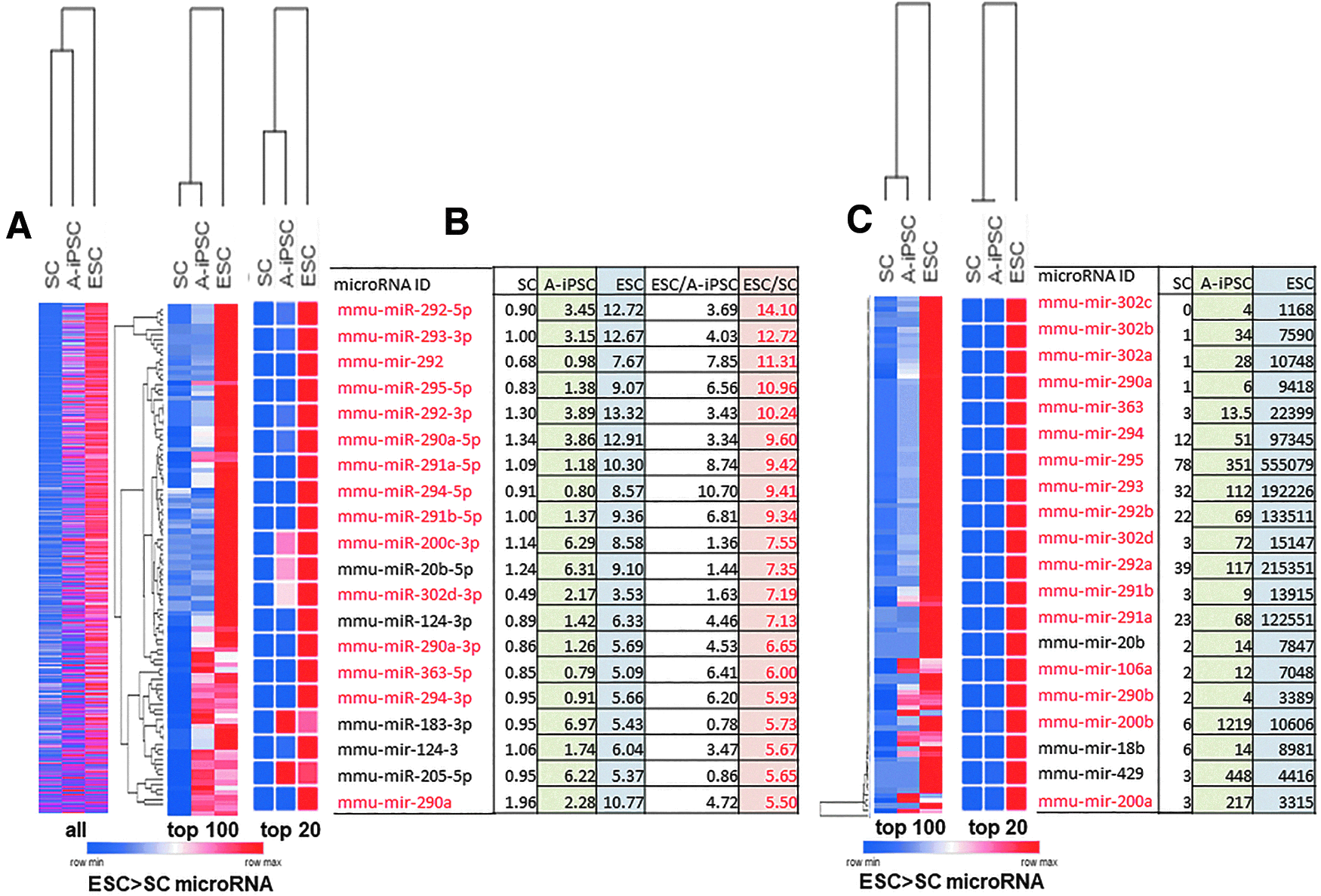
MicroRNA profiling analysis of SCs, A-iPSCs, and ESCs. (Refer to the Supplementary Data.)
We found that, despite expressing pluripotency markers and passing multiple pluripotency tests such as teratoma formation assays and chimera analysis (Skamagki et al., 2017), A-iPSCs exhibited an aberrant microRNA expression signature compared with ESCs (Fig. 1A, B). Our data demonstrated that A-iPSCs expressed lower levels of pluripotency-promoting microRNAs such as members of the miR-302, -106, -363, and -290/294, clusters (Fig. 1A, B).
MicroRNAs are small noncoding RNAs that play essential roles in early embryo development and tissue differentiation (Greve et al., 2013). Members of several microRNA families or clusters, such as miR-302, -106a, -363, or -290/294, have been shown to enhance iPSC reprogramming efficiency. A previous study also demonstrated that expression of the miR-302, -200c, -302, and -369, without the Yamanaka factors, is sufficient to achieve iPSC reprogramming (Greve et al., 2013).
However, the role of microRNAs as a whole during iPSC reprogramming has not been clearly demonstrated, with individual families or clusters acting as enhancers, sufficient sole drivers, or inhibitors. In addition, whether microRNAs are necessary for iPSC reprogramming was unclear. In our previous study, we demonstrated that somatic cells lacking Dgcr8, which encodes a factor required for the biogenesis of miRNAs, can be reprogrammed into iPSCs at reduced efficiencies by the Yamanaka factors (OCT4, SOX2, KLF4, and MYC) (Liu et al., 2015). These data, therefore, suggest that, although miRNAs facilitate reprogramming, microRNAs may be dispensable in converting somatic cells into iPSCs and the maintenance of iPSCs (Liu et al., 2015).
Our study unexpectedly discovered that A-iPSCs that pass standard pluripotency tests (Skamagki et al., 2017) could be obtained in the absence of the complete pluripotent stem cell-specific microRNA expression signatures present in ESCs (Fig. 1A, B). This microRNA signature was confirmed again using an RNA sequencing-based microRNA profiling method in addition to A-iPSC clones (Fig. 1C). These data support our previous discovery that microRNAs are not essential for somatic cell reprogramming (Liu et al., 2015).
Footnotes
Funding Information
K.K. is supported by NIH (Grant No. R00HL093212), NIH (Grant No. R01AG043531), TriStem-Star Foundation (2013-049), Louis V. Gerstner, Jr. Young Investigators Awards, Geoffrey Beene Junior Chair Award, Sidney Kimmel Scholar Award, Alfred W. Bressler Scholars Endowment Fund, and MSKCC Society Fund. MSKCC Core Facilities are supported by NIH Cancer Center Grant No. P30 CA008748. R.Z. is supported by NIH (Grant No. R01OD026594). H.L. is supported by NIH (Grant No. CA196631-01A1) and Mayo Clinic Center for Individualized Medicine.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.