
Abstract
Somatic cell nuclear transfer (SCNT) is an important technique for biological science research. Cytoplasm injection cloning technology (CICT) was developed to improve the reprogramming efficiency as well as to overcome the limitations of SCNT. CICT uses an additional cytoplasm fused with an enucleated oocyte to restore the cytoplasmic volume of the cloned embryo, and this method could improve the reprogramming efficiency of the cloned embryo. In this study, we show that CICT can be adapted to mouse species to overcome the inefficiency of the SCNT method. In this study, results indicate that the two-cell embryo and blastocyst rates of cloned embryos with the use of the CICT method were significantly higher (p < 0.05) than that of the SCNT method (96.6% ± 1.1% vs. 86.7% ± 6.0%, 29.5% ± 2.6% vs. 22.1% ± 3.0%, respectively). Furthermore, the apoptotic cell number per blastocyst was significantly lower in the CICT group than that in the SCNT group (1.7 ± 0.2 vs. 2.9 ± 0.3, p < 0.05). Moreover, the acH3K9/K14 expression level in the CICT group was greater than that of the SCNT group (p < 0.05), and the relative acH3K56 level in the CICT group was significantly (p < 0.05) higher than that in the SCNT group. These results indicate that CICT helps improve the in vitro developmental competence and quality of cloned embryos.
Introduction
The somatic cell nuclear transfer (SCNT) technique has resulted in production of offspring in many species, including sheep, cattle, pig, dog, cat, and monkey (Campbell et al., 1996; Holker et al., 2005; Jang et al., 2007; Meng et al., 1997; Yin et al., 2008; Zakhartchenko et al., 2001). The SCNT method offers many opportunities in research that involve the cloning of transgenic and endangered species. However, the efficiency of SCNT is still very low. In mammalian species, the cloning efficiency is <5% of the reconstructed embryos developing into offspring (Whitworth and Prather, 2010; Wilmut et al., 2002).
It is unclear if the low efficiency of SCNT is due to the reprogramming factors not being sufficient for each embryo, the donor nucleus, or the cloning technique (Han et al., 2003; Kang et al., 2001). Incomplete reprogramming of nuclei can be revealed by abnormal methylation patterns that can be detected in the cloned embryo (Blelloch et al., 2006; Kong et al., 2014). A part of the cytoplasm is removed from enucleated oocytes, which can affect nuclear organization of the cytoskeleton. This can lead to inefficient gene expression during preimplantation development of the cloned embryos (Kwon et al., 2015; Samiec and Skrzyszowska, 2018; Whitworth and Prather, 2010).
To generate normal cloned animals, nuclei from the somatic donor cell need to be completely reprogrammed. Reprogramming of nuclei leads to a reduced genome methylation level and increased histone acetylation activity. In cloned embryos or offspring with insufficient or aberrant genome reprogramming, there are abnormalities and abnormal gene expression levels (Bourc'his et al., 2001; Dean et al., 2001; Santos et al., 2003; Zhang et al., 2016). The histone acetylation level has the potential to recruit transcriptional factors and is associated with the activation of gene transcription (Kurdistani et al., 2004; Wang et al., 2007). The histone methylation level has been associated with gene activation or silencing (Kouzarides, 2002; Martin and Zhang, 2005).
The embryo aggregation (EA) method was developed not only to improve the development and quality of embryos but also because of their effect on inducing change in the levels of methylation in cloned embryos (Buemo et al., 2016; Gambini et al., 2012). Handmade cloning (HMC) was invented to produce embryos without the need of micromanipulators. HMC involves two enucleated oocytes that are fused to construct one embryo. Both the cytoplasmic volume and embryo development rate are increased in the HMC cloned embryo (Du et al., 2007; Vajta et al., 2003).
However, previous research shows that extra cytoplasm of artificial giant (AG) oocytes, two to nine oocytes, which were fused by electrofusion and mechanical fusion, did not improve the reprogramming potential in mice (Sayaka et al., 2008). EA, HMC, and AG have a critical drawback: the reconstructed embryo using EA and HMC is in a zona-free condition. Zona-free embryos have the potential of disease transmission and damage during handling (Vajta et al., 2004; Verma et al., 2015).
Recently, cytoplasmic injection cloning technology (CICT) was developed, in which 30% of the cytoplasm from a donor oocyte is injected into the enucleated recipient oocyte, restoring a normal cytoplasmic volume, and the effects of this process on cloned bovine embryo development have been studied (Xu et al., 2019). Analysis of CICT cloned embryos shows that reprogramming of the nuclei had occurred and the embryo quality was improved, including total cell number (trophectoderm [TE]), genome methylation, and histone deacetylation, in the bovine embryo in comparison with the traditional cloning method. Moreover, CICT has been applied in cats, resulting in improved production of kittens from cloned cat embryos (Song et al., 2019).
In this study, we applied CICT to mouse cloning with the aim of improving the cloned embryo development in vitro. We overcame the inefficiency of mouse cloning by using an adaptation of CICT. We observed the effect of the cloning method on embryo development.
Materials and Methods
Chemicals
All of the reagents were purchased from Sigma Chemical Co. (St Louis, MO), unless otherwise stated.
Mice
B6D2F1 (C57BL/6 X DBA/2) mice were used for the nuclear transfer and donor nuclei. The Gyeongsang National University Institute of Animal Care and Use Committee approved all surgical procedures (GNU-161228-M0075).
Oocyte collection
Mature oocytes were collected from the oviducts of 4- to 8-week-old female mice. The female mice were induced to superovulate by injection of 5 IU of pregnant mare serum gonadotropin (Daesung Bio, Republic of Korea) and followed by 5 IU of human chorionic gonadotropin (hCG; Daesung Bio) 48 hours later. At 13–14 hours after hCG injection, oocyte–cumulus complexes were collected from the oviduct. The collected oocyte–cumulus complex was dropped in M2 with 0.1% bovine testicular hyaluronidase until the cumulus cells were dispersed. The cumulus-free oocytes were placed in CZB medium and kept in an incubator (37°C in 5% CO2) until use.
Preparation of donor cells
After oocyte collection, cumulus cells were placed in M2 + 0.1% hyaluronidase. Cumulus cells were washed several times in M2 and kept in M2 at room temperature (RT) until the injection step. Cumulus cells were from mice of the same strain, but collected from a different cell donor.
Enucleation of oocytes
A group of oocytes (∼20 in number) was placed in M2 with a 5-μg/mL cytochalasin B (CB) droplet for 7 minutes before enucleation began. Then, each oocyte was held with a pipette and rotated to find the metaphase II chromosome. The zona pellucida of the captured oocyte was drilled through by applying several piezo pulses (Prime Tech Ltd., Japan) at the tip of the enucleating pipette. The enucleating pipette was inserted through the zona pellucida, removing the Metaphase II (MII) complex through the enucleation pipette without damage to the oocyte. All of the oocytes in the group were enucleated as soon as possible (within 20 minutes) and transferred into CB-free CZB medium and kept in an incubator (37°C in 5% CO2).
Nuclear transfer
Enucleated oocytes were placed in an M2 droplet for SCNT and CICT. A single donor cell was immersed in Sendai virus (SV; Cosmo Bio, Japan) medium for 1 minute according to the manufacturer's protocol and injected into the perivitelline space of the enucleated oocyte (SCNT). In CICT, cytoplasm from a donor oocyte and donor cell was immersed in SV medium and injected into the perivitelline space of the enucleated oocyte. In this procedure, cytoplasm-donor oocytes and enucleated recipient oocytes were collected from the same donor mouse to produce a homoplasmy of the reconstructed oocytes. The CICT method requires 1.5 times more oocytes and required time than SCNT. The reconstructed oocytes, following the SCNT and CICT methods, were fused by the SV-mediated method (Song et al., 2011) and then transferred into CZB medium for 2 hours in an incubator (Fig. 1)
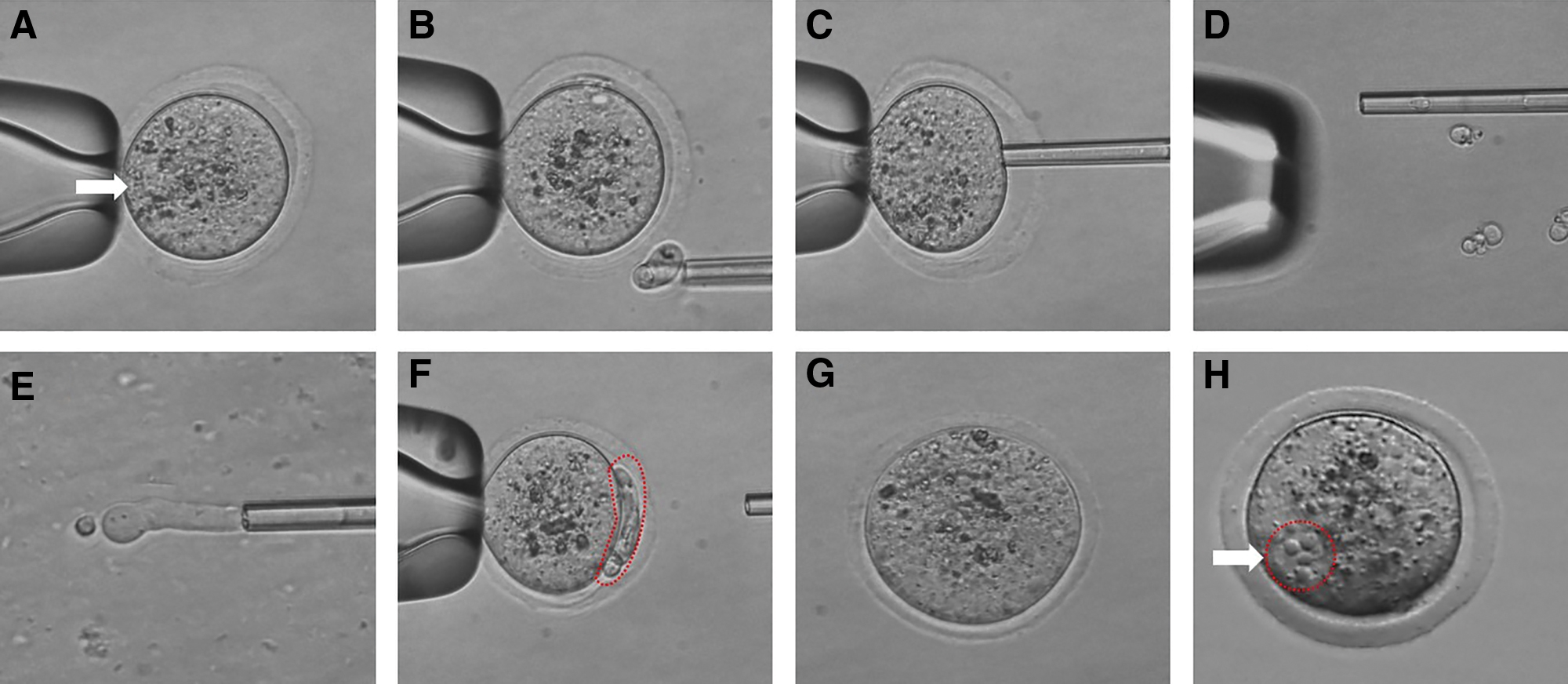
Procedure of cytoplasm injection cloning technology (CICT).
Oocyte activation
The reconstructed oocytes were kept in CZB medium for 2 hours in an incubator (37°C in 5% CO2) to confirm the fusion of the SCNT and CICT complex. A group of reconstructed oocytes was transferred into calcium-free CZB medium with 5 mM strontium chloride (SrCl2), 5 μg/mL CB, and 5 nM trichostatin A (TSA) for 6 hours in an incubator. After activation, reconstructed oocytes were transferred to CZB medium with 5 nM TSA for 2 hours in an incubator.
Presumptive cloned embryo culture
After the activation step, the fused oocytes were washed several times in CZB medium to remove the CB and TSA, and formation of pronucleus (PN) was checked after activation. Chemical-free oocytes were cultured in CZB medium for 72 hours in an incubator until they developed into blastocysts. The two-cell and four- to eight-cell embryos and blastocysts were checked at 24, 48, and 72 hours after activation. PN, cleavage, and blastocyst development competence were confirmed through a stereomicroscope (Olympus, Tokyo, Japan).
Assay of blastocyst quality
To assess the blastocyst, the total cell number and number of apoptotic cells were detected using a terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) system using a death detection kit (Fluorescein; Roche Diagnostics Corp.) (Dey et al., 2012). Fixed embryos were washed with polyvinylpyrrolidone–phosphate-buffered saline (PVP-PBS) (1 M PBS with 0.3% PVP) and incubated with PVP-PBS for 10 minutes at RT. The oocytes were incubated in 0.5% Triton X-100 with 0.1% sodium citrate for 30 minutes at RT and then washed three times in PVP-PBS. Treated oocytes were incubated with fluorescein-conjugated deoxyuridine triphosphate and terminal deoxynucleotide transferase in the dark for 1 hour.
After TUNEL staining, embryos were washed with PVP-PBS and incubated with 4′,6-diamidino-2-phenylindole (DAPI, 3 μg/mL) for 10 minutes, washed in PVP-PBS, and then mounted on a glass slide. The cell numbers and apoptotic cell numbers of blastocysts were counted using an epifluorescence microscope (Olympus IX71, Japan) equipped with a mercury lamp. A total of 15 blastocysts were analyzed per group.
Immunostaining
After activation, the reconstructed embryos were fixed in PBS containing 4% paraformaldehyde. The fixed embryos were washed twice using polyvinyl alcohol (PVA)-PBS with shaking, after which we added 1 μL of proteinase K for 5 minutes at RT. The treated embryos were washed and incubated in PBS containing 0.25% of bovine serum albumin and 0.1% of Triton X-100 for 90 minutes. The embryos were incubated with the primary antibody, mouse monoclonal anti-H3K9me2 (1:100 dilution; Abcam, Inc.) or mouse monoclonal anti-acH3K9/K14 (1:100 dilution; Santa Cruz Biotechnology, Inc.), in PVA-PBS overnight with shaking.
After the embryos were washed with PVA-PBS, they were incubated with the dye-conjugated secondary antibody, goat anti-mouse IgG-FITC (1:100 dilution; Santa Cruz Biotechnology, Inc.), at RT with shaking for 90 minutes. Embryos were washed twice and then incubated with an additional primary antibody, rabbit polyclonal anti-H3K4me2 (1:100 dilution; Abcam, Inc.) or rabbit polyclonal anti-acH3K56 (1:100 dilution; Abcam, Inc.), in PVA-PBS overnight with shaking.
After the embryos were washed in PVA-PBS, they were incubated for 90 minutes with the second dye-conjugated secondary antibody, goat anti-rabbit IgG-TRITC (1:100 dilution). After washing embryos in PVA-PBS, they were dyed with DAPI for 5 minutes. The stained embryos were mounted on a glass slide and observed under a confocal fluorescence microscope (FV-1200; Olympus). The relative expression levels (H3K4me2, M3K9me2, acH3K9/K14, and acH3K56) of cloned embryos were analyzed by Olympus Fluoview (Olympus). A total of 15 embryos at the pronucleus stage (6 hours after activation) were analyzed per group. To measure the relative integrated signal, the ImageJ analysis program (Hartig, 2013) (version 1.52a; National Institutes of Health, Bethesda, MD) was used.
Statistical analysis
Data are expressed as means ± standard errors of the mean and analyzed using one-way analysis of variance in SPSS 18.0 (SPSS, Inc., Chicago, IL). Duncan's multiple range test was used to compare the groups. p < 0.05 was considered significant.
Results
Developmental comparison of a reconstructed cloned embryo with SCNT and CICT
The cloned embryo developmental efficiency was assessed through confirmation of the cleavage of cloned embryos at 24 hours and blastocyst developmental competence on day 3.5 of the culture period. The PN formation rate was not different between the CICT and SCNT groups (77.6% ± 2.2% vs. 77.9% ± 4.1%; Table 1). The percentages of two-cell embryos and blastocysts in the CICT group were significantly higher (p < 0.05) than that in the SCNT group (96.6% ± 1.1% vs. 86.7% ± 6.0%, 29.5% ± 2.6% vs. 22.1% ± 3.0%, respectively; Table 1). Moreover, the four- to eight-cell embryos had similar developmental rates in the CICT and SCNT groups (75.8% ± 3.2% vs. 68.1% ± 7.4%, respectively; Table 1).
In Vitro Development of Cloned Embryos Using Different Methods
Data are expressed as the mean ± SEM. Values with different superscripts (a,b) in the same column differ significantly (p < 0.05).
Cleavage and blastocyst rates were calculated based on the number of PN-formed oocytes for SCNT and CICT.
CICT, cytoplasm injection cloning technology; PN, pronucleus; SCNT, somatic cell nuclear transfer; SEM, standard error of the mean.
Analysis of the fluorescence level of cloned embryos between SCNT and CICT
We examined the epigenetic differences in cloned embryos between the CICT and SCNT groups. To investigate this, the cloned embryos were analyzed for histone methylation and acetylation using immunostaining with antibodies against acetylated histone H3 at lysine 9/14 and lysine 56 (acH3K9/K14 and acH3K56) and dimethylated histone H3 at lysine 4 and 9 (H3K4me2 and H3K9me2). The acetylation modification is shown in Figure 2B. The acetylation level of acH3K9/K14 in the CICT group was observed to be significantly higher (p < 0.05) than that of the SCNT group (Fig. 2A). Moreover, the acH3K56 relative levels were significantly higher (p < 0.05) in the CICT group than in the SCNT group (Fig. 2A). The methylation modification is shown in Figure 3B. The methylation levels of H3K4me2 and H3K9me2 were not different between the CICT and SCNT groups (Fig. 3A).

Global acetylation levels of acH3K9/K14 and acH3K56 in embryos.

Global methylation levels of H3K4me2 and H3K9me2 in embryos.
Quality of cloned blastocysts of SCNT and CICT
The numbers of total and apoptotic cells were measured in day 3.5 blastocysts (Fig. 4). The total cell number of blastocysts was not different between the CICT and SCNT groups (54.3 ± 3.6 vs. 53.8 ± 4.9; Table 2), but the number of apoptotic cells in blastocysts was significantly different (p < 0.05) between the CICT and SCNT groups (1.7 ± 0.2 vs. 2.9 ± 0.3; Table 2). Apoptotic cells were present in TE cells in both groups. In the SCNT group, the apoptotic cell number was increased in TE cells.
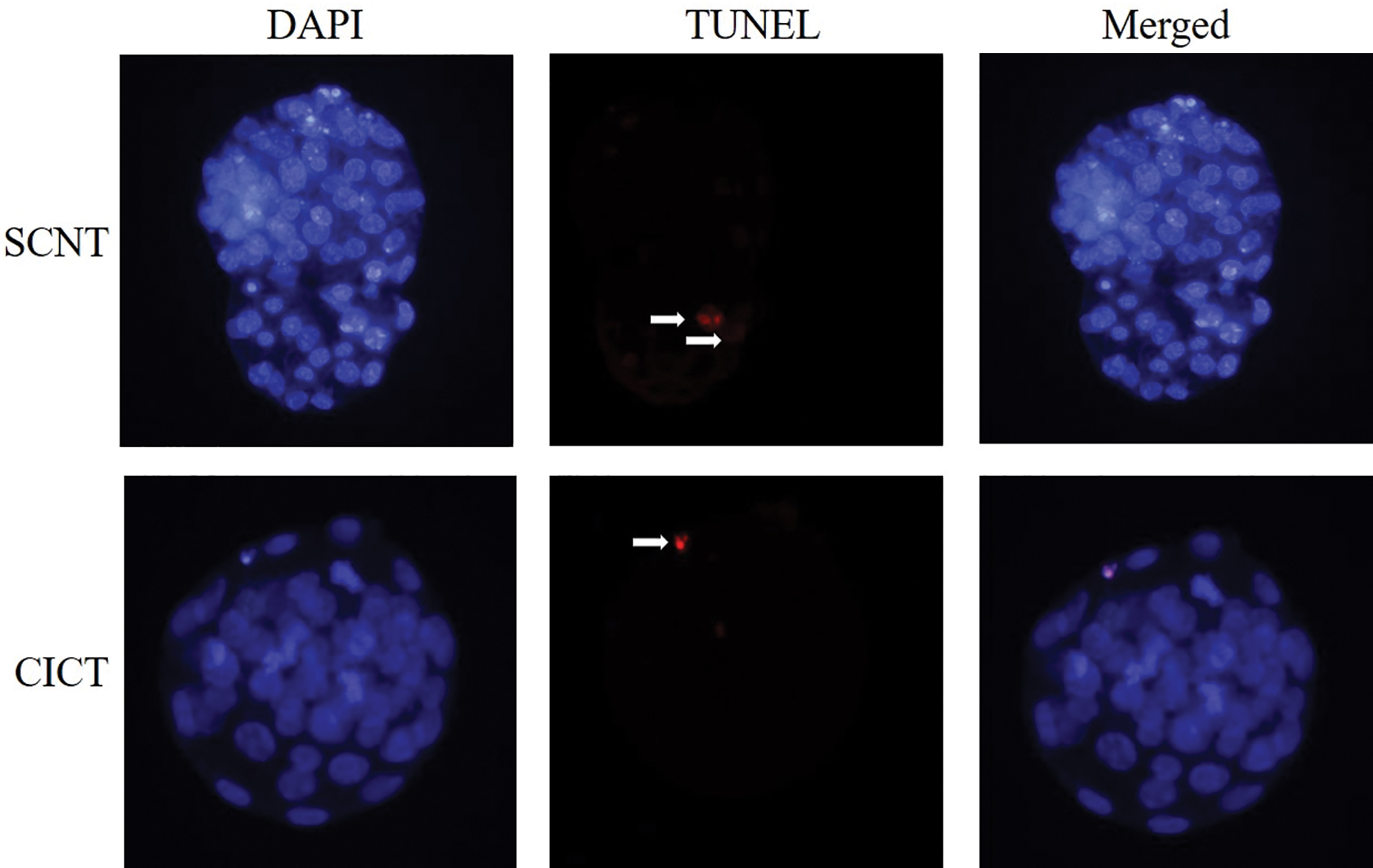
TUNEL assay of day 3.5 blastocysts in the SCNT and CICT groups. Apoptotic cells were detected by TUNEL assay (red). Each sample was counterstained with DAPI to visualize cell nuclei (blue). White arrows indicate apoptotic cells. DAPI, 4′,6-diamidino-2-phenylindole; TUNEL, terminal deoxynucleotidyl transferase dUTP nick-end labeling. Color images are available online.
Quality of Day 3.5 Blastocysts
Data are expressed as the mean ± SEM. Values with different superscripts (a,b) in the same column differ significantly (p < 0.05).
Discussion
A great deal of effort has been focused on animal cloning. However, traditional cloning methods such as SCNT have a problem, in that the cloning efficiency is still low since the birth of Dolly in 1996 (Campbell et al., 1996). Several techniques such as EA and HMC methods have been developed to increase the efficiency of cloning. However, EA and HMC are conducted in a zona-free condition. The zona-free embryos can easily transmit diseases and be damaged during handling (Verma et al., 2015).
Recently, the CICT method has been developed to overcome the drawbacks of traditional cloning methods (Xu et al., 2019). Using the CICT method, the cytoplasmic volume of enucleated oocytes is restored by donor-cytoplast oocytes. This protocol is conducted inside the zona pellucida, which provides protection against external environments, unlike EA and HMC. In feline species, the CICT method has been demonstrated to have enhanced embryo development and production of viable offspring over traditional cloning methods (Song et al., 2019).
To increase the mouse cloning efficiency, we have applied the CICT method to mice. The cloned embryo efficiency is related to gene expression, histone methylation, and acetylation. Moreover, cytoplasmic volume and donor cells positively affect embryonic development and nuclear reprogramming (Beyhan et al., 2007; Liu et al., 2018). In a previous study, the reconstructed cloned embryo, using CICT, showed that gene expression levels of DNMT 1 and DNMT 3a were significantly lower than in the traditional cloning method, indicating that CICT can improve epigenetic reprogramming. (Xu et al., 2019). In this study, we have evaluated the effect of the cytoplasmic volume on acetylation (acH3K9/14 and acH3K56) and apoptosis.
To evaluate the blastocyst quality, the numbers of total cells and apoptotic cells were counted (Gjorret et al., 2003; Maddox-Hyttel et al., 2003). Higher total cell numbers and lower number of apoptotic cells in blastocysts mean that there are high-quality blastocysts that could be implanted, resulting in a live birth (Balaban et al., 2000; Gardner et al., 2000). In this study, the apoptotic cell number was significantly lower in the CICT group than in the SCNT group. However, the total cell number was not significantly different between the SCNT and CICT groups. The present data indicate that the additional cytoplasm effect decreased the number of apoptotic cells, but not the total cell number.
The efficiency of cloning and the consequent embryo quality are related to the epigenetic status of the cloned embryo. The expression levels of histone methylation and acetylation on CICT cloned embryos were different from cloned embryos reconstructed with the traditional SCNT method. Histone methylation and acetylation occur on histone H3 lysine, such as acH3K9/K14, acH3K56, H3K4me2, and H3K9me2, which are related to nuclear remodeling and consequent reprogramming (Santos and Dean, 2004; Wang et al., 2007).
In this study, the fluorescence level of acH3K9/K14 was higher in CICT cloned embryos than that of traditional SCNT embryos. Moreover, the fluorescence level of acH3K56 was significantly higher in the CICT group than in the SCNT group. Histone acH3K9/K14 promotes DNA transcription (Adkins et al., 2007; Karmodiya et al., 2012; Wang et al., 2007), and acH3K56 is associated with genomic stability, DNA damage response, and the need to ensure cell divisions and changes in chromatin assembly pathways occurring in the embryo (Stejskal et al., 2015; Ziegler-Birling et al., 2016).
Therefore, it is possible that the higher levels of acH3K9/K14 and acH3K56 observed in CICT cloned embryos may be influencing the higher number of embryos developing to the blastocyst stage and their lower number of apoptotic cells. H3K9me2 is related to gene silencing, and H3K4me2 leads to initiation of gene transcription (Gupta et al., 2010; Lepikhov and Walter, 2004; Liu, et al., 2004; Rice et al., 2003; Shao et al., 2008).
In addition, nuclear reprogramming was affected by TSA. A previous study explains that TSA is a histone deacetylase (HDAC) inhibitor (Turner, 2000) and it plays a role in the factor of reprogramming during cloning and increases the efficiency of cloned embryo development (Azuma et al., 2018; Kishigami et al., 2006; Lee et al., 2011; Li et al., 2008). In this study, SCNT and CICT cloned embryos were exposed to TSA in the activation process. The fluorescence levels of acH3K9/K14 and H3K56 were higher in CICT cloned embryos than that of SCNT. This result shows that additional cytoplasm affects nuclear reprogramming.
In conclusion, the results of this study indicate that using the CICT method resulted in a reconstructed embryo, restoration of cytoplasmic volume, and enhancement of the developmental competence and quality of cloned embryos. These results may assist with cloning technology and improve the reprogramming efficiency of cloned embryos.
Ethics Approval and Consent to Participate
The Gyeongsang National University Institute of Animal Care and Use Committee (GNU-161228-M0075), Gyeongsang National University, Republic of Korea, approved this study, which was conducted at the Animal Research Facility and Veterinary School of Gyeongsang National University, Republic of Korea.
Footnotes
Authors' Contributions
All authors contributed to this article. S.-H.S., S.-H.O., and L.X. designed and performed the experiments. K.-L.L., J.-Y.H., and M.-D.J. analyzed the data and assisted with culture medium preparation. I.-K.K. wrote the manuscript and managed this work.
Author Disclosure Statement
The authors declare they have no competing financial interests.
Funding Information
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIT) (No. 2020R1A2C200661). S.-H.S., S.-H.O., L.X., J.-Y.H., and M.-D.J. were supported by scholarships from the BK21 Plus Program.