
Abstract
Pig cloning technique is valuable in agriculture, biomedicine, and life sciences. However, the full-term developmental efficiency of cloned pig embryos is only about 1%, which limits pig cloning application. The quality of recipient oocytes greatly affects the developmental competence of cloned pig embryos. Thus, this study investigated the effects of a recipient oocyte source (in vivo matured [IVVM] oocytes vs. slaughter house-derived in vitro matured [IVTM] oocytes), and follicular liquid treatment (slaughter house-derived immature follicle-derived fluid [IFF] vs. in vivo-matured follicle-derived fluid [MFF]) during the in vitro maturation (IVM) of oocytes on the development of the cloned pig embryos. Our results showed that using IVVM oocytes to replace IVTM oocytes as recipient oocytes, and using 10% MFF IVM medium to replace 10% IFF IVM medium could enhance the development of the cloned pig embryos. IFF and MFF contained different levels of oocyte quality-related proteins, resulting in different oocyte quality-related gene expression levels and reactive oxygen species levels between the 10% MFF medium-cultured oocytes and 10% IFF medium-cultured oocytes. This study provided useful information for enhancing the pig cloning efficiency by improving the quality of recipient oocytes.
Introduction
Pig somatic cell nuclear transfer (SCNT) or cloning has valuable applications in agriculture (Prather et al., 2003; Wang et al., 2017), biomedicine (Yan et al., 2018; Yifan et al., 2002), and life sciences (Jin-Ki et al., 2006; Niemann and Lucas-Hahn, 2012). However, the success rate of this technique is low. Only ∼1% of SCNT embryos transferred to surrogate sows can develop to birth (Ian et al., 2015; Liu et al., 2015; Nánássy et al., 2008), and merely 30%–50% (Ao et al., 2017; Kurome et al., 2013; Schmidt et al., 2014) of the born cloned piglets can survive into adulthood.
The extremely low efficiency of pig cloning is caused by the erroneous reprogramming of somatic nuclei in reconstructed embryos (Niemann et al., 2008; Niemann, 2016; Shuang et al., 2015). The induction of somatic cell reprogramming in SCNT embryos is mainly determined by enucleated recipient oocytes. Thus, the quality of recipient oocytes should influence the development of SCNT embryos.
The quality of in vivo matured (IVVM) oocytes is higher than that of slaughter house-derived in vitro matured (IVTM) oocytes (An et al., 2019; Egashira et al., 2019; Nakamura et al., 2017). In cattle (Akagi et al., 2008), goat (Ohkoshi et al., 2015), and sheep (Wells et al., 1997) SCNT, the developmental competence of IVVM oocyte-produced cloned embryos is higher than that of IVTM oocyte-generated counterparts. In pig SCNT, IVTM oocytes are commonly used as recipient oocytes because using IVVM oocytes is expensive, and many low-cost porcine oocytes are available in slaughter houses. Nevertheless, whether the use of IVVM oocytes to replace IVTM oocytes as recipient oocytes can enhance the development of cloned pig embryos is unknown.
The supplementation of porcine follicle fluid (FF) to an in vitro maturation (IVM) medium can improve the quality of IVTM pig oocytes (Vatzias and Hagen, 1999; Yvonne et al., 2013; Zhang et al., 2017a), and 10% (v/v) of slaughter house-derived immature follicle-derived fluid (IFF) is usually added to an IVM medium of recipient oocytes for pig cloning (Jeon et al., 2016; Kwak et al., 2012; Mordhorst et al., 2019). However, whether the use of in vivo-matured follicle-derived fluid (MFF) to replace the IFF in an IVM medium can enhance the quality of IVTM recipient oocytes and the subsequent development of pig SCNT embryos is unknown.
This study aimed to investigate the effects of recipient oocyte sources (IVTM vs. IVVM) and FF treatments (IFF vs. MFF) during recipient oocyte IVM on the development of cloned pig embryos. The results indicated that the use of IVVM oocytes to replace IVTM oocytes as recipient oocytes and the use of MFF to replace the IFF in recipient oocyte IVM medium could enhance the development of cloned pig embryos.
Materials and Methods
Ethics statement
The animal experimental protocol was approved by the Institutional Animal Care and Use Committee of South China Agricultural University. All efforts were made to minimize animal suffering.
Chemicals
All the chemicals used in the experiments were procured from Sigma-Aldrich Company (MO) unless otherwise mentioned.
Preparation of IVM media
Three IVM media were prepared as follows: (1) standard medium, TCM-199 medium supplemented with 10% (v/v) fetal bovine serum (FBS; Gibco, Grand Isle, NY), 0.6 mM cysteine, 10 ng/mL epidermal growth factor, 10 IU/mL equine chorionic gonadotropin, and 10 IU/mL human chorionic gonadotropin; (2) 10% IFF medium, standard medium added with 10% IFF; and (3) 10% MFF medium, standard medium added with 10% MFF.
Preparation of IVTM oocytes
Porcine ovaries of 6- to 7-month-old gilts were obtained from a local slaughterhouse and transported in 0.9% (w/v) NaCl solution supplemented with penicillin-G (100 IU/mL) and streptomycin sulfate (100 mg/L) at 30°C–35°C to our laboratory within 3 hours. Follicular fluid with oocytes was aspirated from antral follicles (3–6 mm in diameter) by using an 18-gauge needle connected to a 10 mL disposable syringe and collected into a 50 mL centrifuge tube. Cumulus-oocyte complexes (COCs) were recovered under a stereomicroscope, and COCs with at least three layers of compact cumulus cells and a homogeneous cytoplasm were selected for IVM. The selected COCs were washed thrice in HEPES-buffered Tyrode's medium containing 0.05% (w/v) polyvinyl alcohol.
Approximately 50–60 oocytes were transferred to each well of a four-well Nunc dish with 500 μL of different IVM media and cultured at 38.5°C with 5% CO2 in a humidified atmosphere for 44 hours. The COCs were treated with DPBS (Gibco) containing 1 mg/mL hyaluronidase to remove the surrounding cumulus cells by pipetting. The oocytes were observed under a stereoscope, and those with the first polar body in the perivitelline space were considered mature. Mature oocytes with intact cell membranes and clear perivitelline spaces were selected for SCNT.
Preparation of IVVM oocytes
Sows in standing estrus on the fifth day after weaning were anesthetized on the seventh day after weaning. Their oviducts were exposed via midline abdominal surgery. IVVM oocytes were flushed out from the oviducts with DPBS, and those with intact cell membranes, clear perivitelline spaces, and extruded first polar bodies were selected for SCNT.
Collection of MFF and IFF
Sows in standing estrus on the fifth day after weaning were anesthetized on the sixth day after weaning. Their ovaries were exposed via midline abdominal surgery. The main ovarian vessels were ligated to avoid hemorrhage. MFF was extracted from large follicles that were about to rupture in ovaries that had at least one freshly ruptured follicle (Fig. 1) by using a 10 mL syringe with an 18-gauge needle and placed in a centrifugal tube for 10 minutes. The supernatant was centrifuged at 3000 rpm/min for 5 minutes, taken, filtered through a 0.22 μm Millex GV filter (Millipore, Bedford, MA), transferred to cryogenic vials, and frozen in liquid nitrogen. All operations were completed within 30 minutes from MFF extraction.
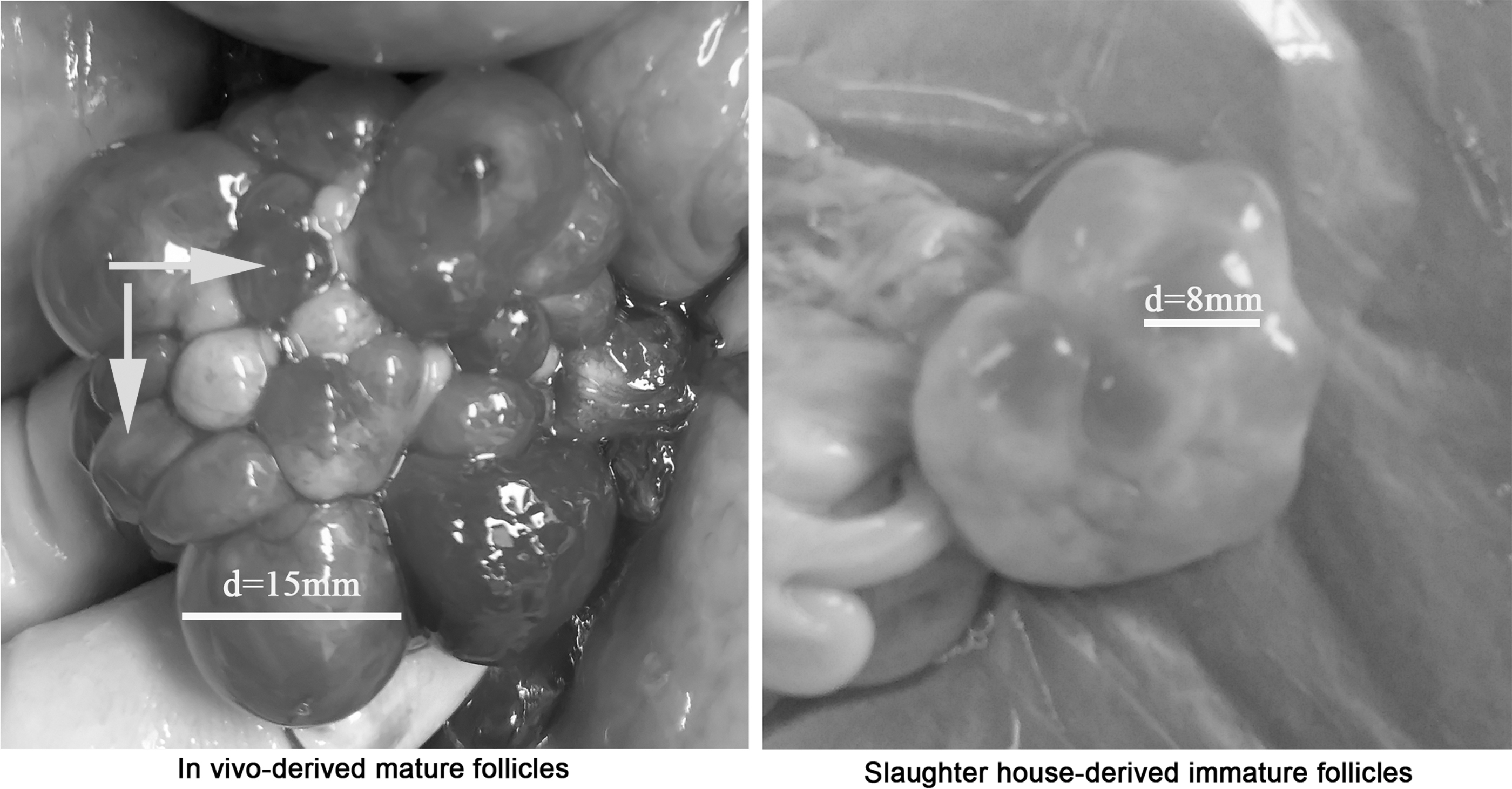
Mature follicles derived in vivo and immature follicles derived from a slaughter house. Arrow points to ovulated follicles. “d” represents the diameter of follicles.
IFF was collected from the antral follicles in immature ovaries derived from the slaughter house (Fig. 1) by using 10 mL syringes with 18-gauge needles. The supernatant was centrifuged, filtered, and stored as mentioned above.
Preparation of donor cells
Fibroblast cells were obtained via a ear biopsy from a Duroc boar. Ear tissues were quickly rinsed in 75% ethanol and DPBS (Invitrogen, Grand Island, NY) supplemented with penicillin (100 IU/mL) and streptomycin (100 μg/mL), minced into 1–2 mm pieces, and washed in Dulbecco's modified Eagle's medium (DMEM) with 10% FBS. Ear tissue fragments were settled at the bottom of 100 mm cell culture dishes and cultured in 5 mL of DMEM with 10% FBS. The dishes were then placed in an incubator humidified with 5% CO2 at 38.5°C, and the culture medium was changed every other day.
The fibroblasts were harvested with 0.25% trypsin-EDTA and passaged at 1:3 split ratios (passage number 1, P1). The fibroblast cells at passages 3–5 were frozen in liquid nitrogen with the medium containing 50% FBS, 40% DMEM, and 10% DMSO. Before SCNT, the cells were thawed and cultured in DMEM plus 10% FBS for 3–4 days until they reached 80%–90% confluence. Adherent cells (passages 6–10) were treated with trypsin for ∼1 minute and used for SCNT.
SCNT and in vitro culture of cloned embryos
Matured oocytes and a small number of donor cells were placed in a T2 medium (TCM-199 plus 2% FBS) containing 7.5 μg/mL cytochalasin B (CB). The oocytes were enucleated using a microinjection needle (Lingen Precision Medical Products Co., Ltd., Shanghai, China) with an inner diameter of 17 μm. The microinjection needle was inserted to aspirate the first polar body with about 15% of the adjacent cytoplasm, which contained the oocyte genomic material. Donor cells with a round and slightly burr-like shape were aspirated into the injection needle, and one donor cell was injected into the perivitelline space of each enucleated oocyte. The reconstructed oocytes were washed thrice in PZM-3 (porcine zygote medium-3) and then electrically fused with two direct current pulses of 150 V/mm for 50 ms in 0.28 mol/L mannitol supplemented with 0.1 mM MgSO4 and 0.01% PVA (polyvinyl alcohol).
The fused oocytes were cultured in a PZM-3 medium for 1 hour before electroactivation, cultured in a medium supplemented with 5 mg/mL CB, and incubated at 38.5°C in the presence of 5% CO2 and saturated humidity for 4 hours. The reconstructed oocytes were activated by two direct current pulses of 100 V/mm for 20 ms in 0.28 mol/L mannitol supplemented with 0.1 mmol/L MgSO4 and 0.05 mmol/L CaCl2. The reconstructed embryos were transferred to fresh PZM-3 and further cultured, and the cleavage and blastocyst rates were evaluated 2 and 6 days after activation. The total blastocyst cell count was performed with Hoechst 33342 staining under an epifluorescence microscope (Ti2; Nikon).
Parallel reaction monitoring analysis of FF protein levels
The highly abundant proteins of the follicular fluid were removed using a ProteoMiner™ protein enrichment kit (Bio-Rad). Then, the samples were lysed with a lysis buffer containing 100 mM NH4HCO3 (pH 8), 6 M urea, and 0.2% SDS and ultrasonicated in an ice bath for 5 minutes. The samples were completely mixed with four times the volume of precooled acetone by vortexing. The samples were centrifuged, and the precipitate was collected. The pellet was dissolved in a dissolution buffer, which contained 0.1 M triethylammonium bicarbonate (pH 8.5) and 6 M urea.
A BSA (albumin from bovine serum) standard protein solution was prepared with a Bradford protein quantitative kit in accordance with the manufacturer's instructions. A standard curve was drawn with the absorbance of the standard protein solution, and the protein concentration of the sample was calculated. Afterward, 120 μg of each protein sample was digested overnight. The eluents were combined and lyophilized after washing and eluting.
Each sample was mixed with the same amount of peptides. The mixture (1 μg) was analyzed with a “label-free” method by using an EASY-nLC™ 1200 UHPLC system coupled with a Q Exactive HF-X mass spectrometer (ThermoFisher Scientific, Bremen, Germany). The sample was injected into a home-made C18 Nano-Trap column (2 cm × 75 μm, 3 μm), and peptides were separated on a home-made analytical column (15 cm × 150 μm, 1.9 μm). The chromatographic separation conditions are listed in Table 1. The separated peptides were examined with Q Exactive HF-X, with an ion source of Nanospray Flex™ (ESI), a spray voltage of 2.4 kV, and an ion transport capillary temperature of 320°C. Full-scan MS spectra (m/z 300–1500) were obtained in an orbitrap with the resolution as 6000 (at m/z 200), and the automatic gain control (AGC) target was 3 × 106.
Liquid Chromatography Elution Gradient
The top 40 intense ions were isolated to increase energy collisional dissociation MS/MS fragmentation. In MS/MS, the resolution was 15,000 (at m/z 200), and the AGC target value was 5 × 104. Fragmentation was performed with a normalized collision energy (NCE) of 27% and a dynamic exclusion parameter of 20 s. The offline data were searched with PD2.2 software. The “missed cleavage” was set as 0, and 1–3 unique peptides were selected for each protein. For PRM (parallel reaction monitoring), the mixed peptides described above were analyzed with a “full scan” followed by a parallel reaction monitoring “PRM” pattern. The chromatographic separation and full scan conditions were the same as above. The PRM was set at a resolution of 30,000 (at 200 m/z) with an AGC target value of 5 × 104, a maximum ion injection time of 80 ms, and an NCE of 27%.
The same amount of the trypsin-treated peptide of each sample was taken and spiked with an equal amount of the labeled peptide DSPSAPVNVT “V” R (“V” for heavy isotope labeling) as an internal standard. The samples were examined with the “full scan” followed by the “PRM” pattern as described above. The off-line data were evaluated with Skyline, and the peak area was corrected using the internal standard peptide.
Real-time quantitative polymerase chain reaction analysis of gene expression levels
Total RNAs were extracted to analyze gene expression from 150 COCs by using a total RNA kit (Omega, Atlanta, GA), and the total RNA concentration was quantified using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Wilmington, DE). Only the RNA samples that did not show signs of degradation were used and stored at −80°C. Then, cDNA was synthesized from 1 μg of total RNA by using the PrimeScript RT Reagent Kit (TaKaRa, Otsu, Japan) in accordance with the manufacturer's instruction. cDNA was stored at −80°C until analysis through quantitative real-time polymerase chain reaction. Quantitative gene expression analysis was performed using the PowerUp™ SYBR Green Mix (Applied Biosystems, MA) in accordance with the manufacturer's protocol.
The primer sequences of all transcripts are presented in Table 2. The expression of each target gene was quantified relative to that of the internal control gene (GAPDH) by using the following equation: R = 2−[ΔCt sample − ΔCt control].
Primers Used for Real-Time Quantitative PCR
Determination of reactive oxygen species levels in oocytes
The reactive oxygen species (ROS) levels of the oocytes were determined with a ROS Assay Kit (Yesen, Shanghai, China). The oocytes were incubated in M199 medium containing 10 μM 2,7-dichlorodihydrofluorescein diacetate (D2DCFDA) at 38.5°C for 15 minutes (in the dark), washed in the M199 medium twice at room temperature, and transferred in 30 μL of DPBS droplets. Fluorescence was observed using an epifluorescence microscope (Ti2; Nikon). A ROS fluorescence intensity was analyzed with ImagePro Plus.
Statistical analysis
The data in Table 3 were analyzed with univariate ANOVA and Tukey's test by using SPSS 21.0 (SPSS, Inc., Chicago, IL). Other data were compared using GraphPad Prism 8.0.2 (GraphPad Software Inc., San Diego, CA). Statistical significance was considered at p <0.05. Data are reported as the mean ± standard error of the mean.
Effects of Recipient Oocyte Sources on the Developmental Competence of Porcine Somatic Cell Nuclear Transfer Embryos
Values in the same column labeled with different superscripts differ at p < 0.05.
IVTM, in vitro matured; IVVM, in vivo matured; SCNT, somatic cell nuclear transfer; SEM, standard error of the mean.
Results
Effects of recipient oocyte sources on the developmental competence of porcine SCNT embryos
The developmental competence between IVTM oocytes-generated and IVVM oocytes-produced pig SCNT embryos was compared. The results indicated that the blastocyst rate and the number of cells per blastocyst of IVVM oocytes-derived SCNT embryos were higher than those of IVTM oocytes-generated SCNT embryos (Table 3). These results suggested that the quality of porcine IVVM oocytes and their capacity to support the development of SCNT embryos were higher than those of IVTM oocytes. The differences in quality between IVVM and IVTM pig oocytes could be related to their differences in morphological characteristics.
In particular, the perivitelline space of IVVM oocytes was large, and the extruded first polar bodies were bright and clear. By contrast, the perivitelline space of IVTM oocytes was small, and the extruded first polar bodies were dark and indistinct (Fig. 2). The perivitelline space of the IVVM recipient oocyte-produced SCNT embryos was also larger than that of the IVTM recipient oocytes-generated counterparts (Fig. 2).
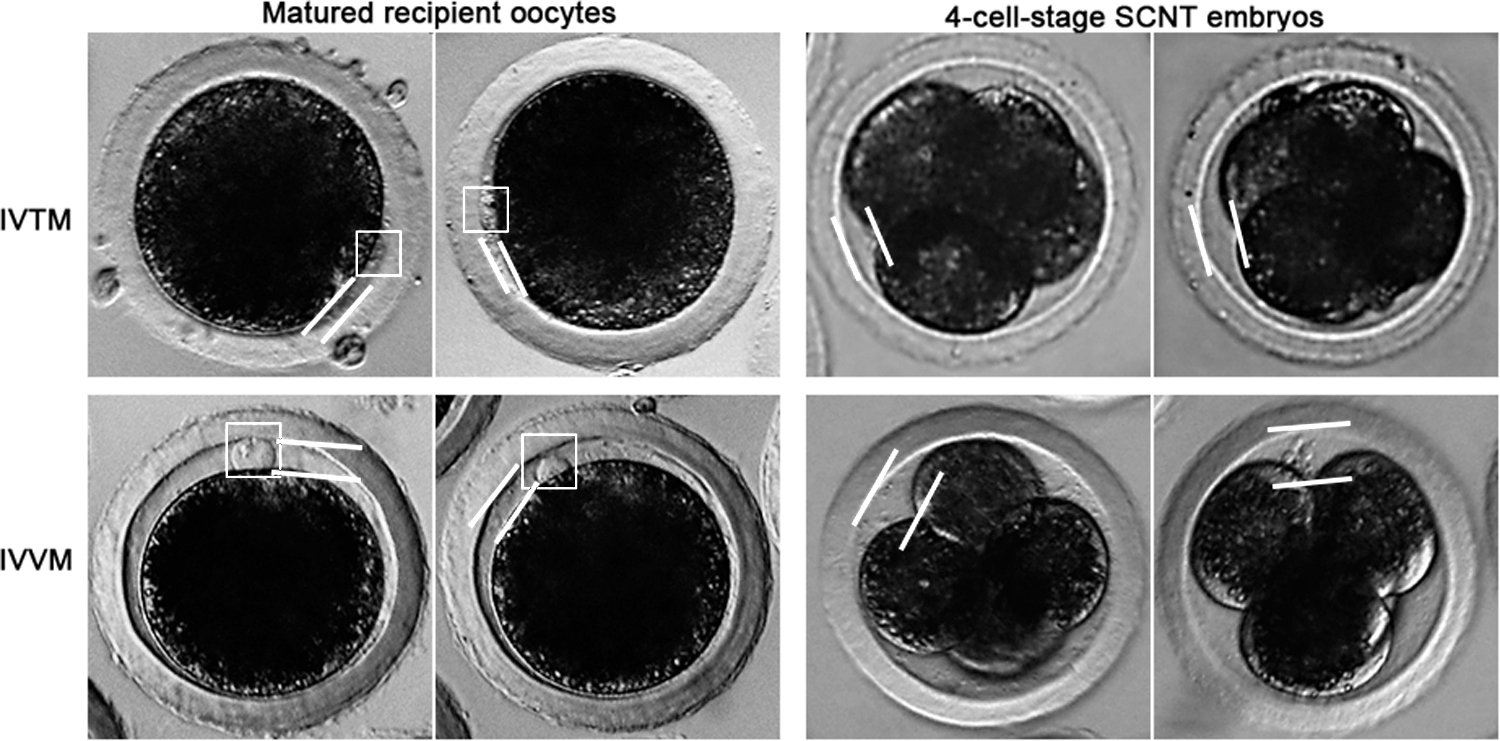
Morphological differences between IVTM and IVVM recipient oocytes and the SCNT embryos produced from them. The extruded first polar bodies (dark frame) in IVTM recipient oocytes are dark and indistinct, whereas they are bright and clear in IVVM recipient oocytes. The perivitelline space (i.e., the gap between two light lines) in IVTM recipient oocytes and IVTM recipient oocyte-generated SCNT embryos is smaller than that in IVVM recipient oocytes and IVVM recipient oocyte-generated SCNT embryos. IVVM, in vivo matured; IVTM, in vitro matured; SCNT, somatic cell nuclear transfer.
Effects of different FF treatments on the IVM of recipient oocytes and the developmental competence of SCNT embryos
The maturation rate was compared between the IVTM oocytes cultured with 10% IFF medium and 10% MFF medium for 44 hours during IVM. No difference in the maturation rate was found between the two groups of IVTM oocytes (Table 4). However, the blastocyst rate of the SCNT embryos derived from 10% MFF medium-cultured IVTM oocytes was higher than those derived from 10% IFF medium-cultured control IVTM oocytes (Table 4).
Effects of Different Follicle Fluid Treatments on the In Vitro Maturation of Recipient Oocytes and the Developmental Competence of Porcine Somatic Cell Nuclear Transfer Embryos
Values in a same column labeled with different superscripts differ at p < 0.05.
10% IFF-44 hours and 10% MFF-44 hours represent oocytes that were cultured with 10% IFF medium and 10% MFF medium, respectively, for 44 hours during IVM.
IFF, immature follicle-derived fluid; IVM, in vitro maturation; MFF, matured follicle-derived fluid.
Effects of double FF treatment on the IVM of recipient oocytes and the developmental competence of SCNT embryos
The maturation rate was compared among the IVTM oocytes cultured with 10% control IFF medium for 44 hours, 10% IFF medium for the first 24 hours and newly changed 10% IFF medium for the subsequent 20 hours (10% IFF-24 hours +10% IFF-20 hours), and 10% MFF medium for the first 24 hours and newly changed 10% MFF medium for the subsequent 20 hours (10% MFF-24 hours +10% MFF-20 hours). These three groups of oocytes did not differ in the maturation rate (Table 5). They also did not significantly vary in terms of the ability to promote the development of SCNT embryos, although the treatment with 10% IFF and 10% MFF twice during the IVM of recipient oocytes (the latter two groups) tended to increase the blastocyst rate of the SCNT embryos compared with the 10% control IFF-44 hours group (Table 5).
Effects of Twice Follicle Fluid Treatment on the In Vitro Maturation of Recipient Oocytes and the Developmental Competence of Porcine Somatic Cell Nuclear Transfer Embryos
10% IFF-44 hours represents oocytes that were cultured with 10% IFF medium for 44 hours during IVM.
10% IFF-24 hours +10% IFF-20 hours represents oocytes that were cultured with 10% IFF medium for the first 24 hours and then cultured with newly changed 10% IFF medium for the subsequent 20 hours during IVM.
10% MFF-24 hours +10% MFF-20 hours represents oocytes that were cultured with 10% MFF medium for the first 24 hours and then cultured with newly changed 10% MFF medium for the subsequent 20 hours during IVM. IVM, in vitro maturation.
Effects of the combined treatment with IFF and MFF on the IVM of recipient oocytes and the developmental competence of SCNT embryos
A combined treatment with IFF and MFF, namely, cultured with 10% IFF medium for the first 24 hours and 10% MFF medium for the subsequent 20 hours (10% IFF-24 hours +10% MFF-20 hours) was conducted during the IVM of the recipient oocytes. The resulting IVTM oocytes and the control IVTM oocytes cultured with 10% IFF medium for 44 hours showed no significant difference in the maturation rate and reprogramming capacity of the SCNT embryos, although the combined treatment with IFF and MFF during the IVM of recipient oocytes tended to improve the blastocyst rate of SCNT embryos compared with the control group (Table 6).
Effects of Combined Treatment of Immature Follicle-Derived Fluid and Matured Follicle-Derived Fluid on the In Vitro Maturation of Recipient Oocytes and the Developmental Competence of Porcine Somatic Cell Nuclear Transfer Embryos
10% IFF-44 hours represents oocytes that were cultured with 10% IFF medium for 44 hours during IVM.
10% IFF-24 hours +10% MFF-20 hours represents oocytes that were cultured with 10% IFF medium for the first 24 hours and then cultured with 10% MFF medium for the subsequent 20 hours during IVM.
Difference in the levels of the oocyte quality-related proteins between IFF and MFF
To investigate why the pig oocytes cultured with an MFF-containing IVM medium have a higher potential to support SCNT embryo development than the control oocytes cultured with an IFF-containing IVM medium (Table 4), we selected seven proteins participating in the regulation of oocyte quality and measured their levels in IFF and MFF. These seven oocyte quality-related FF proteins included heat shock protein 90 beta (HSP90β; Liu et al., 2018; Pires and Khole, 2009), follistatin (FST; Guo et al., 2018; Hussein et al., 2006; Lee et al., 2017), stromal cell-derived factor 1 (SDF1; Wang et al., 2018; Zhang et al., 2017b; Zuccarello et al., 2011), heat shock protein 70 (HSP70; El-Sayed et al., 2018; Souza-Cacares et al., 2019), insulin-like growth factor 1 (IGF1; Sato et al., 2017; Sirotkin et al., 2000), bone morphogenetic protein 15 (BMP15; Mariana Fernandes et al., 2015; Richani and Gilchrist, 2017), and growth differentiation factor 9 (GDF9; Jackowska et al., 2013; Monte et al., 2019).
Only the first three proteins were detected via the PRM method in IFF and MFF (Fig. 3). The HSP90β protein level of IFF was higher than that of MFF, although the difference in the HSP90β protein level between IFF and MFF was not statistically significant (Fig. 3). The FST of IFF was higher than that of MFF, but the SDF1 protein level of the former was lower than that of MFF (Fig. 3). These results indicated that the levels of some oocyte quality regulation proteins were different between IFF and MFF.

Relative levels of oocyte quality-related proteins in IFF and MFF. ND means nondetectable. Values are shown as mean ± SEM. Values are presented as mean ± SEM (***p < 0.001). IFF, immature follicle-derived fluid; MFF, matured follicle-derived fluid.
Difference in the mRNA levels of oocyte quality-related genes between 10% IFF medium-cultured IVTM oocytes and 10% MFF medium-cultured IVTM oocytes
To further investigate why the 10% IFF medium-cultured and 10% MFF medium-cultured IVTM oocytes had different capacities to promote SCNT embryo development (Table 2), we selected six genes involved in the regulation of oocyte quality and measured their mRNA abundance in two groups of oocytes. These six oocyte quality-related genes included tumor protein p53 (TP53), caspase 3 (CASP3) (Arias-Álvarez et al., 2017; Li et al., 2000; Zheng et al., 2005), GDF9, BMP15 (Heydarnejad et al., 2019; Zhang et al., 2005), C-X-C motif chemokine receptor 4 (CXCR4), and C-X-C motif chemokine receptor 7 (CXCR7) (Mcginn et al., 2012; Philipp et al., 2014; Sathish and Macoska, 2012).
The transcript levels of TP53 were lower in the oocytes cultured with 10% MFF for 24 and 44 hours than those in the oocytes cultured with 10% IFF for 24 and 44 hours, but the transcript levels of CASP3, GDF9, BMP15, CXCR4, and CXCR7 were higher in the former than those in the latter (Fig. 4). These results demonstrated that the expression levels of the tested oocyte quality-related genes were different in the oocytes cultured with 10% IFF medium and 10% MFF medium.

Relative mRNA levels of oocyte quality-related genes in oocytes cultured with 10% IFF medium for 24 and 44 hours, and oocytes cultured with 10% MFF medium for 24 and 44 hours. Values are shown as mean ± SEM. Values are presented as mean ± SEM (*p < 0.05).
Difference in ROS levels between 10% IFF medium-cultured IVTM oocytes and 10% MFF medium-cultured IVTM oocytes
ROS-induced oxidative stress is the main cause of poor oocyte quality (Agarwal et al., 2006; Liang et al., 2017; Mishra et al., 2016). To explore whether ROS level is related to the difference in the quality of 10% IFF medium-cultured oocytes and 10% MFF medium-cultured oocytes, we measured the ROS level in these two groups of oocytes. The results indicated that the oocytes cultured with 10% IFF medium for 24 and 44 hours contained a higher ROS level than the oocytes cultured with 10% MFF medium for 24 and 44 hours (Fig. 5). This result suggested that the supplementation of 10% MFF to the IVM medium decreased the oxidative stress in the IVTM oocytes compared with that of the addition of 10% IFF to the IVM medium.
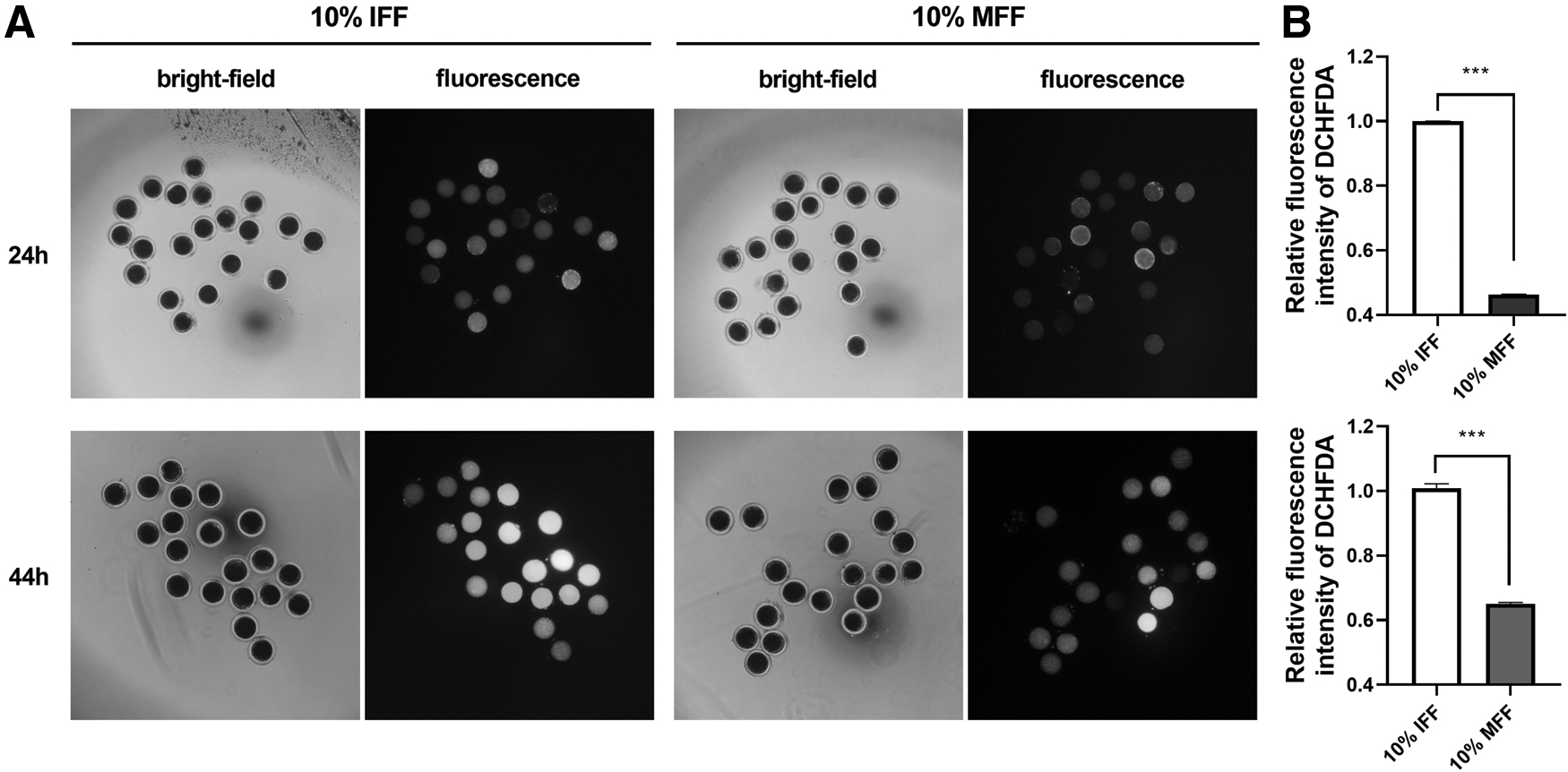
ROS level in oocytes cultured with 10% IFF medium for 24 and 44 hours, and oocytes cultured with 10% MFF medium for 24 and 44 hours.
Discussion
This study demonstrated that the ability of pig IVVM recipient oocytes to support the development of the cloned pig embryos was higher than that of IVTM recipient oocytes.
This result was consistent with findings in cattle (Akagi et al., 2008), sheep (Wells et al., 1997), and goat (Ohkoshi et al., 2015) cloning. Previous studies indicated that the developmental competence of IVVM oocyte-derived porcine in vitro fertilized pig embryos is higher than that of IVTM oocyte-derived counterparts (Egashira et al., 2019; Nakamura et al., 2017). The full-term developmental rate of IVVM oocyte-produced cloned pig embryos reaches 2.9% (Chen et al., 2013), which is about threefold of the average full-term developmental efficiency of cloned pig embryos (Liu et al., 2015). These studies have demonstrated that the quality of pig IVVM oocytes is higher than that of IVTM oocytes. Thus, the capacity of the former to support subsequent SCNT embryo development is higher.
Therefore, using IVVM oocytes to replace the commonly used IVTM oocytes as recipient oocytes for SCNT can improve the pig cloning efficiency. However, the cost of using IVVM oocytes for pig cloning is much higher than that of using IVTM oocytes. The reason is that pigs are polytocous animals, and a large number of cloned embryos are usually needed to be generated and transferred into surrogate sows to establish and maintain their pregnancies (Liu et al., 2015). Hence, a large number of recipient oocytes are required for cloned pig production. Obtaining a large number of IVVM oocytes from female pigs is not only costly but also technically difficult compared with that of obtaining a large number of IVTM oocytes from slaughter houses.
A feasible method for increasing pig cloning efficiency aims to improve the quality of IVTM oocytes through the supplementation of oocyte quality-enhancing molecules during IVM (Lee et al., 2016; Yuan et al., 2017). In the present study, we showed that using 10% MFF medium for the IVM of pig oocytes can improve their reprogramming capacity for SCNT embryos, suggesting that MFF contains higher levels of oocyte quality-enhancing molecules or lower levels of oocyte quality-decreasing molecules than IFF. This finding was confirmed by some studies (Oberlender et al., 2013; Wu et al., 2007).
Our results (Fig. 3) revealed that MFF had a higher SDF1, but a lower FST protein level than IFF. SDF1 is a positive regulator of folliculogenesis, oocyte maturation, oocyte quality, and oocyte developmental potential (Nishigaki et al., 2010; Wang et al., 2018). By contrast, FST negatively affects oocyte maturation and developmental competence (Guo et al., 2018).
The difference in protein composition between MFF and IFF resulted in lower TP53 expression and ROS levels, but higher CASP3, GDF9, BMP15, CXCR4, and CXCR7 expression levels in 10% MFF medium-cultured oocytes than 10% IFF medium-cultured oocytes (Figs. 4 and 5). TP53 and ROS are negative regulators of oocyte quality because TP53 can promote apoptosis and ROS can induce oxidative stress (Arias-Álvarez et al., 2017; Arias-Alvarez et al., 2018). GDF9 and BMP15 are oocyte quality-enhancing factors (Gilchrist et al., 2008; Jeong et al. 2019). CXCR4 and CXCR7 are SDF1 receptors, whose expression levels increase in SDF1-treated IVTM oocytes with an enhanced developmental ability (Zhang et al., 2017b).
The higher SDF1 content in MFF than in IFF resulted in a lower ROS level in 10% MFF-treated IVTM oocytes than 10% IFF-treated IVTM oocytes because SDF1 can protect cells from oxidative stress damage (Feng et al., 2017). The reduction of ROS levels can promote the expression of oocyte quality-enhancing GDF9 and BMP15 in 10% MFF-treated oocytes as previously demonstrated (Zhao et al., 2015). The lower ROS level might decrease the apoptosis rate in 10% MFF-treated COCs because the proapoptotic gene TP53 was downregulated in 10% MFF-treated COCs compared with those of 10% IFF-treated COCs.
However, the expression of another proapoptotic gene, namely, CASP3, increased in 10% MFF-treated COCs. This finding could be related to the reversion of cumulus cells to mural granulosa cells (Chaffin et al., 2012), which probably involve CASP3 regulation (Fernando and Megeney, 2007). The downregulation of TP53 and ROS and the upregulation of CASP3, GDF9, BMP15, CXCR4, and CXCR7 in 10% MFF medium-cultured oocytes explain why this group of oocytes has higher abilities to support the subsequent SCNT embryo development than 10% IFF medium-cultured oocytes. However, we could not rule out the possibility that other differential molecules between MFF and IFF, as identified in some studies (Algriany et al., 2004; Ito et al., 2008; Nandi et al., 2007), contributed to the differences in gene expression pattern and quality between MFF- and IFF-treated pig oocytes.
Although the oocytes in the 10% MFF-44 hours group exhibited a higher reprogramming capacity for subsequent SCNT embryo development than the control oocytes of the 10% IFF-44 hours group, the oocytes of the 10% MFF-24 hours +10% MFF-20 hours group and 10% IFF-24 hours +10% MFF-20 hours group showed a similar reprogramming ability for subsequent SCNT embryo development to the control oocytes of the 10% IFF-44 hours group (Tables 4–6). The treatment of 10% MFF-24 hours +10% MFF-20 hours might provide excess, whereas the treatment of 10% IFF-24 hours +10% MFF-20 hours might supply insufficient oocyte quality-enhancing molecules for the cultured oocytes compared with those of the treatment of 10% MFF-44 hours. These results suggested that oocyte quality was sensitive to the IVM culture medium.
In conclusion, the potential of IVVM recipient oocytes to promote subsequent pig SCNT embryo development was higher than that of IVTM recipient oocytes, and the reprogramming ability of the oocytes cultured with 10% MFF medium for 44 hours during IVM for subsequent pig SCNT embryo development was higher than that of the control oocytes cultured with 10% IFF medium for 44 hours during IVM.
Footnotes
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This study was supported by Guangdong Provincial Promotion Project on Preservation and Utilization of Local Livestock and Poultry Breeds, the National Natural Science Foundation of China (Grant number: 31772554), and the Department of Science and Technology of Guangdong Province, China (Grant numbers: 2018B030314004 and 2018B020203002).