
Abstract
Chimeric pigs harboring organs derived from human stem cells are promising for patient-specific regenerative therapies. Induced pluripotent stem cells (iPSCs) can contribute to all cell types of the fetus, including germline after injection into embryos. However, ethical concerns prohibit testing human iPSCs in chimera assays. Here, we evaluated porcine embryos as hosts for an interspecies chimera assay using iPSCs from either cynomolgus monkeys (cyiPSCs) or mouse (miPSCs). To establish an in vitro culture system compatible for cyiPSCs and porcine embryos, we determined blastocyst development in eight different stem cell media. The highest developmental rates of blastocysts were achieved in Knockout Dulbecco's modified Eagle's medium with 20% knockout serum replacement. We found that cyiPSCs injected into porcine embryos survived in vitro and were mostly located in the trophectoderm (TE). Instead, when miPSCs were injected into porcine embryos, the cells rapidly proliferated. The behavior of chimeras developed in vitro was recapitulated in vivo; cyiPSCs were observed in the TE, but not in the porcine epiblast. However, when miPSCs were injected into in vivo derived porcine embryos, mouse cells were found in both, the epiblast and TE. These results demonstrate that porcine embryos could be useful for evaluating the interspecies chimera-forming ability of iPSCs from different species.
Introduction
The successful introduction of defined transcription factors into somatic cells yielded induced pluripotent stem cells (iPSCs) that are similar to embryonic stem cells (ESCs) with regard to major pluripotent characteristics, including unlimited self-renewal in vitro and the potential to differentiate into all somatic cells, and germ cells, in vitro and in vivo (Choi et al., 2016; Takahashi et al., 2007). Thus, human induced pluripotent stem cells (hiPSCs) hold great promise for the generation of regenerative tissue or organs directly from the patient's somatic cells, thereby avoiding the problem of genetic incompatibility and immunological rejection (Takahashi et al., 2007). However, before application of human stem cells for clinical trials, safety and efficacy of iPSC-based therapies need to be analyzed thoroughly (Hong et al., 2016).
The most stringent criterion for validating the potency of stem cells is the generation of viable chimeras with germline transmission (Mascetti and Pedersen, 2016a). Chimeras can be produced either by combining stem cells or blastomeres with preimplantation embryos (Dvorak et al., 1995; Tarkowski, 1961), injection of stem cells into preimplantation embryos (Eckardt et al., 2011; Okita et al., 2007), or aggregation of stem cells with developmentally compromised tetraploid embryos (Nagy et al., 1990, 1993).
Due to ethical concerns with the use of human stem cells, only the ability to differentiate in vitro as embryoid bodies (EBs) or the teratoma assay were used for testing pluripotency (Hentze et al., 2009). The properties of hiPSCs have mainly been studied in mice and other small animal models (Azhdari et al., 2013; Hayashi et al., 2016; Yang et al., 2017a). However, any long-term engraftment of transplanted cells cannot be monitored using animals with such short lifespan like rodents.
Large animals such as the domestic pig offer major advantages over rodents due to their longer lifespan and high degree of similarity with human genetics, anatomy, and physiology (Rogers, 2016; Swindle et al., 2012; Ziegler et al., 2016). Furthermore, the production of interspecies chimeras between host embryos from large animals and human stem cells could provide a functional in vivo system for exploring human stem cell potency and efficacy of cell replacement therapies (Mascetti and Pedersen, 2016b; Wu et al., 2016).
Ultimately, pigs carrying organs that are entirely derived from human stem cells, or at least have a high contribution of human cells or tissues in the target organ, could be used as a new donor source for regenerative tissue or organ replacement for future treatment of patients. Therefore, we chose porcine preimplantation embryos as hosts in an in vitro and in vivo interspecies chimera assay to monitor allocation and developmental fate of the injected iPSCs.
However, the possibility to create human-animal chimeras raises novel biological and ethical questions (Hermerén, 2015). Therefore, experiments using iPSCs derived from non-human primates as donor cells for an interspecies chimera assay are required for exploiting long-term safety and potential organ integration with greater ethical acceptance (Wunderlich et al., 2014). It had been shown that established iPSCs from cynomolgus monkeys have a primed pluripotent state and are thus comparable in their characteristics with human pluripotent stem cells (Honda et al., 2017; Shimozawa et al., 2013; Wunderlich et al., 2012, 2014). The conversion of primed-state PSCs into a true naive state in cynomolgus monkey was unsuccessful (Honda et al., 2017).
To analyze the contribution of iPSCs from different species into viable porcine embryos and early conceptuses, we employed primed cynomolgus monkey iPSCs (cyiPSCs) and germline-competent naive mouse iPSCs (miPSCs) for this study. The potential of miPSCs and cyiPSCs for integrating into a chimeric organism after injection into porcine preimplantation embryos has not yet been investigated. As porcine preimplantation embryos and cynomolgus iPSCs possess different requirements for in vitro development, we first established culture conditions that were compatible with the in vitro development of chimeric embryos composed of porcine and cynomolgus cells. Subsequently, we analyzed the degree of chimerism after injecting miPSCs and cyiPSCs into early porcine embryos.
Results show that in contrast to cyiPSCs, miPSCs survived in both the epiblast and trophectoderm (TE) of porcine in vivo derived embryos. We observed that the in vitro interspecies chimera assay could recapitulate the behavior of injected iPSCs in in vivo chimeras.
Methods
Animal welfare
The pig experiments had been reviewed and were approved by the external ethics committee (Niedersaechsisches Landesamt fuer Verbraucherschutz und Lebensmittelsicherheit, LAVES, 33.14-42502-04-13/1298). The pigs were maintained and handled according to the German guidelines for animal welfare, and to the German law regarding genetically modified organisms.
Skin biopsies from cynomolgus monkeys (all 5 years of age) were obtained from CovanceR (Münster, Germany). The methods related to cyiPSC derivation were carried out in accordance with the relevant German guidelines and regulations. All experimental protocols related to cyiPSC derivation and culture had been approved by a supervisory committee of Hannover Medical School (MHH).
In vitro culture of cynomolgus monkey (Macaca fascicularis) iPSCs
The cyiPSCs were isolated as described previously (Wunderlich et al., 2012). Briefly, fibroblasts were isolated from skin biopsies from adult monkeys and transduced with the lentiviral vector plSh SSF. hOct34. hKlf4. hSox2. hMyc. i2dTomato. The transduced cells were cultured in fibroblast culture medium Dulbecco's modified Eagle's medium (DMEM), supplemented with 1 mM
Thereafter, cells were passaged onto irradiated murine embryonic fibroblasts (MEFs) and cultured in iPSC-20% medium containing Knockout DMEM, supplemented with 20% knockout serum replacement (KSR), 1 mM
Differentiation of cynomolgus monkey (M. fascicularis) iPSCs
CyiPSCs at passage 60 were detached from feeder cells using Collagenase IV (Gibco™), and seeded into ultra-low attachment plates (Corning®) for 7 days in differentiation medium (80% IMDM, supplemented with 20% FCS, 1% NEAA, 1 mM
For trophoblast differentiation cyiPSCs were plated on Geltrex-coated cell culture plates in medium precultured on irradiated MEFs (conditioned medium: DMEM/F12 supplemented with 15% KSR, 100 μM β-mercaptoethanol, 1% NEAA stock (all from Invitrogen), 100 ng/mL bFGF, and 10 mM Y-27632 (both from the Institute for Technical Chemistry, Leibniz University Hannover)). After 24 hours, the conditioned medium was replaced with differentiation medium (conditioned medium without bFGF and Y-27632, but containing 1 μM A8301 [Tocris], 0.1 μM PD173074 [Sigma-Aldrich], and 10 ng/mL BMP-4 [R&D Systems]). Forty-eight hours after addition of the differentiation medium, the cells were fixed with 4% paraformaldehyde and used for immunostaining analysis.
In vitro culture of miPSCs
An miPSC line was established from fetal fibroblasts isolated from Naval Medical Research Institute (NMRI) fetuses with the ubiquitous expression of the Venus fluorescence protein as described (Garrels et al., 2016). Briefly, fibroblasts were transfected with a piggyBac (PB) transposon construct, composed of the CAGGS promoter driven cassette containing the human reprogramming factors SOX2, OCT4, KLF4, and c-MYC, each separated by sequences coding for the self-cleaving 2A peptide, and flanked by PB-inverted terminal repeats. The complementary DNA (cDNA) coding for the hyperactive PB is driven by the CMV promoter on a helper plasmid (Talluri et al., 2014).
The miPSCs were cultured in vitro under standard conditions on 0.2% gelatin-coated plates in mouse ESC medium (mESC medium), containing DMEM High Glucose (Biowest), supplemented with 15% heat-inactivated FCS (Lot 42Q0154K; Gibco), 2 mM
The medium was changed every day and cells were passaged every 2–3 days. The emerging cells showed typical hallmarks of pluripotency, including expression of the endogenous pluripotency-related genes Oct4, Sox2, c-Myc, Klf4, Nanog, Rex1, and Utf1. Injection of miPSCs into immunodeficient mouse resulted in growth of teratomas containing cell types indicative of the three germ lineages. After injection into mouse embryos, only low contribution to chimeric mice was observed (Mall et al., 2017).
Differentiation of miPSCs
After dissociation with 0.05% trypsin-EDTA, miPSCs transgenic with Venus at passage 39 were counted and single drops containing 1000 cells/20 μL in mESC medium without LIF were plated on the internal face of the lid of ø 10 cm culture dish to form EBs for 3 days. Typical EBs were transferred to 0.2% (w/v) gelatin-coated dishes and cultured in ESC medium without LIF for additional 23 days. Venus fluorescence was evaluated on days 0, 1, 3, and 26, respectively, using fluorescence microscopy (DM IL LED; Leica) and flow cytometry (MACS Quant VYB; Miltenyi Biotec).
Karyotyping of iPSCs
For multicolor fluorescence in situ hybridization (mFISH), metaphase preparations from Venus transgenic miPSCs at passages 25 and 45 were prepared according to standard procedures (Schlegelberger et al., 1999). The analysis was carried out using a 21XMouse mFISH probe (Metasystems, Altlussheim, Germany). The mFISH analysis was performed according to the manufacturer's instructions. ISIS software was used for analysis (Metasystems). Whenever possible, five metaphases from each mouse were analyzed.
Adherent Venus cyiPSCs at passage 58 were treated for 30 minutes with colcemide (Invitrogen) at final concentrations of 0.025, 0.05, and 0.1 μg/mL, and were then dissociated with 0.83% trypsin/EDTA (Invitrogen). After hypotonic treatment with 60 or 75 mM potassium chloride (KCl) for 20 minutes at 37°C and cytogenetic cell preparation, fluorescence R-banding was performed according to standard procedures.
Production of parthenogenetic porcine embryos
Parthenogenetic embryos were produced as described previously (Hölker et al., 2005). Briefly, ovaries from prepubertal gilts were transported at 30°C from a local abattoir and washed three times with 0.9% sodium chloride (NaCl; Sigma-Aldrich), containing 0.06 mg/mL penicillin G potassium salt (AppliChem) and 0.131 mg/mL streptomycin sulfate (AppliChem). Oocytes were aspirated from follicles with a diameter of 2–6 mm using an 18-gauge needle and washed in Dulbecco's phosphate-buffered saline (DPBS) medium (AppliChem), supplemented with 0.33 mM sodium pyruvate (Sigma-Aldrich), 5.56 mM
Oocytes were matured in vitro in 1:1 DMEM high glucose (Biowest) and Ham's F-12 medium (Merck), supplemented with 60 μg/mL penicillin G potassium salt (AppliChem), 50 μg/mL streptomycin sulfate (AppliChem), 2.5 mM
Subsequently, cumulus cells were removed and oocytes with a visible polar body were exposed to a single pulse of 24 V for 45 μseconds in SOR activation medium (0.25 M sorbitol, 0.1 mM calcium acetate hydrate, 0.5 mM magnesium acetate tetrahydrate, and 0.1% bovine serum albumin [BSA; all from Sigma-Aldrich]), followed by 3 hours of incubation with 2 mM 6-dimethylaminopurine (DMAP; Sigma-Aldrich) in porcine zygote medium 3 (PZM-3 medium: 108 mM NaCl, 10 mM KCl, 0.35 mM potassium phosphate monobasic [KH2PO4], 0.40 mM magnesium sulfate heptahydrate [MgSO4 × 7 H2O], 25.07 mM sodium bicarbonate [NaHCO3], 0.2 mM sodium pyruvate, 5 mM hypotaurine, 20 μL/mL BME amino acid solution, 10 μL/mL MEM NEAA solution, 0.05 mg/mL gentamicin sulfate salt, 3 mg/mL BSA [all from Sigma-Aldrich], 2 mM
In vitro culture of porcine parthenogenetic embryos in different stem cell media
Porcine parthenogenetic embryos were cultured in PZM-3 medium for a total of 6 days; subsequently, they were randomly divided into nine groups, each containing 15–20 embryos. Blastocysts from day 6 were cultured in four-well culture dishes (Nunclon) filled with 500 μL culture medium, including (1) knockout DMEM with 10% (iPSC-10% medium) or (2) 20% (iPSC-20% medium) KSR, supplemented with 1 mM
Morphology, blastocyst cell numbers, and developmental rates (expanded, hatched, not-expanded, and degenerated blastocysts) were examined on days 7 and 8 and compared with embryos cultured in medium routinely used for porcine embryos, that is, PZM-3 medium (control).
Determination of blastocyst cell numbers
The total number of cells was counted in day 8 blastocysts after Hoechst 33342 staining (Sigma-Aldrich). Briefly, blastocysts were incubated in 500 μL of PZM-3 with 1 μg/mL Hoechst 33342 for 7 minutes at 39°C, 5% CO2, and 5% O2 and washed once in 500 μL PZM-3 to remove nonincorporated dye. Thereafter, blastocysts were transferred onto slides and covered with cover glass. The number of cells was counted on an average of 14–54 embryos in each group using a fluorescence microscope (Zeiss).
In vitro chimera assay
Porcine-monkey chimeric embryos
Two monkey iPSC lines, either transgenic for GFP or Venus, were used for embryo injection experiments. Cells at passage 56–68 were briefly treated with 0.1% collagenase type IV (Gibco), gently tapped out from monolayer and resuspended in iPSC-20% medium, and stored on ice.
For injection, cyiPSCs and porcine embryos were pipetted into 400 μL drops of injection medium containing a 3:1 mix of Opti-MEM® I (1 × ) + GlutaMAX™-I reduced serum medium (Gibco), supplemented with 10% FCS (Lot 42Q0154K; Gibco) and iPSC-20% medium. Cell clumps containing 4–6 cyiPSCs were injected into porcine parthenogenetic embryos from day 4 (8 cells—morulae) or day 6 (blastocysts) with the aid of a piezo-driven micromanipulator (Zeiss; Eppendorf). Injected embryos were washed twice and cultured in vitro either for 2 days in mixed medium (3:1) of PZM-3 and iPSC-20% medium (day 4 embryos) or in iPSC-20% medium (day 6 embryos) for 1 day at 39°C in 5% CO2 and 5% O2. Embryos were either plated onto MEFs or fixed and stored in DPBS (Sigma-Aldrich) supplemented with 0.5% FCS (Lot 42Q0154K; Gibco) and 1% penicillin/streptomycin solution (Corning) at 4°C for future analysis.
Porcine-mouse chimeric embryos
The miPSCs transgenic with Venus at passage 45 were injected into parthenogenetic porcine embryos. Cells growing on 0.2% gelatin-coated plates were detached with 0.05% trypsin-EDTA (GE Healthcare), washed once and resuspended in mESC medium, and stored on ice. During injection, both embryos and miPSCs were maintained in separate 400 μL drops of 3:1 mix of Opti-MEM I (1 × ) + GlutaMAX-I reduced serum medium (Gibco), supplemented with 10% FCS (Lot 42Q0154K; Gibco) and mESC medium. After injection of cell clumps (4–6 cells), presumptive chimeric embryos were washed twice and cultured in mixed medium (3:1) of PZM-3 and mESC medium for 2 days (day 4 embryos) or in mESC medium (day 6 embryos) for 1 day at 39°C in 5% CO2 and 5% O2.
Embryos were either plated onto MEFs or fixed and stored in DPBS (Sigma-Aldrich), supplemented with 0.5% FCS (Lot 42Q0154K; Gibco) and 1% penicillin/streptomycin solution at 4°C for future analysis.
Outgrowths from chimeric embryos
On days 6 or 7, presumptive porcine chimeric (monkey or miPSCs) blastocysts were mechanically separated from the zona pellucida and cut using ophthalmic scissors (Bausch & Lomb GmbH). Thereafter, embryos were cultured on a monolayer of mitomycin C-treated mouse embryonic fibroblasts in iPSC-20% or mESC medium. Media were changed every 48 hours and outgrowths were determined microscopically.
In vivo chimera assay
Next, we performed embryo transfer experiments to test whether the injected monkey and miPSCs can give rise to chimeric embryos and fetuses.
Peripubertal German Landrace gilts (approximately 7–9 months of age) were used either as embryo donors or recipients and were synchronized as described (Petersen et al., 2008). Briefly, gilts were fed with 5 mL/(day·gilt) altrenogest (Regumate®, 4 mg/mL; MSD Animal Health) for 13 days. On the last day of altrenogest feeding, sows were treated with 1500 IU PMSG (Intergonan 240 I.E./mL; MSD Animal Health). Ovulation was induced by intramuscular injections of 500 IU of hCG (Ovogest 300 I.E./mL; MSD Animal Health) 76 hours later. The sows were artificially inseminated twice at 40 and 48 hours after hCG administration with semen from wild-type boars diluted in Androhep® Plus solution (Minitube).
Four days (for injection of cyiPSCs at P.55) and 5 days (for injection of miPSCs at P.25) after insemination, sows were slaughtered, and the uterine horns were flushed with DPBS medium (AppliChem) supplemented with 1% NBCS (Gibco). Embryos were morphologically evaluated, washed two times in Opti-MEM I (1 × ) + GlutaMAX-I reduced serum medium (Gibco) supplemented with 10% FCS (Lot 42Q0154K; Gibco) and used for microinjection.
In vivo collected embryos were injected as described above. After injection, porcine-monkey chimeric embryos were incubated in mixed medium (3:1) of PZM-3 and iPSC-20% and porcine-mouse chimeric embryos in mESC medium for 1–2 hours. Thereafter, embryos were washed once and collected into the transfer catheter in Opti-MEM I (1 × ) + GlutaMAX-I reduced serum medium (Gibco) supplemented with 10% FCS (Lot 42Q0154K; Gibco) and stem cell medium (3:1). Embryos were surgically transferred into the uterine horns of recipient sows (−24 hours asynchronous to embryo donor sows). Pregnancies were terminated 6/7 days later, and the uterine horns were excised and flushed with PBS solution (Sigma-Aldrich) supplemented with 1% NBCS (Gibco). Recovered embryos were fixed for further analysis.
Evaluation of chimerism in porcine embryos
Porcine parthenogenetic embryos from days 6 to 7 and in vivo flushed embryos from day 11 injected with iPSCs were fixed in 3.7% formaldehyde solution (Honeywell Riedel-de Haën™) for 15 minutes at room temperature. Embryonic nuclei were stained either with NucRed™ Live 647 ReadyProbes™ Reagent (Thermo Fisher Scientific) for 30 minutes at room temperature or with 0.2 μM SiR-DNA (Spirochrome) for 30 minutes at 37°C. Thereafter, embryos were transferred onto a glass slide within a droplet of PBS (Sigma-Aldrich).
Confocal imaging of porcine embryos had been established previously (Tiedemann et al., 2014). Porcine blastocysts were analyzed with a 20 × Plan-Apochromat (NA 0.75) at the LSM510 (Carl Zeiss Micro Imaging GmbH) in “multi-tracking-mode” for comparative imaging of different fluorescence markers. A total of 10–25 medial optical sections of the blastocysts at days 6–7 with a thickness of 3.1 and 2.5 μm intersection differences was imaged covering the inner cell mass (ICM) and adjacent TE. Thus, stacks of 264 × 264 μm and a thickness of 30–50 μm were investigated. In addition, nine optical sections at a thickness of 3.2 and 2.1 μm intersection differences were taken from day 11 embryos.
Thereafter, stacks of 400 × 400 μm and a thickness of 16 μm were generated. The three-dimensional-projections of the stack were used for localization of cyiPSCs and miPSCs in porcine embryos. Also, detailed analyses for E-cadherin connection were performed with the 40 × Plan-Apochromat (1.3 NA) in volumes of 132 × 132 × 7 μm.
Immunofluorescence
Fixed cyiPSCs or parthenogenetic blastocysts injected with iPSCs were washed three times in 0.02 M PBS supplemented with 0.5% goat/donkey serum (Sigma-Aldrich). Thereafter, embryos were permeabilized in 0.2% Triton-X 100 (Merck) for 1 hour and then boiled in 0.01 M citrate buffer (Roth). Blocking was performed in 0.02 M PBS supplemented with 5% goat/donkey serum for 1 hour. Then iPSCs or embryos were incubated with primary antibodies diluted in 0.02 M PBS/0.5% goat/donkey serum for 24 hours at 4°C.
Primary antibodies used for staining were purified mouse anti-E-cadherin (1:10,000 for porcine embryos and 1:1000 for cyiPSCs; BD Transduction Laboratories™) and anti-CDX2, clone CDX2-88 (1:200; BioGenex). After washing three times in 0.02 M PBS/0.5% goat/donkey serum, embryos/iPSCs were incubated for 2 hours at room temperature with Alexa Fluor™-conjugated goat-anti-mouse IgG (embryos; 1:200; Invitrogen) or Cy3-conjugated donkey-anti-mouse IgG (cyiPSCs; 1:200; Jackson ImmunoResearch) antibody. Embryos/iPSCs incubated with isotype control as the primary antibodies (mouse IgG1 for CDX2 and mouse IgG2a for E-cadherin, both from Santa Cruz Biotechnology) and embryos/iPSCs incubated in the absence of the primary antibody were used as negative controls.
Nuclei of the embryos/iPSCs were counterstained with NucRed Live 647 ReadyProbes Reagent or with 0.2 μM SiR-DNA and examined immediately using either Axio Observer A1 microscope (Carl Zeiss MicroImaging GmbH) or a confocal imaging system LSM510 (Carl Zeiss MicroImaging GmbH).
Statistical analysis
Data were analyzed using the statistical software packages RStudio, Version 1.1.453 (2009–2018; RStudio, Inc.) and SAS 9.4 (2002–2012; SAS Institute, Inc., Cary, NC).
To test the effect of stem cell media on blastocyst development grades on days 7 and 8, respectively, a series of Fisher's exact tests was performed, medium “PZM-3” serving as a control. After a Bonferroni correction for multiple testing, a p-value <0.006 was considered statistically significant. One-way analysis of variance was executed to analyze the effect of stem cell media on the total cell number in day 8 blastocysts. To compare each of the experimental groups with the single control “PZM-3,” a Dunnett's post hoc test was applied. Residuals were approximately normally distributed. A p-value <0.05 was considered statistically significant.
To test the relationship between Venus positive/negative blastocysts on days 4 and 6, a chi-square test was performed. A p-value <0.05 was considered statistically significant.
Results
Developmental competence of porcine parthenogenetic blastocysts in different stem cell media
In view of the abundant availability and low cost, porcine parthenogenetically activated embryos were employed as an alternative source of porcine blastocysts in the in vitro experiments (Grupen, 2014). Parthenogenetically activated porcine oocytes can reach the blastocyst stage and after transfer to recipients, can even develop to fetuses up to day 29 of gestation (Kure-bayashi et al., 2000).
The culture media used for cynomolgus or miPSCs and porcine embryos are totally different. The culture conditions for combined cultivation of both embryos and iPSCs need to be optimized to allow the successful generation of interspecies chimeras.
Our initial results indicated clearly that miPSCs and cyiPSCs do not survive under culture conditions that are used typically for porcine embryos (PZM-3 medium). Similarly, it has been reported that human iPSCs injected into porcine parthenogenetic blastocysts underwent apoptosis under porcine culture conditions (Das et al., 2020). We cultured porcine blastocysts from day 6 in mESC medium and this medium was compatible with in vitro development of porcine embryos. In a similar approach, some benefit was achieved when fetal bovine serum (FBS) was added to the culture medium by day 5 (late morula/early blastocyst stage) and 80% of the embryos cultured developed into hatched blastocysts by day 8 (Dobrinsky et al., 1996).
Thereafter, we developed an effective in vitro culture system that enabled development of both cyiPSCs and porcine blastocysts. First, parthenogenetic blastocysts from day 6 were cultured for 1–2 days in either PZM-3 medium or eight different stem cell media, including iPSC-10%, iPSC-20%, conditioned iPSC medium (Condit.), StemPro® hESC SFM (StemPro), mTeSR™ 1, mTeSR™ 2, NutriStem® hPSC XF (NutriStem), and E8™ medium. The highest development of porcine blastocysts was achieved in iPSC-20% medium (Fig. 1A, B). The number of hatched blastocysts on day 7 was significantly higher in iPSC-20% medium than in the commonly used porcine PZM-3 medium (Fig. 1A). Compared to PZM-3 medium, a significantly lower number of degenerated blastocysts was found in iPSC-20% medium.

In vitro development of porcine parthenogenetic blastocysts cultured in different stem cell media. Blastocysts from days 7
Similarly, the number of hatched blastocysts on day 8 was significantly higher in iPSC-20% medium than in PZM-3 medium (Fig. 1B). We found significantly fewer not-expanded blastocysts and degenerated embryos on day 8 cultured in iPSC-20% medium compared to PZM-3 medium (Fig. 1B). The number of nuclei per blastocyst was significantly higher in iPSC-20% medium than in the PZM-3 medium (Fig. 2). Based on these results, we decided to use iPSC-20% medium for culture of porcine-monkey chimeric blastocysts from day 6.

Total cell number of porcine blastocysts on day 8 cultured in different stem cell media. Values with different letters between blastocyst from stem cell media were significant (p < 0.05). Numbers in parentheses indicate the whole number of embryos tested per group. Data shown as mean ± SEM. The experiment was performed three times. The lower picture shows representative fluorescence images of porcine parthenogenetic blastocysts on day 8 cultured in different stem cell media. Nuclei were stained with Hoechst 33342. SEM, standard error of the mean.
Subsequently, we tested the iPSC-20% medium for culture of 3-day-old porcine parthenogenetic embryos. However, these early stages developed poorly in pure iPSC-20% or mESC medium (blastocyst rate: mESC medium [1.3%], iPSC-20% medium [3.7%], and PZM-3 medium [41%]). The best results were achieved with PZM-3 mixed with 10% or 25% iPSC-20% medium (38.8% and 30% blastocyst rates, respectively) (Burchardt et al., 2015).
Stability of Venus expression during differentiation and karyotyping of iPSCs
One iPSC line from either mouse or cynomolgus monkey, each transgenic with Venus protein, was used in the interspecies chimera assay. Both iPSC lines maintained stable Venus expression during spontaneous differentiation, during in vitro culture for 16–26 days, and could thus be successfully traced in porcine embryos or fetuses by fluorescence microscopy (Supplementary Figs. S1A, B and S2A, B). Analyses of both iPSC lines revealed a normal karyotype with 40 chromosomes in miPSCs at passage 18 (Talluri et al., 2014) and 42 chromosomes in monkey iPSCs at passage 32 (Wunderlich et al., 2012).
However, Venus transgenic miPSCs at passage 25 showed trisomy for chromosome 11 in 2/7 metaphases (41,XY,Ts11, inc[2]/40,XY,inc[5]) and after 45 passages in all metaphases (10/10) 41,XY,Ts11[10] (Supplementary Fig. S1C). In addition, karyotyping of cyiPSCs at passage 58 revealed trisomy of chromosome 12 in all metaphases (43,XY,Ts12[21]) (Supplementary Fig. S2C).
Interspecies in vitro chimera assay using parthenogenetic embryos
Monkey and miPSC lines used for the generation of interspecies chimeras maintain stable Venus/GFP-expression after spontaneous differentiation and therefore can be traced in porcine embryos or fetuses by fluorescence microscopy.
In this study, we examined survival and proliferation of cyiPSCs in porcine preimplantation embryos (Fig. 3A–D). Injected parthenogenetic embryos were cultured in vitro for 24 hours (day 6 embryos) or 48 hours (day 4 embryos) and analyzed using confocal microscopy. At the same time, we determined whether the developmental stage of the host embryos influenced survival of cyiPSCs in porcine embryos. Injection into porcine embryos from day 4 resulted in 38 of 51 blastocysts (74.5%), with an average of 19 Venus-positive cyiPSCs per embryo, while only 43% (57/133) of blastocysts contained an average of 10 Venus-positive cyiPSCs after injection of day 6 embryos (Fig. 3B).
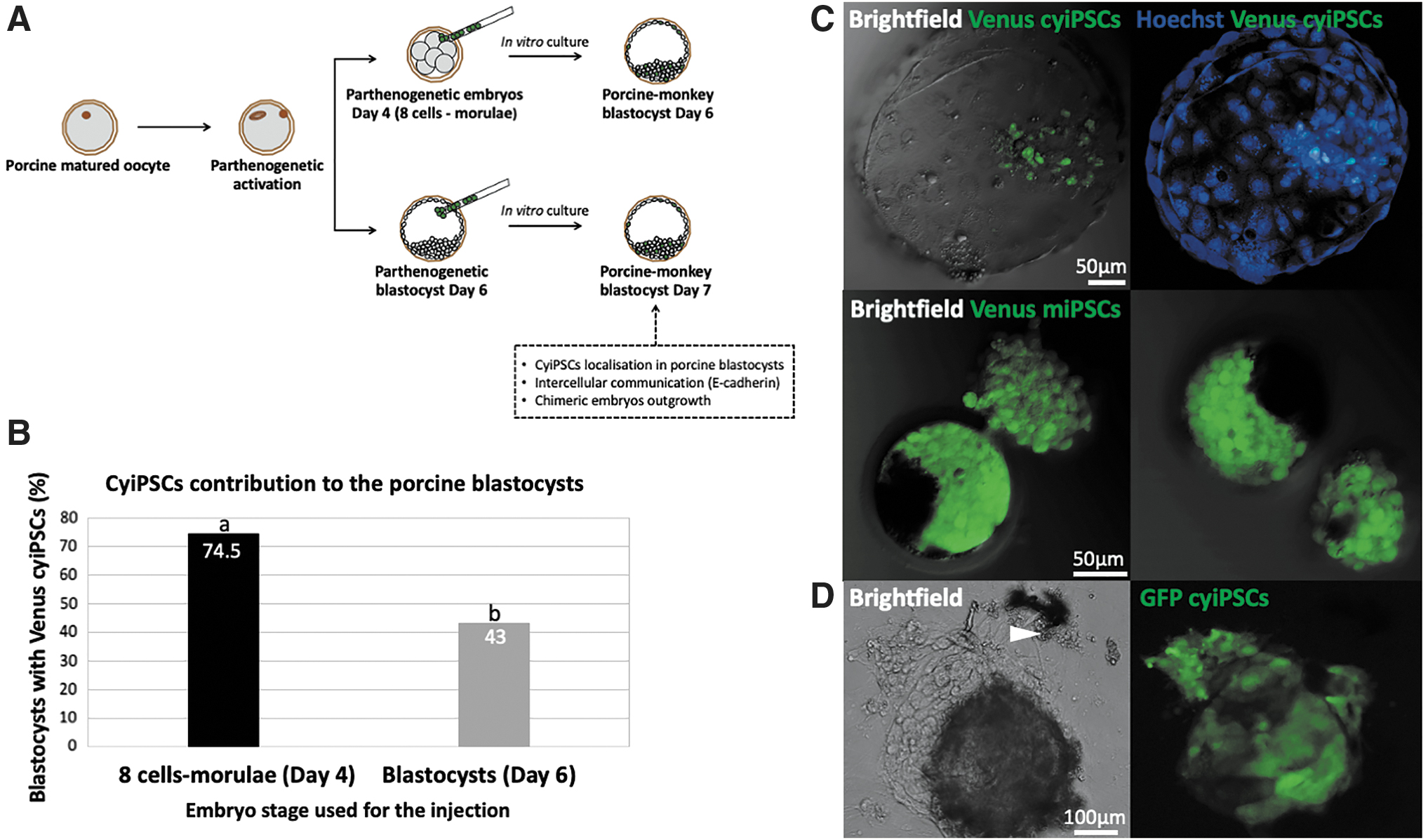
In vitro interspecies chimera assay.
After nuclei staining, confocal optical sectioning demonstrated Venus-expressing cells located in the ICM in 31% (64/206) of the blastocysts after injection on day 4. We observed 16% (32/206) blastocysts with cyiPSCs in both the ICM and TE (Fig. 4A, B). The majority [53% (110/206)] of cyiPSCs was observed in trophectodermal cells, as shown after staining with the TE-specific marker CDX2 (Fig. 5). In vitro cultured cyiPSCs can differentiate into TE and stained positive for CDX2 (Fig. 5A, B). However, none of the Venus-expressing cyiPSCs expressed CDX2 when located among their CDX2-positive porcine counterparts (n = 28) (Fig. 5C).

Localization of cyiPSCs in porcine parthenogenetic embryos.
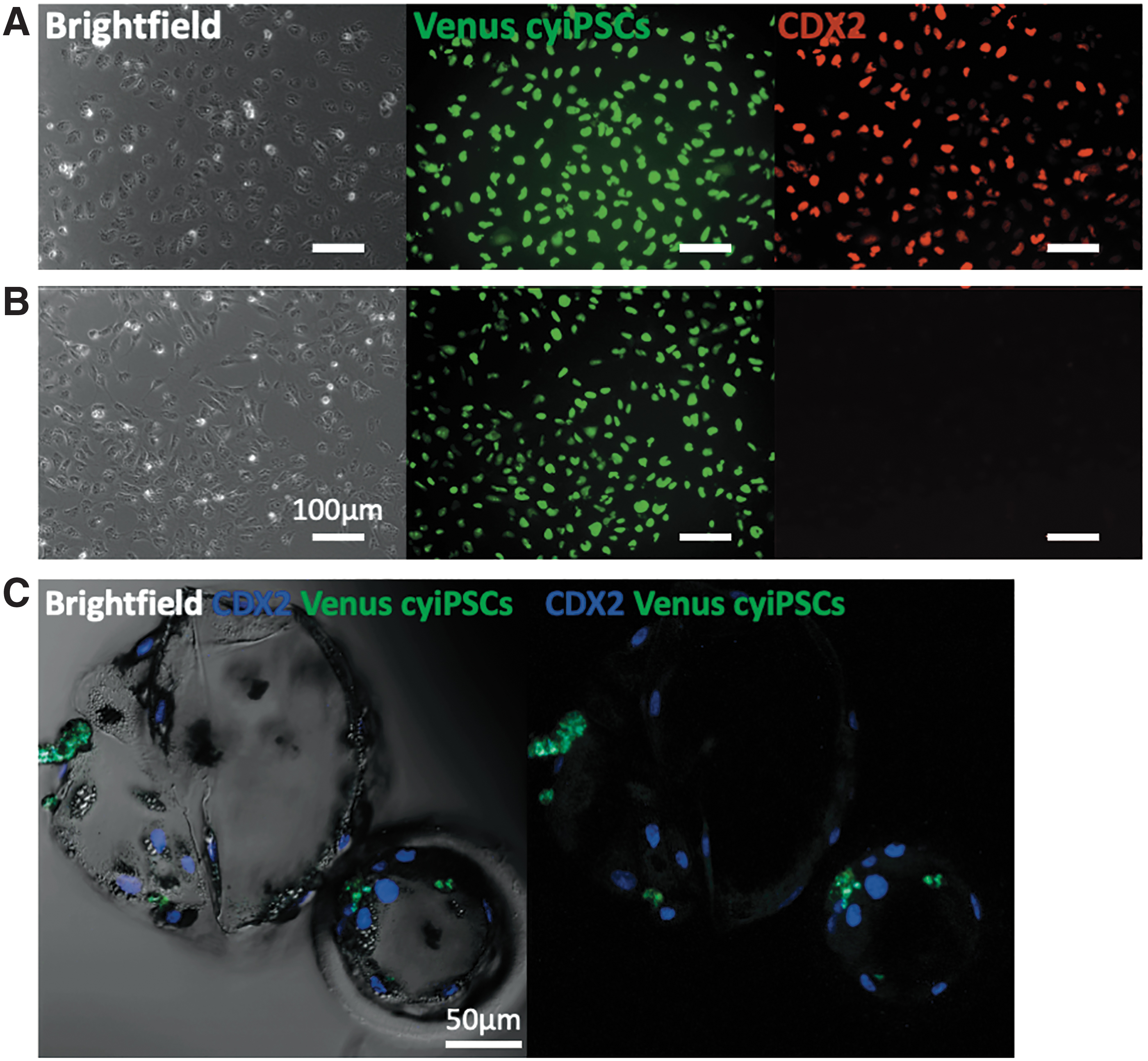
Expression of the trophoblast-specific marker CDX2 in differentiated cyiPSCs and porcine-monkey blastocysts on day 6. Representative fluorescence images of
Finally, we investigated E-cadherin-based cell-cell contacts between cyiPSCs and porcine embryonic cells. Interestingly, 77% (119/155) of porcine blastocysts showed intimate connection between cyiPSCs and porcine embryonic cells (Fig. 6A). The high dilution of E-cadherin antibody used for staining allowed detection of cell-cell connection driven only from the porcine embryo (Fig. 6A, B). In contrast, miPSCs injected into porcine parthenogenetic embryos overgrew the blastocoel almost completely, rendering it impossible to accurately count and localize numbers of miPSCs (Fig. 3C).
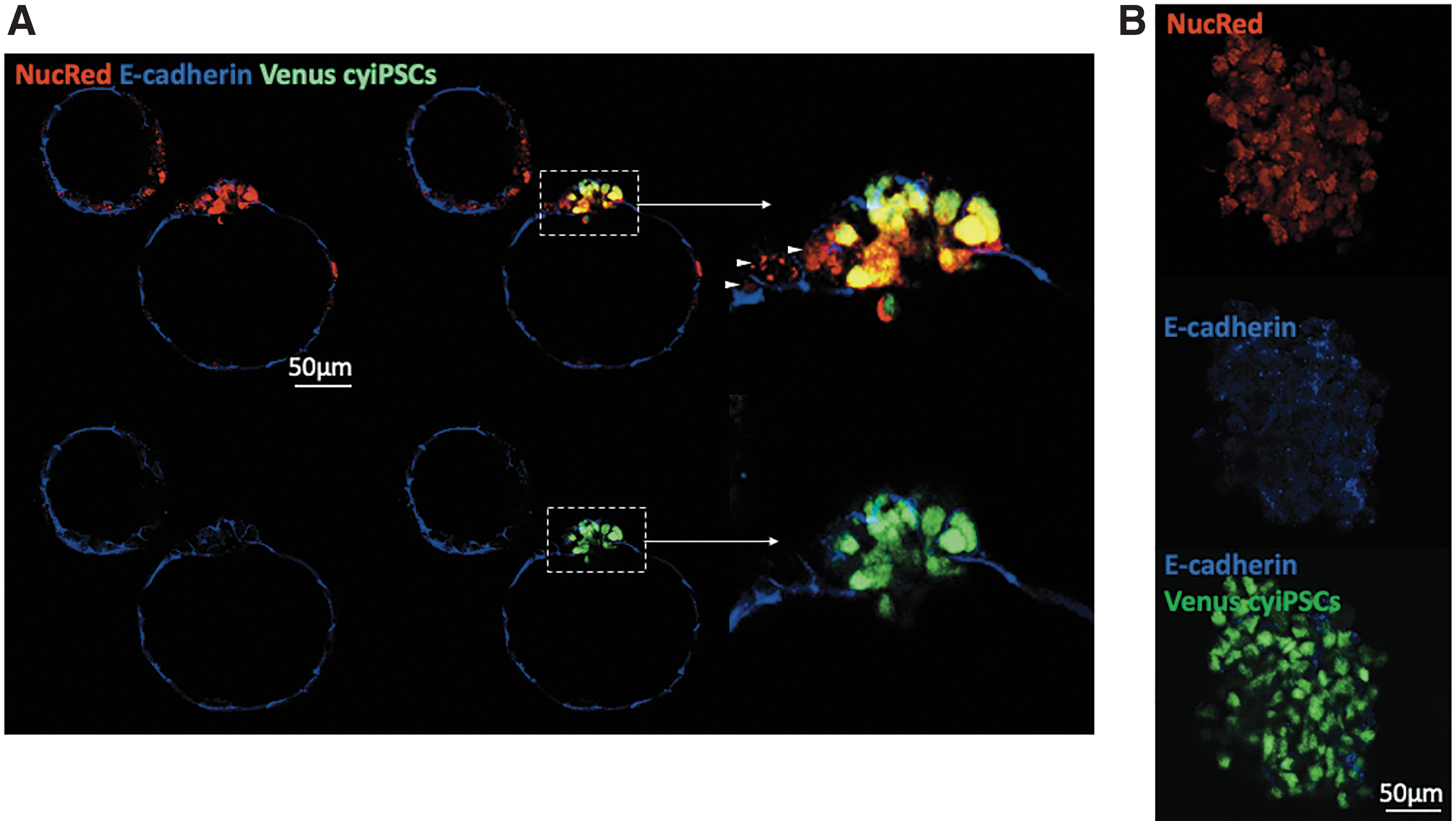
E-cadherin expression in porcine-monkey parthenogenetic blastocysts from day 6. Representative confocal images of
Survival, proliferation, and intermixing of injected iPSCs with porcine embryonic cells were also evaluated after attachment of chimeric embryos onto MEF cells. In vitro cultured blastocysts (17.4%, 72/414) successfully attached to the dish and cyiPSCs proliferated rapidly (Fig. 3D). Unfortunately, porcine ICM did not proliferate after attachment and porcine outgrowths could not be observed (Fig. 3D).
Interspecies in vivo chimera assay using in vivo derived embryos
After the in vitro chimera assay had confirmed that both monkey and miPSCs can be detected in porcine parthenogenetic embryos, we investigated whether these cells can contribute to in vivo embryonic development (Fig. 7A). We transferred a total of 88 porcine in vivo derived embryos (39 4-cells -8-16-cells and 49 morulae) after injection of cyiPSCs at passage 55 into two recipient wild-type sows (Table 1). After seven days, 34 embryos (38.6%) could be recollected and were analyzed for the presence of injected cells within the in vivo developing conceptuses. We analyzed a total of 20 porcine-monkey embryos from day 11 and did not find any Venus cyiPSC localized in the embryonic disc (Fig. 7B). Half (10/20) of the analyzed embryos possessed only single Venus cyiPSCs in the TE (Fig. 7B).

The production and characterization of in vivo interspecies embryos.
Generation of Porcine-Monkey Interspecies Embryos
TE, trophectoderm.
Next, we explored the possibility of interspecies chimera formation after injection of Venus-expressing miPSCs at passage 27 into 117 porcine in vivo derived embryos (25 8-16-cells and 92 blastocysts), which were transferred into three recipient wild-type sows (Table 2). From 117 transferred embryos, 64 (55%) were recollected on day 11. Venus miPSCs were discovered in the TE of all embryos recovered from recipient sow number 823 (an average of 96 miPSCs per embryo) and recipient sow number 827 (an average of 40 miPSCs per embryo) (Fig. 7C and Table 2). In two cases, we found Venus miPS cell clumps (15–20 cells) localized in trophoblastic tissue. Few embryos (six embryos from recipient sow number 823 and three embryos from recipient sow number 827) had fluorescent cell clumps within the porcine embryonic disc (Fig. 7C).
Generation of Porcine-Mouse Interspecies Embryos
ED, embryonic disc; miPSCs, mouse induced pluripotent stem cells; n.a., not analyzed.
Discussion
The ability of stem cells to contribute to chimeric offspring with germline transmission is an eminent criterion for defining pluripotency. Embryonic and iPSCs have been successfully established in the mouse (Evans and Kaufman, 1981; Martin, 1981; Okita et al., 2007), rat (Buehr et al., 2008; Hamanaka et al., 2011; Li et al., 2008), non-human primates (Thomson et al., 1995, 1996), and human (Thomson et al., 1998). Both, human and non-human primate pluripotent ESCs, either derived from the ICM or iPSCs generated after somatic cell reprogramming, were categorized as having a primed pluripotent state and were considered to be equivalent to mouse epiblast stem cells, which correspond to the pluripotency status of early postimplantation day 5.5 (E5.5) mouse embryos (Nichols and Smith, 2009; Reubinoff et al., 2000; Tesar et al., 2007).
Contrary to naive stem cells, primed stem cells seem to have a more restricted developmental potential in vivo and do not contribute to chimeric tissues of the embryo proper after introduction into preimplantation host embryos (Nichols and Smith, 2009).
The ability of cultured pluripotent stem cells to contribute to chimera formation has been extensively tested in rodents. A major obstacle in human stem cell research is that ethical considerations prevent from testing their capacity to generate chimeras with germline contribution. Therefore, engraftment of human stem cells into animal host embryos has been proposed as a suitable alternative approach for evaluating pluripotency rather than in vitro differentiation or the teratoma assay (Masaki et al., 2015). Recently, several groups have studied developmental potency and behavior of human stem cells in the interspecies chimera model using mouse embryos (Gafni et al., 2013; James et al., 2006). Interestingly, human stem cells injected into mouse epiblast stage embryos differentiated into the three primary tissue layers and participated in normal mouse development after 2 days of in vitro culture (Mascetti and Pedersen, 2016b).
Different culture protocols have been developed to convert primed human or non-human primate stem cells into the naive state (Chen et al., 2015; Fang et al., 2014; Theunissen et al., 2016; Wu et al., 2015). The naive-like human stem cells contributed to mouse preimplantation embryos, although at low frequency (Gafni et al., 2013; Theunissen et al., 2014). Thus, host embryos derived from species evolutionarily closer to humans would be needed for the successful generation of interspecies chimeras. In this study, we employed porcine preimplantation embryos as a host for interspecies chimeras using two developmentally distinct cell types from two different species: cyiPSCs and miPSCs. We evaluated whether the different sources of iPSCs could affect engraftment in vitro and in vivo in the interspecies chimera model using porcine host embryos.
One of the important factors affecting the efficiency of interspecies in vitro chimera production is the selection of an appropriate culture medium that is compatible with survival and proliferation of both, the injected stem cells and the host embryos. CyiPSCs were maintained in distinct culture conditions and could not proliferate in PZM-3 medium, which is commonly used for in vitro culture of porcine embryos.
Therefore, initially, we tested a variety of established stem cell culture media and mixtures thereof with commonly used embryo culture systems in their suitability to support the development of porcine parthenogenetic blastocysts. Development of parthenogenetic porcine blastocysts and the total cell number were significantly higher in iPSC-20% medium than in any of the other media. The iPSC-20% medium was supplemented with KSR and contained small organic molecules, trace elements, and three proteins, including insulin, transferrin, and albumin (Sakurai et al., 2015), which provided better support for development and enabled porcine blastocyst hatching.
These results confirmed previous work showing that addition of KSR to the medium significantly increased survival and hatching rates of blastocysts (Sakurai et al., 2015), possibly due to KSR that promotes lipid metabolism and thereby provides the necessary energy source to enhance embryo development. and even hatching from the zona pellucida (Jin et al., 2018; Sakurai et al., 2015). Our results demonstrate that commercially available serum-free stem cell media tested in this study did not improve viability and development of porcine blastocysts. mESC culture medium supplemented with FBS facilitated in vitro development of interspecies porcine-mouse embryos. This supports previous findings that FCS supplementation promotes development from morulae to blastocysts and hatching of porcine embryos (Dobrinsky et al., 1996).
Another critical factor of chimera production is the developmental stage of the host embryo. To address this question, we traced the fate of cyiPSCs after injection into porcine embryos from days 4 to 6. Our results show that injection at an earlier stage of development (8-cell—morulae) may increase survival of the injected cells compared to blastocyst injections. This is consistent with previous observations in the mouse that injection of stem cells into 8-cell mouse embryos resulted in higher cell contribution and germline transmission compared to blastocyst injections (Poueymirou et al., 2007; Tokunaga and Tsunoda, 1992).
Similarly, the generation of intraspecies chimeras using porcine parthenogenetic 4-8-cell-stage embryos (day 3) and porcine parthenogenetic ICMs (day 6) resulted in the production of chimeric blastocysts (Burchardt et al., 2015; Nakano et al., 2013). Based on our results, it is reasonable to assume that cleavage-stage embryos may provide a better niche for the injected stem cells rather than blastocysts with an already formed ICM. Injected stem cells were covered and engulfed by the host blastomeres, thus facilitating integration and survival of the cells in the embryo proper (Hanoka et al., 1987). Contrary to cyiPSCs, miPSCs proliferated rapidly after injection into porcine 4-day-old embryos. To reduce the risk of overgrowth in early developmental stages, which may compromise embryo development, we injected miPSCs into blastocyst stages.
The presence or absence of specific cell to cell communication elements between injected and host cells within the chimera may also be critical for interspecies chimera generation. One of the main intercellular adhesion elements is E-cadherin (Pieters and van Roy, 2014), which is essential for epithelization of early embryos, cell rearrangement, organization, and homeostasis of tissues and organs (Edelman, 1984; Li et al., 2012; Pieters and van Roy, 2014; Soncin and Ward, 2011).
The presence of functional E-cadherin is important for compaction of mouse preimplantation embryos and E-cadherin-negative blastocysts failed to form a blastocyst cavity and trophectodermal epithelium (Larue et al., 1994). In ESCs, E-cadherin-mediated cell-to-cell contacts play a crucial role in proliferation, cell survival, and maintenance of pluripotency (Li et al., 2010; Redmer et al., 2011). The inefficient chimera formation after injection of E-cadherin knockout mESCs into blastocysts suggests that adhesion of stem cells to the inner cell mass of blastocysts is essential for integration of injected cells (Larue et al., 1996).
A detailed analysis of individual interspecies blastocysts revealed that cyiPSCs were closely associated by E-cadherin-mediated cell-to-cell contacts with the porcine cells within 2 days of in vitro culture. However, once the outgrowths from interspecies chimeric embryos were established, neither monkey nor miPSCs intermixing with the outgrowing porcine cells was observed. The reason why porcine cells were not intermixing with monkey or miPSCs may suggest that the miPSC and mESC culture conditions do not support the survival and proliferation of porcine cells.
Previously, a study using mouse host embryos for injection of rhesus or baboon ESCs revealed that murine cells grew separately from primate ESCs (Simerly et al., 2011). Probably, cell-to-cell contacts emerging during blastocyst development were not sufficient for incorporation into interspecies outgrowths after attachment to the bottom of the dish (Simerly et al., 2011).
Integration of injected stem cells into the host ICM is critical for the contribution to all tissues of the emerging chimeric fetus (Simerly et al., 2011). In this study, the cyiPSCs injected into porcine blastocysts proliferated after 2 days in vitro culture and the vast majority could be localized in the TE of parthenogenetic blastocysts. However, parthenogenetic embryos exhibited reduced total cell numbers and fewer cells in the inner cell mass compared to blastocysts derived from in vitro fertilized oocytes (Hao et al., 2004; Kurihara et al., 2002). In these chimeric embryos, monkey cells located in the TE did not express the TE-specific marker CDX2, which led us to conclude that cyiPSCs survived in the TE of porcine blastocysts, but did not adopt the trophectodermal fate.
The potential of monkey cells to integrate into chimeric offspring with extensive contribution to the embryo proper and extraembryonic lineages was demonstrated after aggregation of three or more cleaving monkey embryos (Polejaeva and Mitalipov, 2013; Tachibana et al., 2012). However, injection of whole monkey ICMs into host blastocysts resulted in extensive contribution to the extraembryonic compartment consisting of chorionic and amniotic tissues and with very low chimeric contribution to liver and spleen (Polejaeva and Mitalipov, 2013; Tachibana et al., 2012).
It is well known that chromosomal aberrations may appear during prolonged in vitro culture periods and indeed have regularly been observed in human pluripotent stem cells (Imreh et al., 2006; Ludwig et al., 2006; Mitalipova et al., 2005). However, often detailed information about the karyotype stability of stem cells after long-term-culture has not been provided (James et al., 2006; Simerly et al., 2011; Yamaguchi et al., 2018). This is why in this study, we analyzed the karyotype of used stem cells at various intervals after establishment of culture. Not surprisingly, karyotyping revealed a trisomy of chromosome 12 in late passages of the cyiPSC line used in the interspecies chimera assay.
The higher expression of some genes located on human chromosome 12, for example, human Stella related (STELLAR), NANOG, growth differentiation factor-3 (GDF3) (Clark et al., 2004; Turinetto et al., 2017), can be the reason for an advantage of these human cell clones in culture (Spits et al., 2008). In the cynomolgus monkey, these genes are located on other chromosomes, which makes it extremely unlikely that cyiPSC behavior is affected. In support of this assumption, karyotypically abnormal cyiPSCs retained normal stem cell morphology and the ability to differentiate in vitro.
In the cyiPSC clone used in this study, it was obvious that the trisomy did not provide a selective advantage in growth rates in comparison to the original cyiPSC clone used for genetic modification. Overall, our results indicate that regular monitoring of the cell karyotype could be beneficial, but there is no evidence that the observed chromosomal aberrations would have affected the interspecies formation capacity.
In this study, monkey iPSCs injected into in vivo generated porcine embryos failed to yield any embryo chimeras after in vivo transfer. The single surviving cyiPSCs were localized exclusively in the TE of porcine in vivo derived embryos on day 11. These findings have to be considered in light of the fact that cyiPSCs used in our experiment had been shown to possess the capacity to differentiate into the three germ layers in vitro and formed teratomas with tissues of all three germ layers; however, their contribution into cynomolgus monkey embryos had not been tested (Wunderlich et al., 2012).
Remarkably, we observed that mouse naive iPSCs rapidly expanded and overgrew the blastocoel of the porcine parthenogenetic embryos. We cannot completely rule out the possibility that the additional chromosome 11 found in miPSCs after prolonged culture was associated with a better survival of miPSCs in porcine embryos. In the mouse, it is revealed that over one third of the mESCs possess large chromosomal genetic aberration, mainly trisomy 8 and 11 that are associated with a selective growth advantage (Gaztelumendi and Nogués, 2014; Liang and Zhang, 2013; Sugawara et al., 2006; Weissbein et al., 2014). One hypothesis is that trisomy 11 may confer a proliferation benefit to miPSCs used for chimera assays.
After injection of miPSCs into porcine in vivo derived embryos, clusters of iPSCs were found in the host epiblast and also numerous single cells within the TE, suggesting that miPSCs can migrate into both, the epiblast and TE of host embryos. However, it will be important to evaluate if the miPSCs are growing ectopically in the porcine epiblast or become ultimately integrated in the host embryo. Recent studies have identified that some mESCs contained a subpopulation of cells that can contribute to both, embryonic and extraembryonic lineages as well (Macfarlan et al., 2012; Morgani et al., 2013). Also, specific chemical cocktails enabled the derivation of stem cells with an extended pluripotent potential from established PSC lines from mouse and human blastocysts, and iPSCs after cell reprogramming. These cells possess the capacity to form both embryonic and extraembryonic cell derivatives (Yang et al., 2017b).
In summary, the results reported in this study demonstrate that the interspecies chimera assay using porcine embryos as host serves as promising model for studying chimeric potency and developmental potential of stem cells from different species. We found that, iPSCs behaved similar after exposure to in vitro interspecies chimeric preimplantation embryos compared with that of chimeras developed in vivo. Our results also emphasize the necessity for regular karyotyping of stem cells as prolonged cultures are well known to be associated with chromosomal aberrations.
In this study, we injected stem cells from one well-characterized culture from both, mouse and cynomolgus, which are representative for pluripotent cells from these species. However, we cannot completely rule out the possibility that other stem cell lines would have yielded different results. To clarify this question, various stem cell lines would have to be compared after injection into early porcine embryos. Moreover, conversion of primed state cyiPSCs into a true naive state with a higher chimeric potential could improve the contribution of cyiPSCs into porcine development. Further investigations are required to determine the competency of iPSCs for chimera formation in postimplantation porcine conceptuses.
Footnotes
Acknowledgments
The authors thank S. Petkov for his help with the optimization of blastocyst injections and T. Talluri and E. Mall for providing Venus-expressing miPSCs. The authors also thank the staff from the pig facility, G. Moeller, E. Kufeld, T. Peker, and J. Kun, for taking excellent care of the pigs.
Author Disclosure Statement
The authors declare they have no competing financial interests.
Funding Information
The project was financially supported by German Research Foundation (DFG) (EXC 62; Cluster of Excellence REBIRTH).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.