
Abstract
The objective of this study was to examine the relationship between the timing of the first cleavage and in vitro development of somatic cell nuclear transfer (SCNT) embryos produced by different activation protocols. SCNT embryos were activated with calcium ionophore A23187 and further treated with 6-dimethylaminopurine (DMAP group), cycloheximide (CHX group), or anisomycin (ANI group). The proportion of SCNT embryos that cleaved within 18 hours after activation was significantly higher in the DMAP group (20%) than that in the CHX and ANI groups (3% and 2%, respectively). More than 70% of the cleaved embryos were observed within 24 hours in the DMAP and CHX groups, and within 26 hours in the ANI group. The blastocyst formation rate of SCNT embryos decreased gradually as the time from activation to the first cleavage increased in the DMAP group. The blastocyst formation rate of SCNT embryos cleaved at 22 hours (>20 to ≤22 hours) in the CHX group or within 26 hours in the ANI group was significantly higher than that of SCNT embryos cleaved more than 26 hours in each group. These results indicate that the activation protocol affects the timing of the first cleavage and subsequent in vitro development potential of bovine SCNT embryos and that late-cleaving embryos have a low developmental potential irrespective of the activation protocol.
Introduction
The timing of the first cleavage is an indicator of the subsequent developmental potential of embryos derived from in vitro fertilization (IVF) (Lechniak et al., 2008). Early-cleaving embryos are more developmentally competent in vitro than those that cleave relatively later after IVF in cattle (Lonergan et al., 1999), pigs (Isom et al., 2012), mice (Kobayashi et al., 2004), and humans (Fenwick et al., 2002). It has also been demonstrated that the transfer of early-cleaving embryos results in a higher pregnancy rate in human IVF and intracytoplasmic sperm injection (ICSI) (Lee et al., 2012; Lundin et al., 2001).
In somatic cell nuclear transfer (SCNT), several studies examined the relationship between the timing of the first cleavage and the in vitro development. In porcine SCNT, early-cleaving embryos have a higher blastocyst yield than late-cleaving embryos (Isom et al., 2012; Son et al., 2018). In contract, the potential to develop into blastocysts in intermediate-cleaving embryos is higher than in early- or late-cleaving embryos in mouse embryonic stem cell nuclear transfer (NT) (Kobayashi et al., 2004). In cattle, early-cleaving embryos derived from cumulus cells and intermediate-cleaving embryos from fibroblast cells have a high potential to develop into blastocysts (Amarnath et al., 2007), suggesting that the type of donor nucleus used for NT affects the timing of the first cleavage.
Artificial activation of recipient oocytes, which is an essential step in SCNT, plays a vital role in the proper development of embryos following SCNT (Alexander et al., 2006). In bovine SCNT, enucleated oocytes are fused electrically with donor cells followed by activation with calcium ionophore A23187 or ionomycin to elevate intracellular calcium levels. SCNT embryos are then treated with a protein kinase inhibitor or a protein synthesis inhibitor to inhibit maturation-promoting factor (MPF) activity (Alberio et al., 2001b; Whitworth and Prather, 2010). Treatment with 6-dimethylaminopurine (DMAP) for 3–4 hours (Cibelli et al., 1998; Galli et al., 1999; Wells et al., 1999) or cycloheximide (CHX) for 5–6 hours (Akagi et al., 2003; Kasinathan et al., 2001; Kato et al., 1998) is the most common method used in bovine SCNT.
Several reports suggest that these activation protocols affect in vitro development and ploidy of bovine SCNT embryos (Bhak et al., 2006; Galli et al., 2002; Shen et al., 2008). Anisomycin (ANI), a protein synthesis inhibitor, has previously been used to activate mouse oocytes (Liu et al., 2000). Recent studies demonstrate that the treatment of bovine oocytes with ANI for 5 hours results in a higher blastocyst formation rate than that with DMAP or CHX after ionomycin treatment in SCNT (Felmer and Arias, 2015) and ICSI (Arias et al., 2016). In addition, a lower percentage of chromosomal abnormalities has been observed with ANI-treated SCNT embryos compared to DMAP and CHX treatments (Felmer and Arias, 2015).
In a previous study, differences in the timing of the first cleavage are detected between bovine SCNT embryos treated with DMAP and CHX after ionomycin (Bhak et al., 2006). However, no study reports the relationship between the timing of the first cleavage and development potential of ANI-treated SCNT embryos. The objective of the present study was to investigate the relationship between the timing of the first cleavage and development potential of SCNT embryos produced by different activation protocols. Bovine SCNT embryos were activated with calcium ionophore A23187 followed by the treatment with DMAP, CHX, or ANI, and the time of the first cleavage and subsequent in vitro development of SCNT embryo from various time of first cleavage were examined.
Materials and Methods
All animal care and use protocols were implemented in accordance with the animal experimentation guidelines of the National Agriculture and Food Research Organization (NARO; Ibaraki, Japan). The study protocol was approved by the Animal Care and Use Committee at the Institute of Livestock and Grassland Science, NARO (Ibaraki, Japan; permit numbers: 1911C037).
Donor cells
Donor cells were prepared as previously described (Akagi et al., 2011). In brief, the ear skin tissue was obtained from a 3-year-old Japanese black cow. The tissue was finely chopped, and small pieces were seeded in Dulbecco's modified Eagle's medium (DMEM, Sigma-Aldrich, St. Louis, MO) with 10% fetal bovine serum (FBS), 100 IU/mL penicillin G potassium (Meiji Seika Pharma Co., Ltd., Tokyo, Japan), and 100 μg/L streptomycin (Meiji Seika Pharma Co., Ltd.) at 38.5°C in 5% CO2 atmosphere. The cultured fibroblast cells were frozen and stored at −80°C until use. Before NT, the fibroblast cells of passage 11–14 were cultured for 1–3 days after reaching confluency and dissociated by the treatment of 0.125% trypsin and EDTA buffer for 5 minutes.
In vitro maturation
Bovine cumulus-oocyte complexes (COCs) were collected from ovarian follicles of 2–6 mm in diameter. Only COCs with at least three layers of cumulus cells were selected for in vitro maturation (IVM). The IVM medium was TCM-199 (Gibco, Invitrogen, Grand Island, NY) with 10% FBS, 100 IU/mL penicillin G potassium, and 100 μg/mL streptomycin. Fifty to 70 COCs were cultured in each well of a four-well dish (Nunc, Thermo Fisher Scientific, Roskilde, Denmark) containing 700 μL of IVM medium for 18–20 hours under 5% CO2 atmosphere at 38.5°C.
Somatic cell nuclear transfer
The SCNT technique was performed as described previously (Akagi et al., 2014). In brief, cumulus cells surrounding the oocytes were removed by vortexing in 0.1% hyaluronidase for 5 minutes, and oocytes with a visible first polar body and homogeneous cytoplasm were selected under a dissection microscope for enucleation. Oocytes were enucleated using micromanipulators (MMO-202ND, Narishige, Tokyo, Japan) mounted on an inverted-microscope.
A first polar body and cytoplasm presumptively containing metaphase chromosomes were pushed out through a slit on the zona pellucida with a fine glass needle. Successful enucleation was confirmed by 5 mg/mL Hoechst 33342 (Sigma-Aldrich) staining the extruded cytoplasm under a fluorescence microscope. Donor cells were transferred to the perivitelline space of enucleated oocytes. The oocyte-cell complex was sandwiched with a pair of electrodes in Zimmerman mammalian cell fusion medium (Zimmerman and Vienken, 1982), and a single direct current pulse of 25 V was applied for 10 μs for oocyte-cell fusion 22–23 hours after IVM.
Chemical activation and embryo culture
The fused couplets were chemically activated 1 hour after fusion. Three different activation protocols were used. The fused couplets were exposed to 10 μM calcium ionophore A23187 (Sigma-Aldrich) for 5 minutes. Furthermore, fused couplets were treated with 2 mM DMAP (Sigma-Aldrich) for 4 hours in the DMAP group, 2.5 μg/mL cytochalasin D (CD) (Sigma-Aldrich), and 10 μg/mL CHX (Sigma-Aldrich) for 1 hour followed by 10 μg/mL CHX alone for 4 hours in the CHX group, or 2.5 μg/mL CD and 1 μg/mL ANI (Sigma-Aldrich) for 1 hour followed by 1 μg/mL ANI alone for 4 hours in the ANI group. After chemical activation, the fused couplets were cultured in a serum-free medium (IVD-101, Research Institute for the Functional Peptides, Yamagata, Japan; Hoshi, 2003) under 5% O2, 5% CO2, and 90% N2 atmosphere at 38.5°C.
SCNT embryos were examined at 16, 18, 20, 22, 24, 26, and 48 hours after the start of chemical activation (Amarnath et al., 2007). At each time point, each of the embryos with two or more distinct blastomeres was transferred into a microwell culture dish well (Dai Nippon Printing Co. Ltd., Tokyo, Japan; Sugimura et al., 2010) and cultured in fresh IVD101 until day 7 (day 0 = the day of NT).
Blastocyst cell number
Blastocysts were stained with 25 μg/mL Hoechst 33342 in 99.5% ethanol for 2 hours on day 7. After washing with glycerol, the blastocysts were mounted on slides, gently flattened with coverslips, and examined under UV light using a fluorescence microscope. A digital image of each embryo was taken, and cell numbers were counted using the ImageJ software (National Institutes of Health, Bethesda, MD).
Statistical analysis
The experiments were repeated five times in each group. Values are presented as the mean ± standard error of the mean. Statistical analysis was performed after arcsine transformation of the percentage data. Differences were analyzed using the general linear model procedure of SAS (Statistical Analysis System Institute, Inc., Cary, NC) for analysis of variance and Tukey's test. p < 0.05 was considered statistically significant.
Results
The cleavage and blastocyst formation rates and the blastocyst cell number of SCNT embryos are shown in Table 1. There was no significant difference in cleavage rates between the DMAP, CHX, and ANI groups. However, the blastocyst formation rates of the CHX and ANI groups (49.4% ± 3.9% and 49.2% ± 2.1% respectively) were significantly higher than that of the DMAP group (23.2% ± 1.1%). In addition, the blastocyst cell number in the CHX and ANI groups (132.0 ± 4.8 and 133.1 ± 3.3, respectively) was higher than that in the DMAP group (113.4 ± 6.3).
In Vitro Development of Somatic Cell Nuclear Transfer Embryos Activated by Different Protocols
Values are expressed as the mean + SEM.
Values without common characters in the same column differ significantly (p < 0.05).
ANI, anisomycin; CHX, cycloheximide; DMAP, dimethylaminopurine; SEM, standard error of the mean.
Three embryos (1.9% ± 1.8% of cleaved embryos) cleaved at 16 hours after the start of chemical activation in the DMAP group (Fig. 1A), but no cleavage was observed at 16 hours in the CHX and ANI groups (Fig. 1B, C).
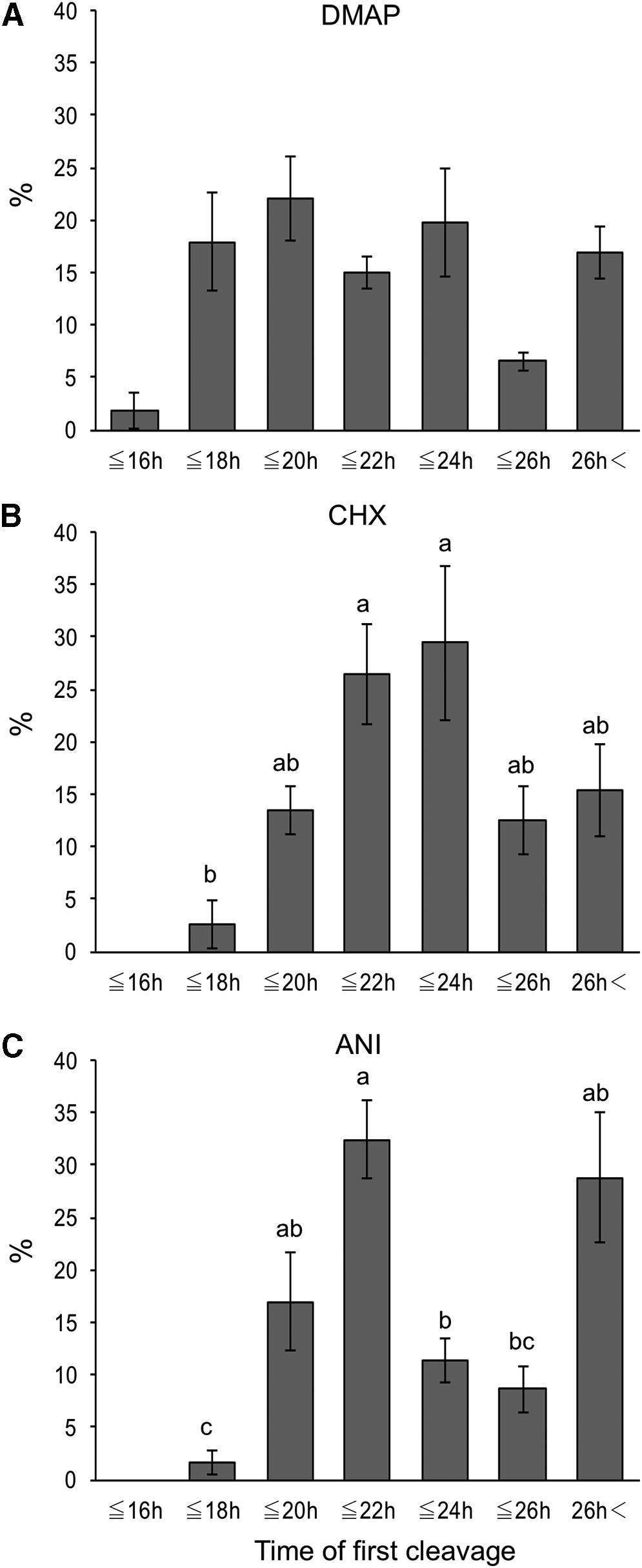
The timing of the first cleavage of SCNT embryos activated with 6-dimethylaminopurine (DMAP group), cycloheximide (CHX group), or anisomycin (ANI group).
In the DMAP group, 62.9% of embryos cleaved within 18 hours after activation developed to the blastocyst stage, and the blastocyst formation rate of SCNT embryos decreased as the time required to complete the first cleavage increased (Fig. 2A).
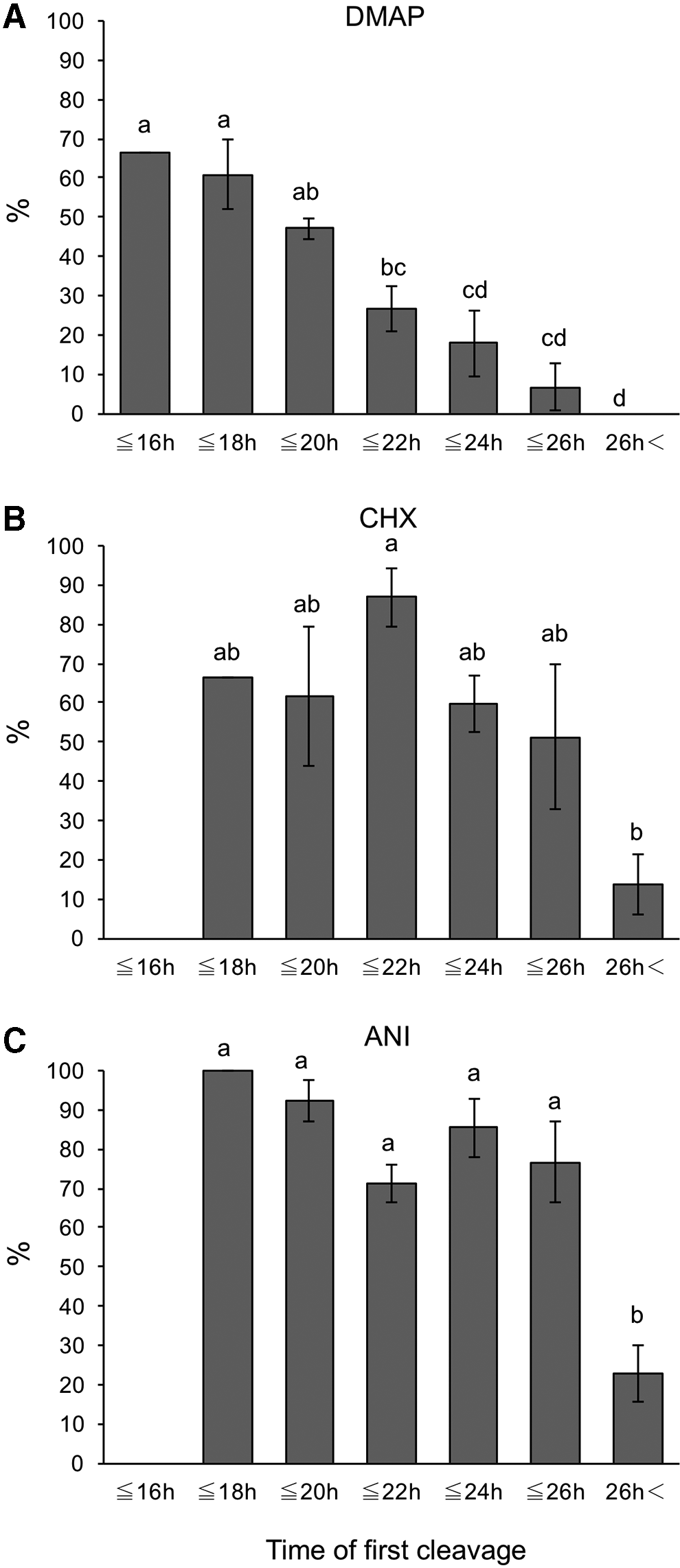
The blastocyst formation rate of SCNT embryos activated with 6-dimethylaminopurine (DMAP group), cycloheximide (CHX group), or anisomycin (ANI group).
In the CHX group, the proportion of SCNT embryos that cleaved at 22 hours (>20 to ≤22 hours) and 24 hours (>22 to ≤24 hours) tended to be high (Fig. 1B), and the blastocyst formation rate of SCNT embryos cleaved at 22 hours (87.0% ± 7.4%) was significantly higher than that of SCNT embryos cleaved more than 26 hours (13.5% ± 7.7%) (Fig. 2B).
In the ANI group, more than 30% of cleaved embryos were observed at 22 hours (Fig. 1C). Moreover, SCNT embryos that cleaved within 26 hours had higher blastocyst formation rate in the ANI group (Fig. 2C).
When the proportion of SCNT embryos that cleaved at the same time point was compared among the three groups, the proportion of embryos that cleaved within 18 hours in the DMAP group (19.8% ± 6.1%) was significantly higher than that in the CHX and ANI groups (2.6% ± 2.3% and 1.7% ± 1.2%, respectively), whereas the proportion of embryos that cleaved at 22 hours in the ANI group (32.5% ± 3.6%) was significantly higher than that in the DMAP group (15.0% ± 1.5%).
When the proportion that developed to blastocysts at the same time point was compared among the three groups, SCNT embryos that cleaved at 22 and 24 hours in the CHX and ANI groups had significantly higher blastocyst formation rates (87.0% ± 7.4% and 59.8% ± 7.3% in the CHX group, and 71.2% ± 4.8% and 85.4% ± 7.5% in the ANI group, respectively) than those in the DMAP group (26.7% ± 6.0% and 17.8% ± 8.2%, respectively). There was no significant difference in the cell number of blastocysts at each time point among the three groups (Table 2).
Relationship Between the Timing of the First Cleavage and Blastocyst Cell Number Of Somatic Cell Nuclear Transfer Embryos Activated by Different Protocols
Values are expressed as the mean ± SEM.
Discussion
In SCNT, the lack of sperm-induced fertilization requires artificial activation to initiate further embryonic development. Bovine SCNT embryos are usually activated by chemicals that induce an increase in cytoplasmic calcium levels followed by inhibitors of protein synthesis or kinase activity to prevent reactivation of MPF. In the present study, three chemicals (DMAP, CHX, and ANI) were used to inhibit MPF activity in SCNT embryos after treatment with calcium ionophore A23187. DMAP and CHX have been widely used in the successful production of cloned cattle (Galli et al., 2002; Whitworth and Prather, 2010). Thus, comparisons between the two chemicals after calcium ionophore A23187 or ionomycin treatment have been performed in previous studies.
Several reports found no significant difference between DMAP- and CHX-treated SCNT embryos on in vitro development (Bhak et al., 2006; Felmer and Arias, 2015; Ross et al., 2009). On the contrary, Shen et al. (2008) demonstrated that the blastocyst formation rate of bovine SCNT embryos activated with DMAP was higher than that of SCNT embryos activated with CHX. In the present study, however, the blastocyst formation rate in the CHX group was significantly higher than that in the DMAP group. In addition, the in vitro developmental potential of the ANI group was the same as that of the CHX group, which is inconsistent with a previous report by Felmer and Arias (2015).
The blastocyst formation rates of SCNT embryos in the CHX and ANI groups were 49%, which is relatively high compared with those of IVF embryos incubated under the same culture condition (Takeda et al., 2019). There were several differences in the SCNT embryo production protocols between our study and previous studies, such as donor cell types (e.g., fetal or adult) and conditions (e.g., serum-starved or confluent cells), oocyte maturation, and embryo culture. However, the reason behind the difference between our results and previous studies on in vitro development by various activation protocols is not apparent, as many factors contribute to embryonic development after SCNT.
In the present study, activation protocols affect the timing of the first cleavage in bovine SCNT. The first cleavage in SCNT embryos was observed earlier in the DMAP group than in the CHX and ANI groups. As shown in Figure 1, 20% of cleaved embryos have already cleaved 18 hours after activation in the DMAP group. On the contrary, the proportions of embryos that cleaved within 18 hours were only 3% and 2% in the CHX and ANI groups, respectively. Similar results were obtained with bovine parthenogenetic embryos. Holm et al. (2003) reported that the first cleavage of DMAP-treated parthenogenones occurs earlier than that of CHX-treated parthenogenones and suggested that the delayed cleavage in CHX-treated parthenogenones may be a result of the temporary inhibition of de novo protein synthesis, which is necessary for proper cleavage, during CHX treatment.
DNA synthesis during the first cell cycle is influenced by the activation treatment (Alberio et al., 2001a; Ledda et al., 1996; Soloy et al., 1997). In the present study, the initiation of DNA synthesis may be delayed in SCNT embryos activated by CHX or ANI. In the previous report, the delayed initiation of DNA synthesis is observed in SCNT embryos activated by ethanol and CHX (Alberio et al., 2001a). However, further studies are required to elucidate the mechanisms responsible for the differences in the timing of the first cleavage by activation protocols.
In the present study, we investigated the timing of the first cleavage and the subsequent in vitro developmental potential of SCNT embryos produced by different activation protocols. In the DMAP group, early-cleaving embryos had higher developmental potential than late-cleaving embryos. Consistent with previous results observed in bovine IVF embryos (Amarnath et al., 2007; Dinnyes et al., 1999), the potential of SCNT embryos to develop into blastocysts decreased as the time from the activation to the first cleavage increased. No embryo that cleaved more than 26 hours developed to a blastocyst.
In the CHX group, SCNT embryos that cleaved within 26 hours were more likely to develop to the blastocyst stage than those that cleaved more than 26 hours. The blastocysts from SCNT embryos that cleaved within 22 hours showed high cell number (>140). In the ANI group, a high blastocyst formation rate (more than 70%) was observed in SCNT embryos that cleaved within 26 hours, while only 23% of SCNT embryos that cleaved more than 26 hours developed to the blastocyst stage. These results suggest bovine SCNT embryos that cleaved more than 26 hours have a low developmental potential in vitro irrespective of the activation protocols. The exact reason that late-cleaving embryos have a lower developmental potential is not apparent.
However, late-cleaving bovine SCNT embryos may have a higher incidence of DNA damage than early-cleaving embryos. Bohrer et al. (2015) demonstrated that the incidence of DNA double-strand breaks is higher in late-cleaving and developmentally less-competent parthenogenetic embryos.
In general, the selection of bovine embryos used for transfer is based on morphological quality. The morphologic appearance of SCNT blastocysts is not different from fertilized embryos (Tsunoda and Kato, 2002). However, the calving rate after embryo transfer is very low for SCNT (Akagi et al., 2013). Therefore, new criteria for the selection of embryos for transfer are required to improve the in vivo developmental potential of SCNT embryos. In the present study, the results indicated that the timing of the first cleavage affects the subsequent in vitro development potential of bovine SCNT embryos.
It has been reported in bovine IVF that there are differences between early- and late-cleaving embryos in terms of gene expression (Fair et al., 2004), mRNA polyadenylation (Brevini et al., 2002), and chromosomal abnormality (Sugimura et al., 2012), as well as in vitro developmental potential. Reportedly, the in vivo developmental potential after the transfer of early-cleaving IVF embryos is higher than that of late-cleaving embryos in human (Lee et al., 2012; Lundin et al., 2001) and mouse (Lee et al., 2015). However, we did not examine the posttransfer viability of SCNT blastocysts from various time of the first cleavage. To evaluate whether the timing of first cleavage can be used as one criterion for the selection of viable embryos, further studies are necessary to examine the viability of SCNT embryos after transfer.
Footnotes
Author Disclosure Statement
The authors declare they have no competing financial interests.
Funding Information
This work was supported, in part, by JSPS KAKENHI Grant no. JP18H03953.