
Abstract
Chicken embryonic stem cells (cESCs) isolated from the egg at the stage X hold great promise for cell therapy, tissue engineering, pharmaceutical, and biotechnological applications. They are considered to be pluripotent cells with the capacity to self-renewal and differentiate into specialized cells. However, long-term maintenance of cESCs cannot be realized now, which impedes the establishment of cESC line and limits their applications. Therefore, the separation locations, isolation methods, and culture conditions especially the supplements and action mechanisms of cytokines, including leukemia inhibitory factor, fibroblast growth factor, transforming growth factor beta, bone morphogenic protein, and activin for cESCs in vitro, have been reviewed here. These defined strategies will contribute to identify the key mechanism on the self-renewal of cESCs, facilitate to optimize system that supports the derivation and longtime maintenance of cESCs, establish the cESC line, and develop the biobank of genetic resources in chicken.
Introduction
Embryonic stem cells (ESCs) derived from the inner cell mass of blastocyst in mammals have been a favorable tool for the developmental biology, regenerative medicine, and applied sciences. ESCs own unique characteristics, including self-renewal and the ability to generate the endoderm, mesoderm, and ectoderm both in vitro and in vivo (Gazina et al., 2018; Lorzadeh and Kazemirad, 2018). Although considerable efforts to obtain ESC lines have been done, the ESC lines are only established in mouse (Evans and Kaufman, 1981), rat (Iannaccone et al., 1994), primate (Thomson et al., 1995), human (Thomson et al., 1998), and cattle (Bogliotti et al., 2018).
Cells isolated from blastoderm of fertilized egg at embryonic stage X in chicken are usually termed as chicken embryonic stem cells (cESCs) (Sun et al., 2018), and this kind of cell has relatively small amount of cytoplasm and a large translucent nucleus (Zhang et al., 2013). cESCs share many characteristics of self-renewal and multilineage differentiation with ESCs in mammal; they can express some pluripotent markers such as alkaline phosphatase (AKP) and stage-specific embryonic antigen-1 (SSEA-1) and generate teratoma with three germ layers in vivo and embryoid body (EB) in vitro (Boast and Stern, 2013). cESCs have provided an ideal opportunity to test cytotoxicity of newly developed drugs, generate cell-based transgenic birds, produce vaccines for poultry and human viral diseases, provide insights into basic researches, and so on (Han et al., 2015).
With the continuous improvements of culture system, chick blastodermal cells could be harvested and maintained with their native characteristics in vitro (Lopez-Diaz et al., 2016). But the cESC line has not been established because it is very difficult to maintain their undifferentiated state for long time in vitro and produce germ-line chimeras (Aubel and Pain, 2013). It is necessary to eliminate the variations in culture conditions to develop standard protocols in the culture of cESCs for gene manipulation and biological researches. Therefore, various approaches and suggestions are reviewed here to optimize the isolation strategy and define longtime culture system of cESCs.
Isolation of cESCs
Blastoderm cells (BCs) of chicken embryo at stage X are similar to mouse embryonic stem cells (mESCs), which contribute to generate all types of somatic tissues (Eyal-Giladi, 1984; Zhang et al., 2013). Unlike mESCs, chicken blastoderm is found in egg, which facilitates the isolation and culture of cESCs in vitro (Lavial and Pain, 2010). After blastoderms are washed with phosphate buffer saline (PBS) and mechanically dispersed, cESCs can be harvested.
Four kinds of strategies are commonly used to collect blastoderm (Fig. 1). (I) Spoon method. After the blastoderm is cut around with scissor, the embryonic disc is scooped up, and the yolk is peeled off. This procedure itself is simple and convenient, but it is a pity that more yolk is also carried (Alev et al., 2013; Man et al., 2011); (II) Paper ring method. The paper ring is directly placed on the embryonic disc; after being cut along the outer circumference of the paper ring, the internal round paper attached with embryonic disc is gently extracted (Horiuchi et al., 2006). This method is easier for removing the blastoderm from the yolk, the paper ring needs to be replaced when each embryonic disc is extracted, so a certain number of filter paper rings should be prepared; (III) Hair ring method.
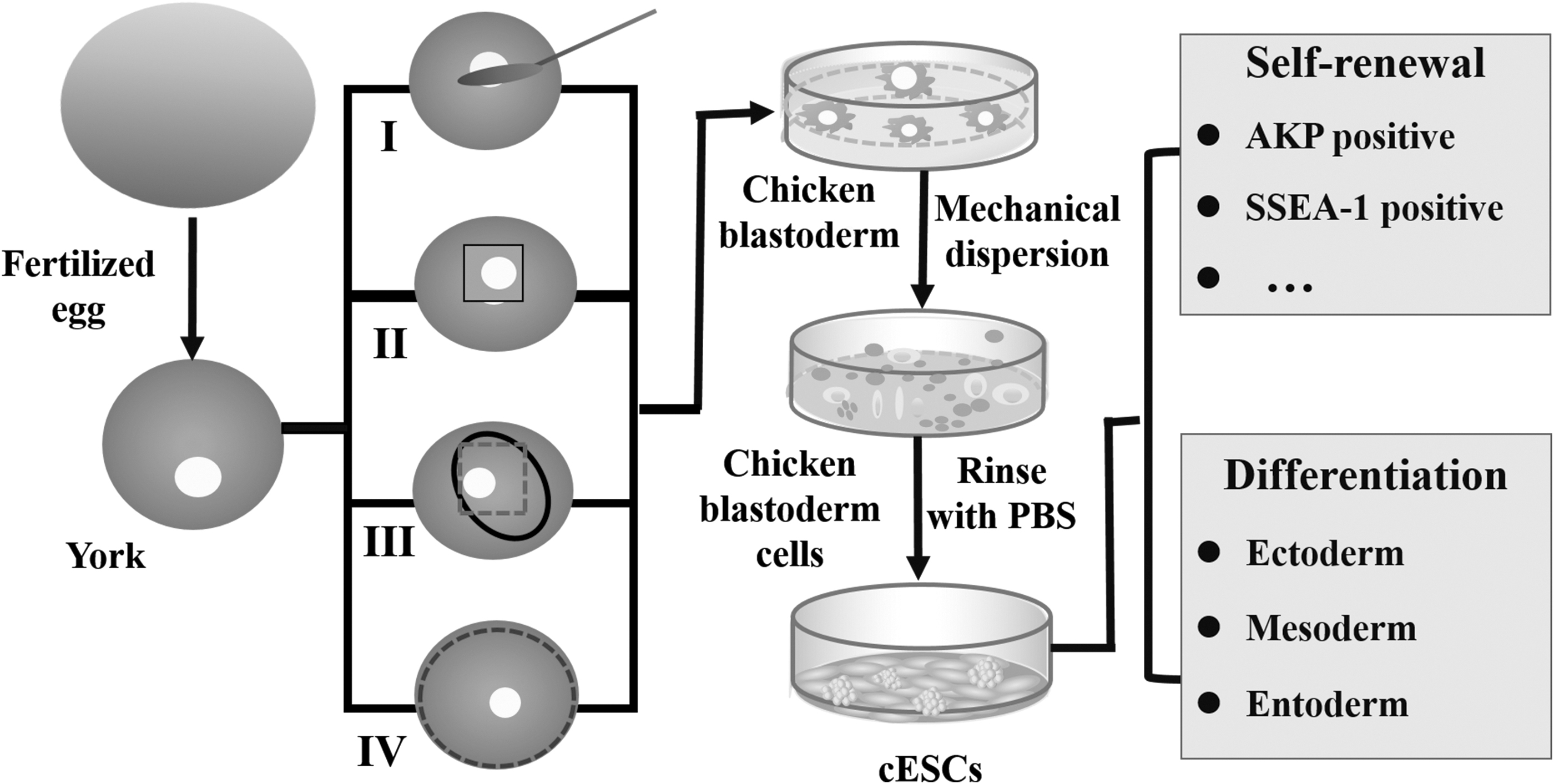
Schematic diagram of different methods for the isolation of cESCs. (I) Spoon method; (II) Paper ring method; (III) Hair ring method; (IV) Whole embryo extraction method. cESCs, chicken embryonic stem cells.
The blastoderm is gently cut off directly using scissor, and then the yolk is gently peeled off with the hair ring (Du and An, 2003; Etches et al., 1997). Compared to the other strategies, this method is more sophisticated, the precise control of hair ring in strength and intensive labor during isolation are required; it is not suitable for extracting large amounts of embryonic disc under laboratory condition; (IV) Whole embryo extraction method. The embryonic disc is isolated with scissor, followed by being gently lifted from the outside with ophthalmic tweezer (Alev et al., 2013). The intact embryonic disc can be obtained; furthermore, the yolk attached to the disc is easy to be stripped. The characteristics of these separation methods are summarized in Table 1.
Characteristics of Methods for Isolating Chicken Blastoderm Cells
+, very simple; ++, simple; +++, difficult; ++++, very difficult; *, incomplete; **, relatively complete; ***, complete; ****, very complete; #, relatively clean; ##, clean; ###, dirty; ####, very dirty.
In addition to these isolation strategies mentioned above, it is vital to prevent cESCs from being contaminated, because the eggshell and egg yolk which adhered to the embryonic disc are the main carriers of the contaminant. Wiping with 70% alcohol or soaking in formaldehyde is often used to interdict the contact between microbes and the embryonic disc.
Compared with the other three methods, the hair ring method is more capable of preventing the contamination, because less yolk was carried, but to keep a sterile environment, penicillin, streptomycin, and gentamicin are often added in culture medium (Alev et al., 2013; Du and An, 2003). The blastoderm isolated with the spoon method or the whole embryo extraction method will be easily contaminated due to a large amount of egg yolk, so it is crucial to rinse the blastoderm with PBS. On account of rare yolk, the paper ring method can effectively avoid the contamination.
Factors Influencing Long-Term Culture of cESCs In Vitro
Separation site
When being observed dorsally, the chicken embryonic disc is usually divided into three different regions (Fig. 2A) as follows: (1) Pellucida zone, a more transparent central area that overlies the subgerminal space; (2) Opaca zone, the more dense area that consists of the peripheral BCs, which remains in contact with the yolk; and (3) Junctional zone, a region at the border of these two areas. The isolation efficiency of cESCs was associated with the different sites of chicken embryonic disc.

Separation sites and isolation efficiency of cESCs.
Compared to other two regions, cells isolated from the pellucida zone showed a better capacity in terms of growth, proliferation, and maintenance of cESC characteristics such as expression of AKP (Fig. 2B). In addition, colonies found in the pellucida area had more complete cells; they held larger potential for subsequent growth and generation of much more positive clones during culture in vitro (Horiuchi et al., 2006; Kessel and Fabian, 1986; Khaner et al., 1985).
Growth factors and cytokines
The long-term culture of cESCs in vitro is mainly dependent upon several specific growth factors and cytokines; some commonly used factors are listed in Table 2.
Summary of Factors Associated with “Self-Renewal” or Differentiation of Chicken Embryonic Stem Cells In Vitro
Akt, protein kinase B; bFGF, basic fibroblast growth factor; BMP, bone morphogenic protein; cESC, chicken embryonic stem cell; cPGCs, chicken primordial germ cells; ERK, extracellular signal-regulated kinase; IGF, insulin-like growth factor; JAK, Janus kinase; LIF, leukemia inhibitory factor; MAPK, mitogen-activated protein kinase; MEK, MAPK kinase; PI3K, phosphoinositide 3-kinase; SCF, stem cell factor; SSCs, spermatogonial stem cells; SMAD1/5/8, drosophila mothers against decapentaplegic protein1/5/8; STAT3, signal transducer and activator of transcription 3; TGFβ, transforming growth factor beta.
Leukemia inhibitory factor
Leukemia inhibitory factor (LIF) is a member of the interleukin-6 (IL-6) family which also includes ciliary neurotrophic factor, IL-6, and interleukin-11 (IL-11) (Martello et al., 2013). It was initially identified as an inducer for the functionalization of M1 leukemia cells and then was discovered as an inhibitor to the differentiation of mESCs (Pease et al., 1990).
As a pivotal activator for the self-renewal of ESCs, LIF is mainly involved in three different signaling pathways, including SH2 domain-containing tyrosine phosphatase 2/mitogen-activated protein kinase signaling pathway (SHP2/MAPK signaling pathway), phosphoinositide 3-kinase/protein kinase B signaling pathway (PI3K/Akt signaling pathway), and Janus kinase/signal transducer and activator of transcription 3 signaling pathway (JAK/STAT3 signaling pathway) (Onishi and Zandstra, 2015). The cascade reactions of these signal pathways mediated by LIF are shown in Figure 3.
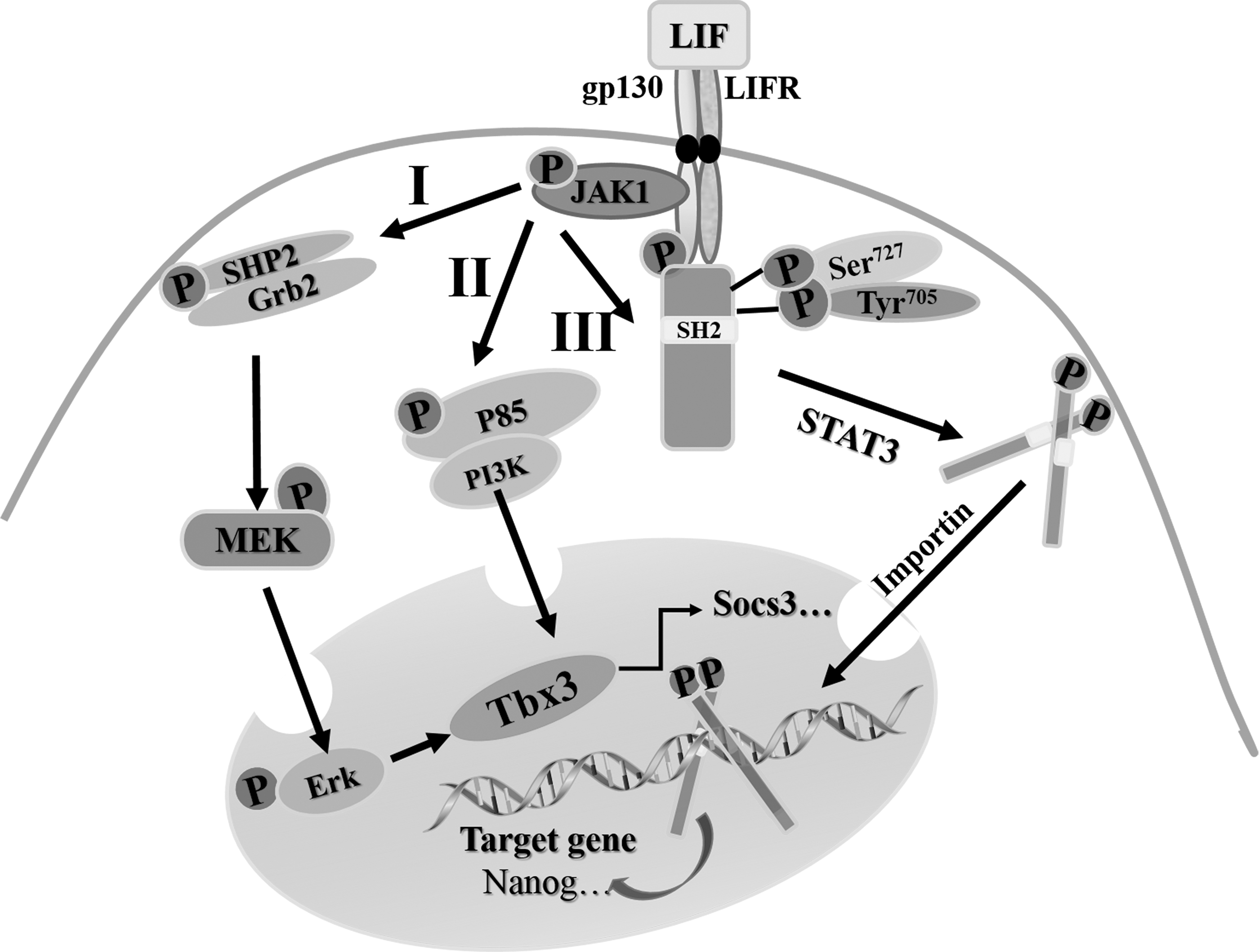
LIF-mediated signaling pathway involved in maintenance of cESC pluripotency. I. LIF/MEK/ERK signaling pathway. The activation of JAKs stirs the union of SHP2 and Grb2 and their phosphorylation, then the MEK is phosphorylated and translocated into the nucleus to give rise to the incitant of ERK after the stimulation of the union of SHP2 and Grb2. II. LIF/PI3K signaling pathway. The combination between the activated JAKs and p85 initiates the excitation of PI3K, then the ally of p85 and PI3K is formed. III. LIF/STAT3 signaling pathway. Binding of LIF to its membrane receptor leads to the activation of JAKs and phosphorylation of STAT3. The phosphorylated STAT3 dimers interact with reciprocal SH2, then the dimer of STAT3 is translocated into the nucleus, regulating the gene transcription and controlling the cell behaviors. ERK, extracellular signal-regulated kinase; Grb2, growth factor receptor-bond protein 2; JAK, Janus kinase; LIF, leukemia inhibitory factor; MEK, MAPK kinase.
LIF works through a cell surface receptor complex which is composed of the low affinity leukemia inhibitory factor receptor (LIFR) and gp130. gp130 is a common receptor subunit for the member belonging to IL-6 cytokine family. LIFR and gp130 form heterodimer and activate associated tyrosine kinases such as members of JAK family; JAKs subsequently phosphorylate the tyrosine residues in their receptors, which in turn provide the critical docking sites for recruitment of SH2, p85, and cytoplasmic STAT3 monomer (Huang et al., 2015; Ye et al., 2016).
The dimerization of LIFR/gp130 receptor leads to the phosphorylation of p85 and SHP2, which activates the PI3K/Akt and SHP2/MAPK pathways, respectively. The binding of activated p85 and PI3K will form a coalition, then SHP2 can unite growth factor receptor-bond protein 2 (Grb2) to agitate for the phosphorylation reaction of MAPK kinase (MEK) (Tai and Ying, 2013). Recruited STAT3 molecules become their own substrates for JAK-mediated phosphorylation, the dimer of STAT3 combines with reciprocal SH2 to form a conjugate, and then the phosphorylated MEK, coalition of p85 and PI3K, and conjugate of STAT3 are translocated into the nucleus, where the homodimers activate the transcription of target genes (Sutherland et al., 2018).
JAK/STAT3 pathway represents one of the common mechanisms on how extracellular signaling proteins regulate the transcription of pluripotent genes (such as Nanog and Oct4) and control the behaviors of ESCs (Herrera and Bach, 2019). LIF could not support the self-renewal of STAT3−/− ESCs; it has been verified that STAT3 could also be phosphorylated on two different sites as follows: Tyr705 and Ser727 (Hirai et al., 2011). In STAT3-mediated self-renewal of ESCs, the phosphorylation of Tyr705 is essential; however, Ser727 phosphorylation is dispensable, which just enhances the transcriptional activity of STAT to achieve proliferation and optimal pluripotency of ESCs (Huang et al., 2014). Furthermore, the phosphorylation of Ser727 is critical in the transition from self-renewal to neuro-commitment, which suggests that the phosphorylations of Tyr705 and Ser727 in STAT3 are dynamically balanced in controlling the fate of ESCs (Zhang et al., 2017).
The inhibitory effect of LIF on the differentiation of cESCs was obvious, especially in the passage culture medium supplemented with 1000 IU/mL LIF; it was found that the AKP-positive clones could be maintained for 5 days (Du and An, 2003). When the recombinant chicken LIF consisted of 211 amino acids and was generated from the chicken monocytic cell line (IN24) cells were added to culture cESCs; the expression of AKP and SSEA-1, as well as the activation of STAT3, was observed (Nakano et al., 2011; Zhang et al., 2016b). Meanwhile, basic culture medium supplemented with 20 ng/μL human basic fibroblast growth factor (hbFGF), 20 ng/μL human LIF (hLIF), and 20 ng/μL mouse stem cell factor (mSCF) could stimulate the growth and self-renewal of cESC colonies (Zhang et al., 2018).
Fibroblast growth factor
FGF is a kind of polypeptides that transmit the developmental signal and promote cell division in mesoderm and neuroectoderm. FGF4 is one of the important growth factors expressed throughout the early development of embryo, and it works mainly through PI3K/Akt signaling pathway and rat sarcoma/MEK/extracellular signal-regulated kinase signaling pathway (RAS/MEK/ERK signaling pathway) in maintaining the pluripotency of ESCs (Li et al., 2007; Rao, 2004). Fibroblast growth factor receptor substrate 2 (FRS2) can mediate the activation of PI3K/Akt signaling pathway, and Grb2 has the capability of activating RAS/MEK/ERK signaling pathway (Mark et al., 2008).
FGF signaling is triggered by an interaction between ligands and receptors, leading to the auto-phosphorylation of intracellular tyrosine residues in fibroblast growth factor receptor (FGFR) (Miles et al., 2013). Grb2 couples with the FGFR to activate the MAPK/ERK pathway, which results in Ras-phosphorylation, and in turn the phosphorylation of Ras triggers phosphorylation of MEK. Under the regulation of MEK, the ERK is phosphorylated, and finally the phosphorylated ERK is shifted into the nucleus to initiate the expression of pluripotent genes such as Nanog and Oct4 (Eiselleova et al., 2009; Kunath et al., 2007). FGF has been shown to cooperate with Activin/drosophila mothers against decapentaplegic protein2/3 (Smad2/3) signaling pathway to maintain high level of Nanog expression in human ESCs and activate PI3K/Akt signaling pathway to enhance proliferation and survival of human ESCs (Fig. 4) (Zhang et al., 2019).
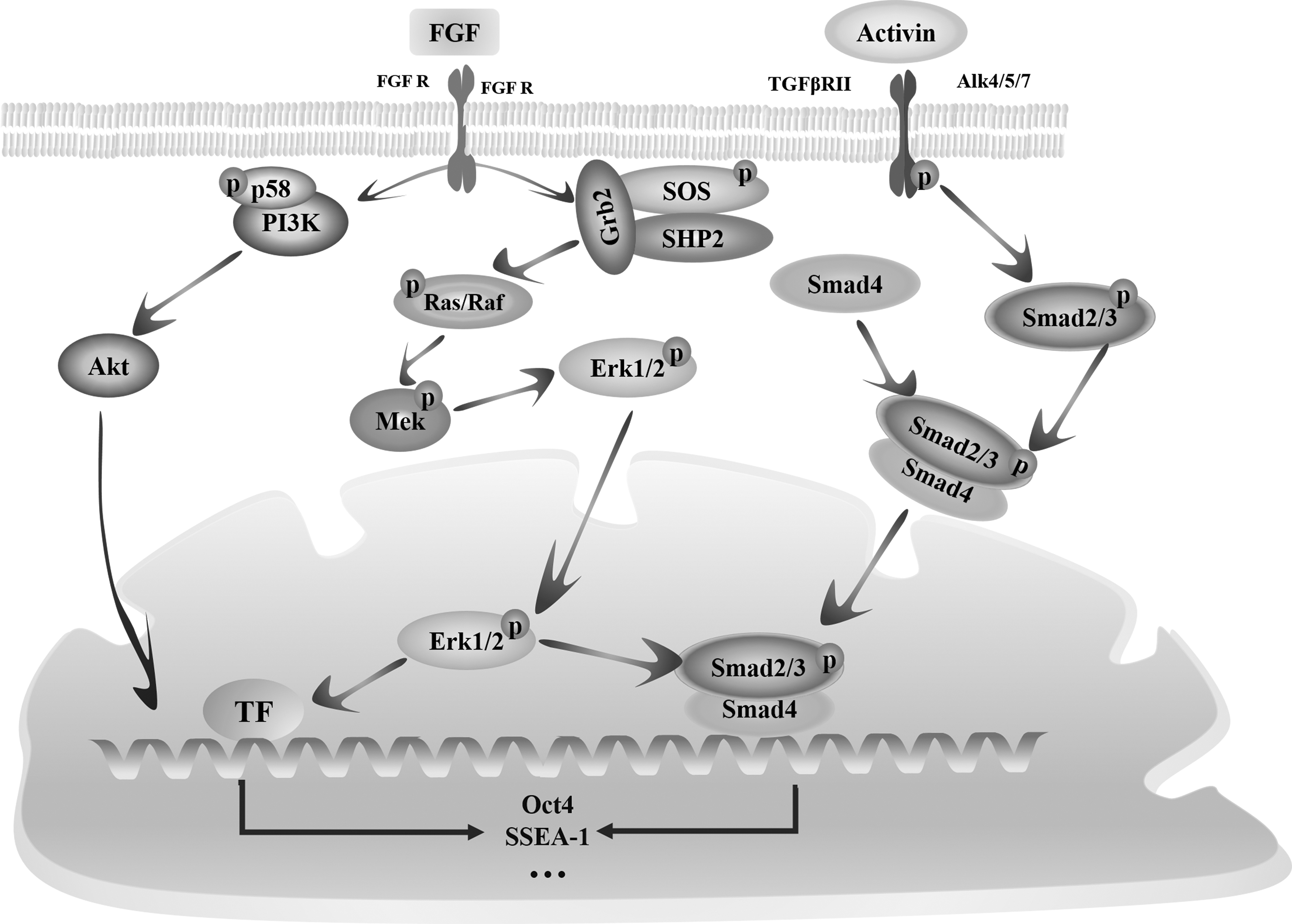
FGF-and Activin-mediated signaling pathways involved in the maintenance of cESCs in vitro. When FGF binds to FGFR, which can not only promote the Grb2, SOS, and SHP2 to form a solidarity but also activate the union of p58 and PI3K. Then the RAS/RAF/MEK/ERK signaling pathway is cut through after the irritation of Grb2/SOS/SHP2, the clique of p58 and PI3K makes Akt phosphorylated, and the phosphorylated Akt is translocated into the nucleus. Activin arouses the cascade reactions of Smad proteins after binding to TGFβR/Alk heterodimer receptors, then Smad2 and Smad3 are phosphorylated and form the dimer, the Smad4 binds to the dimer to produce a trimer-Smad2/3/4, and the complex of Smad2/3/4 is transferred into the nucleus to enhance the expression activity of target genes such as Oct4 and SSEA-1. FGFR, fibroblast growth factor receptor; SSEA-1, stage-specific embryonic antigen-1; TGFβ, transforming growth factor beta.
In chicken, bFGF displays the necessity for the proliferation of preblastodermal cells and the primordial germ cells (PGCs) in vitro (Miyahara et al., 2016). Compared to be treated with hLIF or mSCF, the number and size of cESC colonies and the expression level of FGFRs in treatment group supplemented with hLIF, mSCF, and hbFGF were significantly increased. When the FGFR was inhibited by PD173074, cESCs were partially differentiated, which suggested that cESCs were prone to differentiate when bFGF was blocked (Zhang et al., 2018).
Transforming growth factor beta family
The transforming growth factor beta (TGFβ) super family is composed of three subgroups of conserved proteins, including TGFβs, bone morphogenic proteins (BMPs), and Activin. They exert evolutionally fundamental functions in a wide variety of important biological processes such as morphogenesis, cell fate determination, proliferation, differentiation, and apoptosis by regulating the TGFβ/Activin/Nodal pathway and BMP/growth and differentiation factor/Muellerian inhibiting substance pathways (Abou-Ezzi et al., 2019).
TGFβ or Activin protein activates the TGFβ/Smad signaling pathway by binding to the TGFβR/Alk heterodimer receptor (Schmierer and Hill, 2007). Then, the serine/threonine kinase in the intracellular domain of type I receptor (Alk1) phosphorylates the TGFβ signaling mediators—Smad proteins (Samd1/2/3/5/8). When this pathway is activated, Smad2/3 forms dimer that conjugates with Smad4 to generate a Smad2/3/4 trimer, and then the trimer is translocated into the nucleus and binds to the proximal promoter region of Nanog, directly activating the expression of Nanog in ESCs (Brown et al., 2011). In turn, the expression of Nanog blocks the endodermal differentiation induced by activating TGFβ/Smad pathway and enhances the maintenance of pluripotency (Bertero et al., 2015).
The TGFβ/Smad signaling pathway plays a central role in promoting the self-renewal of epiblast stem cells and human ESCs, while treatment with TGFβ receptor inhibitor (SB431542) seriously impairs the self-renewal of ESCs in mice (Madhu et al., 2016). After the inhibitory Smad family members, including Smad6 and Smad7, were transfected into ESCs to antagonize the BMP signaling pathway, a phenomenon of generating fewer and smaller colonies of cESCs was observed (Fang et al., 2016; Xu et al., 2008).
Although all members of the TGFβ super family regulate downstream reactions mainly through Smad proteins, there are some differences in the types of these Smad proteins (Mullen et al., 2011). For example, Activin can activate the Smad2/3/4 in a similar way, which is initiated by binding of ligand to the type I transmembrane receptor on the surface of cells, while the mediators of phosphorylated BMP are the Smad1/5/8 (Fig. 5) (Vallier et al., 2009). Smad1, Smad5, and Smad8 undergo homo-trimerization and form heteromeric complexes, after being combined with Smad4; these complexes will be translocated into the nucleus to regulate the transcription of targeted genes.

Mechanism on regulating the pluripotent maintenance of cESCs through BMP/Smad signaling pathway and LIF/STAT3 signaling pathway. BMP leads to homo-trimerization of Smad1/5/8 and subsequent association with co-regulatory component Smad4 to activate the expression of Id1/2/3 and DUSP9, which inhibits neuroectoderm differentiation. In parallel, the capacity of STAT3 to induce self-renewal of cESCs is enhanced by Smad1/5/8, which probably involves direct and indirect mechanisms. Therefore, the withdrawal of LIF results in a switch of BMP action from supporting self-renewal of cells to promoting lineage commitment. BMP, bone morphogenic protein; DUSP9, dual-specificity phosphatase 9; Id, inhibitor of differentiation.
BMP could induce the expression of inhibitor of differentiation genes (Id genes) and dual-specificity phosphatase 9 (DUSP9) with the synergistic action of LIF using the BMP/Smad pathway, and the overexpression of Id gene could liberate ESCs from BMP- or serum-dependence; even the self-renewal of cESCs could be maintained in the culture medium supplemented with only LIF (Hayashi et al., 2016). Forced expression of DUSP9 elevated the effects of BMP on the repression of early neural differentiation, which could substitute BMP4 to promote ESC self-renewal. Therefore, DUSP9 may strengthen BMP4 signaling by attenuating ERK activity and solidifying the self-renewal status of mESCs under the joint action of LIF (Li et al., 2012).
Junb and Socs3, two known targets for LIF, appear to be more highly induced by LIF in the presence of BMP. However, neither Junb nor Socs3 is candidate of effector for self-renewal of ESCs. Junb– ESCs show no defects, and Socs3 acts as a negative feedback regulator of gp30 (Schmitz et al., 2000). BMP could exacerbate the perception of LIF to STAT3, enhance the cascade reactions of LIF/STAT3, and block the expression of proteins which are responsible for the differentiation of ESCs to neural cells (Antebi et al., 2017; Ying et al., 2003). These results suggest that the suppression of differentiation and concomitant self-renewal of ESCs require an additional signal provided by BMP.
Serum
Serum such as fetal bovine serum (FBS) is the natural medium, which contains abundant nutrients for supporting the growth and proliferation of cells and for keeping a stable state during their culture (Guo et al., 2016). Moreover, it is the source of factors needed for cells to adhere to and spread on the plastic culture matrix (Grün et al., 2014). The heterogeneity of gene expression in culture medium has been noticed. For example, T-box3, a highly variable pluripotent marker, exhibited a distinct bimodal distribution in serum-cultured ESCs, which suggested that the serum-based culture condition for ESCs might be a major contributor to variable gene expression observed in single-cell analysis (Kanatsu-Shinohara et al., 2014).
The problems raised from the use of serum in cell culture, such as the addition of uncertain proteins, the varying quality and composition of different batches, and the risk of contamination with mycoplasma, viruses, bovine spongiform encephalopathy agent, and so on, have led to the development of a variety of serum free media (Ma et al., 2018).
As an alternative protein supplement, the knockout serum replacement (KSR) was originally developed by Life Technologies for culturing ESCs and had been successfully used in a variety of mammal cells to compensate the functions of the serum (Desai et al., 2015). Albumax is a component of KSR; spermatogonial stem cells (SSCs) could be maintained for at least 5 months in a serum-free Iscove's modified Eagle's medium supplemented with fetuin and KSR (Garcia-Gonzalo and Juan-Carlos, 2008). The cESC-like colonies could be generated in KSR medium added with R2i (MEK inhibitors D0325901 and TGFβ inhibitor SB431542) and LIF (Farzaneh et al., 2018). KAv-1 medium supplemented with KSR and R2i was also useful for the proliferation and longtime maintenance of the undifferentiated state of chicken PGCs (Naito et al., 2015).
Feeder layer
The plastic culture dishes coated with feeder layers are mainly developed to culture cESCs. The feeder layers are prepared from various types of somatic cells such as mouse embryonic fibroblasts (MEFs) and chicken embryonic fibroblasts (CEFs) after being treated with mitomycin C or γ ray to inhibit their proliferation; these cells can secrete paracrine factors, including LIF and bFGF, which inhibit the differentiation of ESCs and increase their self-renewal (Azizi et al., 2019). The establishment of cESCs was successfully realized using a Sandos inbred mouse (SIM)-derived 6-thioguanine- and ouabain-resistant (STO) cell line feeder cells in a FBS-containing culture medium, but high-density STO cells would impede the growth of cESCs (Lavial et al., 2008).
MEF and CEF feeder layers were efficient for the derivation of ESC-like cells and embryonic germ cell-like cells in duck; however, the pluripotent features of these cells were difficult to be maintained after the first passage (Guan et al., 2010). The CEF feeder cells could support the growth of cESCs, but unfortunately failed to maintain the pluripotency of them (Miyahara et al., 2016). Meaningfully, cESCs cultured on DF-1 feeder layer in the basic culture medium supplemented with hbFGF, mSCF, and hLIF showed typical ESC-like morphology, expressed primitive stem cell markers, and maintained a relatively stable proliferation rate and high telomerase activity in vitro for a long term (Zhang et al., 2018). These studies mentioned above indicate that the feeder layer is conducive for the pluripotency maintenance of cESCs in vitro.
In addition to the feeder layers, gelatin, matrigel, and collagen are commonly used to improve the adhesion, migration, and proliferation of ESCs. The effects of different feeder layers and various feeder layer-free extracellular matrix (ECM) on the derivation and maintenance of cESCs were compared, and a relatively typical morphology of ESCs was found on MEF feeder layer, but these were not occurred on human foreskin fibroblast cells, CEFs, Buffalo rat liver-3A cell line (BRL 3A) cells, Matrigel, Rogel (ECM secreted by MEFs), and agarose. None of these ECMs, including Matrigel, Rogel, agarose, and gelatin, could support the self-renewal of cESCs. On the contrary, they potentiated the differentiation of cESCs into three germ layers (Farzaneh et al., 2018).
ECM offered a support for cell adhesion through the integrin receptor-delivered extracellular signals; these signals could regulate the proliferation, migration, and differentiation of mouse SSCs by activating the expression of integrin α6 and integrin β1 (Rastegar et al., 2013).
However, the traditional feeder layers or ECMs for culturing cESCs always bring a series of unintended outcomes in morphologic change, geometrical alteration, abnormal expression of several abnormal genes and proteins, etc (Lavial and Pain, 2010). In most current culture systems, cESCs are exposed to animal feeder layers and xenogeneic reagents such as serum, which raises the safety concern in clinical applications (Mallon et al., 2006). Therefore, the defined culture systems for cESCs without both serum and feeder layer are the trend of future development.
Applications of cESCs
Production of recombinant therapeutic protein and monoclonal antibody
The great demand for proteins in therapy of diseases (such as cancer and metabolic disorders) and biological researches increases the focus on finding the appropriate hosts for the mass production of high_activity recombinant proteins within a short generation time. Various transgenic animal species have the capability to produce recombinant proteins in their milk or eggs. Especially transgenic chicken has been of interest for this purpose due to their short breeding time, simple management, and high productivity (Fig. 6) (Houdebine, 2008; Wang et al., 2017b). Furthermore, the human monoclonal antibodies and therapeutic proteins isolated from egg white have almost correct post-translational modification, making them become an effective resource for developing therapeutic drugs (Kojima et al., 2014).
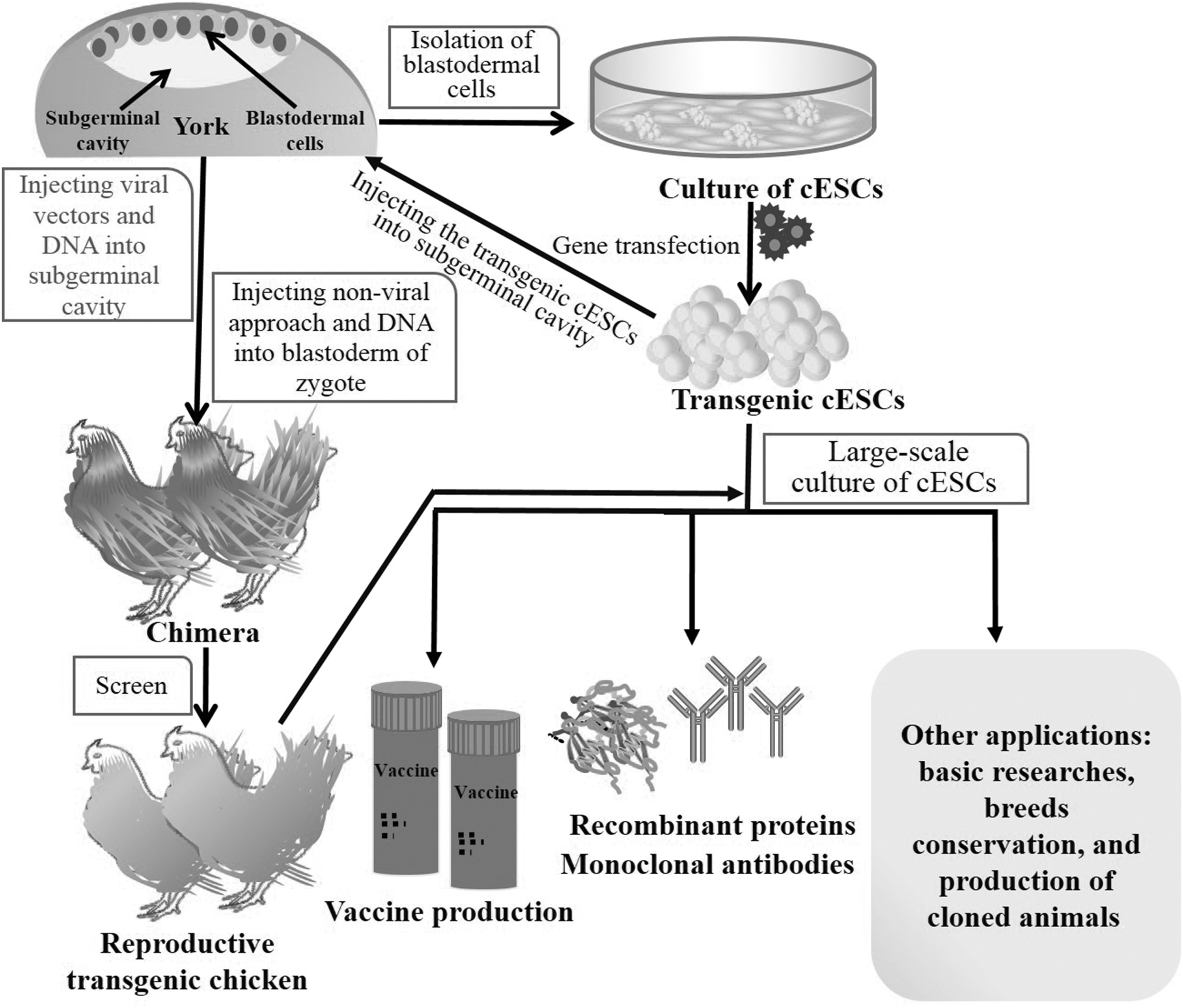
Schematic diagram of applications of cESCs.
There are about 4 g proteins in one egg white, more than 50% of them come from the expression of ovalbumin (OV) gene. If an exogenous DNA is inserted into the OV locus, it will be expressed under the control of the OV promoter, producing large quantities of recombinant proteins without any change in functionality. A variety of human recombinant proteins (such as interferon and erythropoietin) and transgenic avian protein (e.g., epidermal growth factor) were successfully produced in the egg white of transgenic bird (Kwon et al., 2018). Commendable and relatively simple protein components in egg white and the mature methods for the separation and purification of these proteins make the egg white become an ideal source for producing recombinant proteins.
Moreover, like the human oligosaccharides, the oligosaccharide residues of polypeptides in egg whites can be decomposed in a similar manner, which may contribute to overcome the allergic reactions when these proteins are used in clinical applications.
Vaccine production
It is more important to find an ideal model to overcome the weakness such as low efficiency, high cost, and low titer during the production of the vaccines. The focus of attention has shifted to the generation of cell lines from the early stage of avian embryo (Fig. 6) (Farzaneh et al., 2017; Giotis et al., 2019). Avian-derived cell lines are an alternative for virus production, because they provide a simple flexible system that can be controlled for an extended time with the capability of rapid vaccine production, especially during the outbreak of pandemic. cESCs have been evaluated as hosts for viruses that maintain genomic stability after repeated cell passages.
Until now, more than 20 different families of viruses such as influenza, poxvirus, parainfluenza, and measles have been efficiently propagated in embryonic 66 (EB66) cells derived from cESCs; over 60 human and veterinary EB66-based vaccines were developed (Perugi et al., 2019).
Avian iPSCs generated from avian embryonic fibroblasts through being transfected with transcription factors such as OCT4, NANOG, SOX2, KLF4, LIN28, and c-MYC may also be an excellent candidate for producing viral vaccines (Lu et al., 2012a). Although avian iPSCs have multiple advantages and applications, numerous challenges should be eliminated before they can be used in the production of vaccine. For example, these cells should be tested to eliminate the existence of endogenous virus; the presence of them in the host would result in allergic reactions after inoculation (Shittu et al., 2016).
Preparation of transgenic chicken
The great application potential of transgenic birds in husbandry and researches has attracted extensive attention; the genetic manipulation of birds offers insights into early embryonic development, cell tracking, species hybridization, and conservation of endangered species (Fig. 6) (Kagami, 2016; Yu et al., 2014). So far, two major strategies have been tried to produce transgenic chicken. One involves injecting the viral vectors and DNA directly into the subgerminal cavity of the recipient embryo at stage X, then these exogenous genes were transfected into the BCs to get chimera. The germline transmission of those transgenes in transgenic chicken has not yet been successfully achieved mainly because of the limitation of DNA fragment size (Collarini et al., 2019).
Another method is related to nonviral approach for carrying foreign gene with a large size, and the more striking difference is that the exogenous genes are not injected into the subgerminal cavity but the blastoderm of zygote; however, the method is not easy to access, there are many difficulties in determining the exact location of the blastoderm (Bednarczyk et al., 2018).
A new and more efficient transgenic strategy is based on cell-mediated target gene injection through cESCs and PGCs in vitro. After being transfected, transgenic cells are injected with microtubule needle into the subgerminal cavity or the dorsal aorta of recipient embryos at stage 14. The PGCs may migrate into the gonads, where they contribute to the generation of transgenic germ cells (Collarini et al., 2019). The male and female chimeras have been generated by injecting the donor cESCs derived from one blastoderm into recipients (Lu et al., 2012b). But the transgene efficiency is not up to the expectation, and those other strategies need further optimization.
Others
cESCs can be used not only in the preparation of medicinal proteins, monoclonal antibodies, vaccines, and transgenic chickens but also in basic researches, conservation of breeds, and production of cloned animals. As the most primitive ESCs in domestic birds, cESCs have the potential to differentiate into a variety of cells and can be induced to reproduce the relevant stages of normal early embryonic development, constructing the ideal experimental models for studying avian developmental biology (Fig. 6) (Lin et al., 2015).
In addition, cESCs can be applied to explore the mechanisms on cell differentiation and gene regulation of development, which undoubtedly provides a broader idea for the applications of human ESCs, because there is high resemblance of genes between poultry and mammals. cESCs cryopreserved in vitro can furnish a reliable means for protecting the rare or endangered breed. As the nuclear donors, cESCs can also be used to produce cloned animals to obtain a large number of excellent individuals in a short time and maximize the production efficiency of improved species.
Conclusion
cESC can be obtained from blastoderm and maintained in vitro. They have the potential to form EBs and differentiate into derivatives of all three embryonic germ layers in teratoma. However, the existing challenges in the isolation and culture of cESCs still impede their long-term maintenance of pluripotency in vitro; developing the isolation strategies and improving the culture system based on further understanding about action mechanisms of growth factors and cytokines will contribute to the establishment of cESC line. Serum-free and feeder layer-free culture system of cESCs is preferable for reducing or eliminating the undefined factors and improving their safety. We are indeed convinced that cESCs represent a unique model to study and bring a better understanding of stem cell physiology and offer an important cell resource for genetic, developmental, and vaccine manufacture studies.
Footnotes
Author Disclosure Statement
The authors declare they have no competing financial interests.
Funding Information
This work was supported by Chongqing Technological Innovation and Application Demonstration for Social and Livelihood Development (cstc2018jscx-msybX0240), the National Natural Science Foundation of China (Grant No. 31572488), and the Based and Advanced Research Projects of Chongqing (Grant No. cstc2017jcyjAX0477).