
Abstract
This study aimed to investigate the mechanism of miR-142-5p and Yin Yang 1 (YY1) on regulating epithelial–mesenchymal transition (EMT) in lung cancer cell metastasis. The expressions of YY1 and miR-142-5p in different lung cancer cell lines were negatively correlated. The results of the dual-luciferase reporter assay further validated that miR-142-5p directly targeted YY1. Subsequently, transwell assays, wound-healing assay, and transplantation tumor model in nude mice proved that YY1 could promote the metastasis of lung cancer cells, whereas miR-142-5p impaired the stimulating effect of YY1 on the metastasis ability of lung cancer cells in vitro and in vivo. Western blot and quantitative real-time polymerase chain reaction analysis of the EMT-related proteins indicated that YY1 could enhance the metastasis ability of lung cancer cells by promoting EMT. On the contrary, miR-142-5p constrained the expression of mesenchymal markers by targeting YY1, reversed the differentiation of cells into mesenchymal cells, and weakened the metastasis ability of tumor cells in vitro and in vivo. In summary, miR-142-5p may regulate the expressions of EMT-related proteins by targeting YY1, thereby inhibiting lung cancer metastasis, which provides a promising therapeutic target for lung cancer.
Introduction
Primary bronchogenic carcinoma (PBC) is a common tumor with increasing mortality and morbidity worldwide. According to clinical pathological morphology, PBC is divided into small cell lung cancer (SCLC) and nonsmall cell lung cancer (NSCLC) (Saber et al., 2015), among which NSCLC accounts for 80% of the total incidence of lung cancer (Jin et al., 2018). In recent years, the invasion and metastasis of NSCLC have attached more and more attention. Compared with SCLC, the diffusion and metastasis of NSCLC are relatively slow. However, most patients have developed viscera metastasis at the time of diagnosis and are prone to irreversible metastasis to bone and central nervous system (Hendriks et al., 2014; Kilvaer et al., 2016).
At present, the traditional treatments for lung cancer, namely surgery, chemotherapy, and radiotherapy, have unsatisfactory therapeutic effects such as distant metastasis and tumor recurrence, leading to poor prognosis and reduction of 5-year survival rate (Marcus and Zhou, 2010; Seok et al., 2015). Therefore, identifying a novel molecular targeted therapy for lung cancer is urgently required.
As a member of the GLI-Krüppel family of transcription factors, multifunctional protein Yin Yang 1 (YY1) can suppress or activate gene expression according to cellular context (He and Casaccia-Bonnefil, 2010; Khachigian, 2018; Kwon et al., 2018). Several studies have shown that the abnormal expression of YY1 plays a role in the regulation of tumor proliferation and metastasis through interaction with different protein cofactors, including NSCLC (Huang et al., 2017; Khachigian, 2018). However, the mechanism of action of YY1 in NSCLC remains unclear.
Dysregulation of gene expression is considered a vital factor in tumorigenesis (Decai et al., 2018). MicroRNAs (miRNAs), as noncoding small RNA molecules with endogenous gene expression regulation, can bind to target gene mRNAs to regulate the target gene expression at the post-transcriptional level and further participate in various disease processes, including a variety of miRNAs such as miR-21, miR-574-5p, miR-34a, miR-1254, and miR-15a/16, which have been reported to play key regulatory roles in the development of NSCLC (Bandi and Vassella, 2011; Foss et al., 2011; Wei et al., 2011).
As an emerging miRNA in recent years, miR-142-5p has been found to be involved in the formation of atherosclerotic plaque, the occurrence of colorectal cancer, and the proliferation and differentiation of stem cells (Islam et al., 2018; Yang et al., 2018). A research conducted by Wang et al. (2017) has clarified that miR- 142-5p is less expressed in NSCLC patients and can inhibit the development of NSCLC by targeting PIK3CA. Nevertheless, whether it participates in the metastasis of NSCLC and its possible mechanism of action has not been studied. In this study, we found that miR-142-5p had a binding site to YY1 through online miRNA targeting prediction method such as TargetScan, and further explored their roles in NSCLC cells and their possible mechanisms, providing theoretical basis for new molecules markers of NSCLC.
Materials and Methods
Materials
Human bronchial epithelial cells (BEAS-2B), human lung cancer cell lines (H1299, A549, L78, H460, H1650, H3122, and PGCL3) were purchased from the Cell Resource Center of Shanghai Research Center of Life Sciences, Chinese Academy of Sciences.
Fetal bovine serum (FBS), phosphate-buffered saline (PBS), Trypsin, RPMI-1640 medium, and DMEM (high glucose) were purchased from Hyclone. Anti-YY1, anti-Snail 1, anti-E-cadherin, anti-ZO-1, anti-N-cadherin, and HRP Goat anti-Rabbit IgG were purchased from Abcam; YY1 overexpression vector and negative control (NC), anti-miR-142-5p, miR-142-5p mimic, and miR-142-5p disorder control (NC mimic) were synthesized by Shanghai GenePharma Company. The pGL3 luciferase reporter vector was purchased from Promega, and the corresponding pGL3-PIK3CA-3′UTR and mutation reporter were constructed and synthesized by Shanghai Sangon Biotech Co., Ltd, as well as the design and synthetization of polymerase chain reaction (PCR) primers for specific detection (Table 1).
Primer Sequences
Cell culture and transfection
The BEAS-2B, H1299, A549, L78, H460, H1650, H3122, and PGCL3 cell lines were cultured into T25 flasks supplemented with 10% FBS (complete medium), and incubated at 37°C with 5% CO2 and at constant temperature and humidity. The solution was changed once every other day, and the cell density was fused to 90% under a light microscope before digestion and passage. The cells were cultured to logarithmic growth phase for subsequent experiments.
A549 cells were seeded into 24-well plates (2 × 104 cells/well) and divided into NC, YY1 (YY1 overexpression), and miR-142-5p mimic groups. Besides, H460 cells were also seeded into 24-well plates (2 × 104 cells/well) and divided into NC and anti-miR-142-5p groups. Then A549 and H460 cells were transfected with different plasmids using Lipofectamine™ 3000 kit (Invitrogen) according to manufacturer instructions. After 48 hours, the stably transfected cells were screened for YY1 and miR-142-5p expression, and then they were used in subsequent experiments.
Dual-luciferase reporter assay
According to TargetScan (www.targetscan.org) and MiRanda (www.microrna.org/microrna/home.do) databases, the wild-type YY1 3′UTR (YY1-3′UTR-WT) fragment containing the predicted miR-142-5p binding sites was amplified using the primers: forward, 5′-ATCCTAGCCCTTACTGAACATTC-3′; and reverse, 5′-CTAAAGAAGTAACCTTGGAGGCA-3′. Meanwhile, the mutant YY1 3′UTR (YY1-3′UTR-MUT) was obtained by mutating the core sequence (GCCCT) in YY1 3′UTR region. Subsequently, the YY1-3′UTR-WT or YY1-3′UTR-MUT was constructed into the pGL3 luciferase reporter vector, namely pGL3-PIK3CA-YY1 WT and pGL3-PIK3CA-YY1 MUT.
The above luciferase reporter plasmid was cotransfected with miR-142-5p mimic or NC mimic into HEK293T cells, and the pRL-TK luciferase reporter vector was used as NC. After transfection for 48 hours, dual-luciferase reporter assay system (Promega) was used to detect and calculate the relative luciferase activity of cells in each group to verify the correlation of miR-142-5p with YY1.
Transwell assay
The cells in logarithmic growth phase were collected after transfection. The digested cells were washed once with PBS and resuspended in serum-free RPMI-1640 medium to a final density of 2 × 105/mL. After blowing and mixing, a 200 μL of cell suspension was evenly added into 24-well Transwell chamber (Costor), and 600 μL of complete medium was added to the lower chamber. The cell culture plate was transferred to an incubator for routine culture. After 60 hours, the culture plate was removed, the remaining medium was aspirated, and the inner wall of the chamber was washed twice with PBS. Then, 600 μL of precooled 70% alcohol were added into the lower chamber to fix cells for 30 minutes, and the fixing solution was discarded. Next, 600 μL of 0.1% crystal violet staining solution was added at room temperature without light for 10 minutes, and the staining solution was discarded.
The number of cells was counted in five random selected fields under a microscope. The experiment was repeated three times, and the mean value was recorded as the number of cell migration.
The experimental procedures of transwell assay were consistent with the cell migration experiment, except for a procedure as follows: 4 hours before the cell invasion assay, 10 mg/mL Matrigel gel (Millipore) was diluted with empty medium at a ratio of 1:3, and then 20 μL Matrigel was evenly spread on the chamber membrane and stored at 37°C for future use.
Wound-healing assay
The cells in logarithmic growth phase were placed in a 96-well plate so as to 80% confluence. Subsequently, 10 μL of micropipette tip was used to create a scratch along the midline of the well. After this, the original medium in the well was aspirated, washed slowly with PBS once, and cultured in 200 μL of low serum medium containing only 2% FBS. The cell migration was observed under light microscope and the scratches were calculated at 0, 12, and 24 hours.
Transplantation tumor model in nude mice
All mouse experiments were approved by the Ethics Committee of the Tumor Hospital Affiliated to Xinjiang Medical University. Eighteen BALB/c nude mice (female, 4–5 weeks, 20.00 ± 2.25 g) were routinely housed in SPF animal rooms, and all animals were handled according to the guidelines of the Tumor Hospital Affiliated to Xinjiang Medical University. Nude mice were randomly divided into NC group, YY1 group, and YY1+miR-142-5p mimic group (n = 6 per group).
The cells of each group were collected and made into single-cell suspension with a concentration of 5 × 107 cells/mL, which were filtered through a 70 μm sterile mesh to avoid cell adhesion. Next, mice were fixed and their tails were sterilized with 75% alcohol. After the venous blood vessels were filled, 0.1 mL of filtered cell suspension was given to each mouse along the tail vein. NightOWL II LB983 small animal living imaging system was used to observe the metastasis and tumorigenesis of the transplanted tumors in vivo, and photographed. After 8 weeks, the mice were killed by cervical dislocation, and all the organs were stained with hematoxylin and eosin (H&E) to observe the distant metastasis of the tumor.
Real-time quantitative PCR
After digestive centrifugation of stably transfected cells (4 × 106) in each group, the total RNAs were extracted from the cell pellet by 1 mL of TRIzol reagent (Invitrogen), and reverse-transcribed to corresponding cDNAs. Next, the concentration and purity of cDNAs were measured using Nanodrop nucleic acid quantitation, and the targeted gene expression was quantitatively detected in accordance with Power Up SYBR Green Kit instruction (Invitrogen). GAPDH and U6 were used as the internal reference genes of mRNA and miRNA, respectively.
PCR system (reaction mixture 20 μL) was set as follows: up to 40 cycles consisting of predenaturation at 95°C for 5 minutes; amplification at 95°C for 1 minute, annealing at 55°C for 2 minutes, and extension at 72°C for 1 minute. Three subwells were set in each group of cell samples, and the 2−ΔΔCt method was used to calculate relative expression of the target gene using data collected from mean fluorescence signal value (Ct value). At the same time, a negative control was used to exclude PCR contamination and primer–dimer interference in the reaction system.
Western blot analysis
Single-layer adherent cells were collected, washed twice with precooled PBS, and pelleted by centrifugation. Then, 1 mL of RIPA lysate (Lianke Bio) containing 1% phosphatase inhibitor was added into the cell pellet to lyse cells for 30 minutes in an ice bath, and the cell pellet were resuspended by blowing with a pipette every 10 minutes. After centrifugation at 12,000 rpm for 15 minutes at 4°C, the supernatant of the lysate was collected and transferred to a new 1.5 mL EP tube, and the protein concentration in each group was determined using BCA Protein Quantification Kit (Lianke Bio). The same amount of protein was added to an equal volume of 1 × SDS (sodium dodecyl sulfate) loading buffer, and was denatured by heating at 100°C for 10 minutes in a water bath.
Next, proteins were separated by 10% SDS-PAGE under 100 V constant pressure electrophoresis for 120 minutes and transferred onto PVDF (polyvinylidene difluoride) membranes (Millipore). The corresponding molecular weight band antibody was incubated, followed by the addition of luminescent solution. Finally, Bio-Rad CheciDOC XRS chemiluminescence imaging system was performed to detect the bands, and the luminescence images were acquired for quantitative analysis.
Statistical analysis
SPSS 13.0 software was used for statistical analysis. Comparison between different experimental groups was performed with one-way analysis of variance (the one-way ANOVA). The expression levels of candidate miRNA in different cells were investigated using real-time fluorescence quantitative analysis. All experiments were performed at least in triplicate and the data were presented as the means ± standard deviations. A value of p < 0.05 was considered statistically significant.
Results
The correlation between the expression of miR-142-5p and YY1 in different NSCLC cell lines
To find out whether YY1 was a target of miR-142-5p, the expression fold changes of YY1 mRNA and miR-142-5p in various lung cancer cell lines (H1299, A549, L78, H460, H1650, H3122, and PGCL3) and human bronchial epithelial cells (BEAS-2B) were detected by quantitative real-time PCR (qRT-PCR). As given in Figure 1A, YY1 was highly expressed in all lung cancer-related cell lines when compared with BEAS-2B cell lines, with the largest change in expression multiples in A549 cell lines and the lowest expression in H460 cell lines. This finding indicated that high expression of YY1 might play a role in promoting tumor genesis and metastasis. Besides, miR-142-5p showed low expression in five kinds of lung cancer cells except L78 and H460, and the expression level was the lowest in A549 cells (Fig. 1B). Therefore, A549 was selected for subsequent studies.
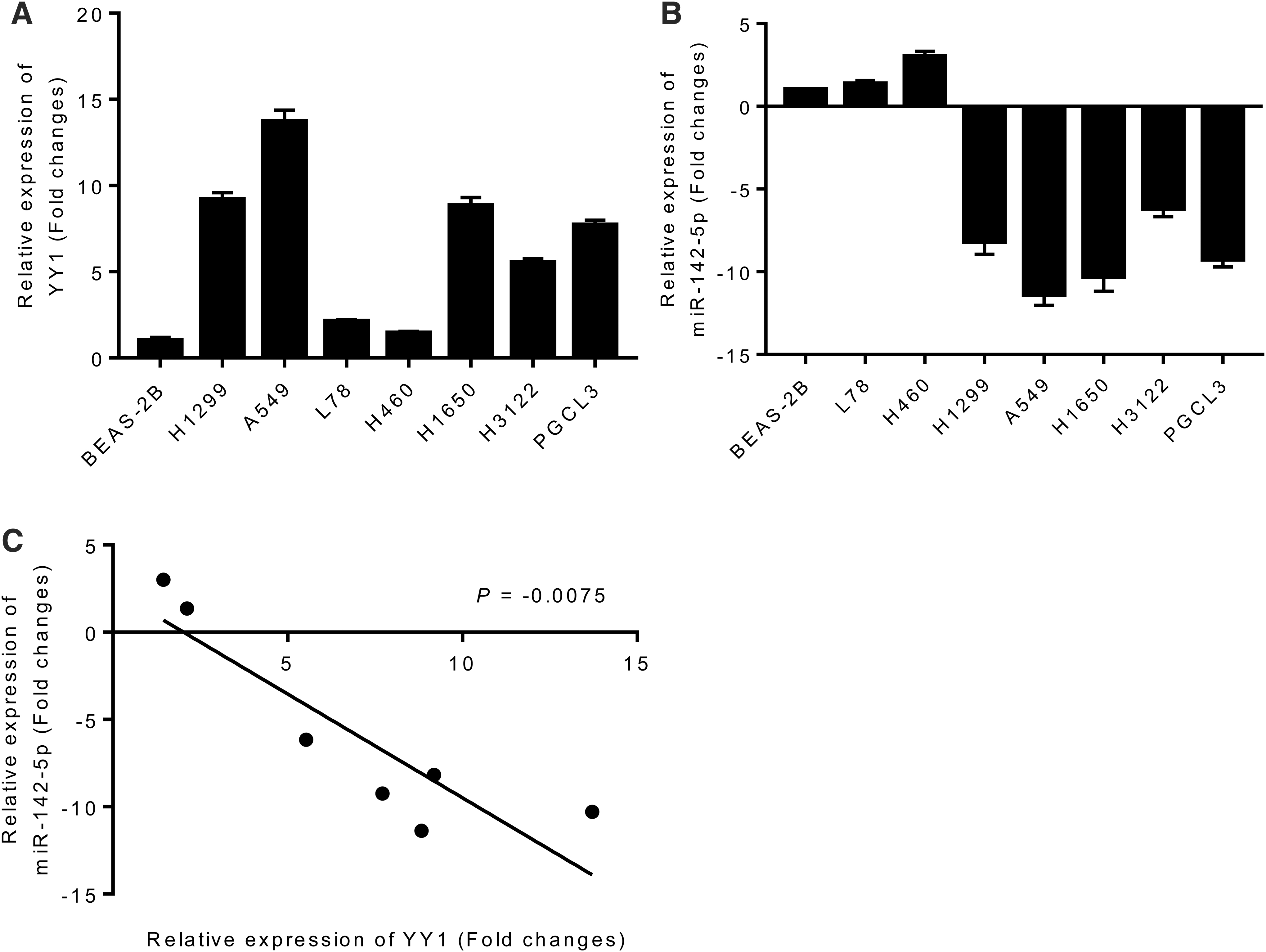
The expression of YY1 mRNA
Then, we analyzed the correlation between the expression of YY1 and miR-142-5p in different lung cancer cell lines. From Figure 1C, the expression levels of YY1 and miR-142-5p in different lung cancer cell lines were negatively correlated (p = 0.0075).
MiR-142-5p directly targeted YY1
Based on the results of miRNA seed sequence targeting prediction, a potential miR-142-5p binding site in the 3′-untranslated region (3′UTR) of YY1 was observed (Fig. 2A). Therefore, pGL3-PIK3CA-YY1 WT or pGL3-PIK3CA-YY1 MUT was constructed into the luciferase gene plasmids. The expression plasmid was then cotransfected into the A549 cells with miR-142-5p mimic or NC mimic. The dual-luciferase reporter assay system showed that the luciferase activity was significantly decreased in pGL3-PIK3CA-YY1 WT by miR-142-5p mimic, whereas it remained unchanged in pRL-TK and pGL3-PIK3CA-YY1 MUT (p < 0.01), indicating that miR-142-5p had a targeted binding site to YY1 (Fig. 2B).
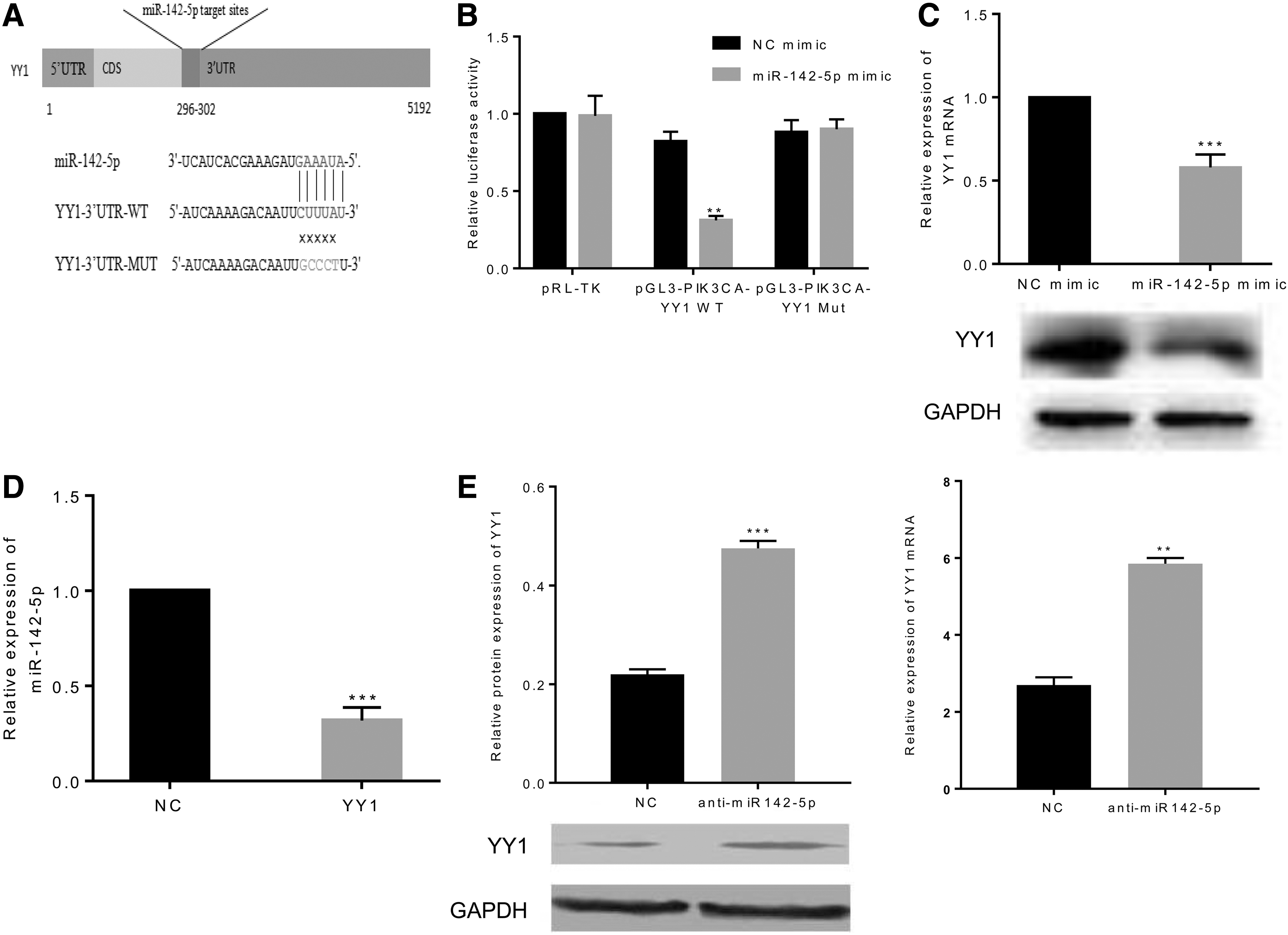
MiR-142-5p directly targeted YY1.
To further investigate the relationship between YY1 and miR-142-5p in A549 cells with high expression of YY1, qRT-PCR and western blot were performed to detect the effect of miR-142-5p on YY1 both at mRNA and protein level. Results suggested that transfection with miR-142-5p mimic downregulated the expression of YY1 at both protein and mRNA levels (p < 0.001) (Fig. 2C). Similarly, the overexpression of YY1 significantly suppressed the relative expression of miR-142-5p in A549 cells (p < 0.001) (Fig. 2D). In addition, in the H460 cells with low expression of YY1, the inhibition of miR-142-5p expression significantly improved protein and gene expression of YY1 (p < 0.001 and p < 0.01, respectively) (Fig. 2E).
MiR-142-5p inhibited cell migration and invasion in vitro by targeting YY1
To identify the effects of miR-142-5p on YY1 regulation on migration and invasion abilities of A549 cells in vitro, transwell assays and wound-healing assay were performed. As given in Figure 3, YY1 overexpression significantly facilitated the invasion, migration, and wound-healing rate of A549 cells (p < 0.01 for migration and invasion assays and p < 0.001 for wound-healing assay), whereas miR-142-5p overexpression had the opposite effect (all p < 0.01). Besides, compared with YY1 group, YY1+miR-142-5p mimic significantly reversed the role of YY1 in promoting the invasion ability of A549 cells (p < 0.05), indicating that miR-142-5p might be a regulator of YY1.

MiR-142-5p inhibited cell migration and invasion in vitro by targeting YY1.
MiR-142-5p regulates NSCLC metastasis in vivo by targeting YY1
To further study the effects of miR-142-5p on YY1 regulation on the migration and invasion of A549 cells in vivo, BALB/c nude mice were used in in vivo experiments. The results showed that after 4 weeks of tumor implantation, significant fluorescence aggregation was observed in liver of YY1 group, and there was significant difference between NC group and YY1+miR-142-5p mimic group (p < 0.01). Eight weeks later, significant metastasis appeared in the liver of the mice. Nevertheless, cotransfection of miR-142-5p mimic significantly attenuated the fluorescence intensity of metastatic lesions in the liver tissues of mice (Fig. 4A–B).

MiR-142-5p regulated metastasis of NSCLC in vivo by targeting YY1.
Then, histopathological changes were noticed by H&E staining in isolated liver tissue sections. As given in Figure 4C, the liver of mice in YY1 group showed severe lesions, with rupture of liver tissue cells and a large amount of inflammatory infiltration in the intercellular space. Of note, the tissue lesions in YY1+miR-142-5p mimic group were alleviated compared with that of YY1 group.
MiR-142-5p suppresses epithelial–mesenchymal transition-related protein expression by targeting YY1
Western blot and qRT-PCR were adopted to determine epithelial–mesenchymal transition (EMT)-related protein and mRNA expressions. YY1 overexpression significantly promoted the expressions of intermesenchymal markers N-cadherin and Snail 1 at protein and mRNA level in A549 cells, and inhibited the protein and mRNA expression levels of epithelial markers, E-cadherin and ZO-1 (p < 0.001). On the contrary, miR-142-5p mimic overexpression remarkably downregulated N-cadherin and Snail 1 expression at protein and mRNA level and upregulated E-cadherin and ZO-1 protein and mRNA expression (p < 0.001).
Moreover, cotransfected miR-142-5p mimic reversed the promotion effect of YY1 on N-cadherin and Snail 1, as well as the inhibitory function of YY1 on E-cadherin and ZO-1. There was no significant difference between YY1+miR-142-5p mimic group and NC group (Fig. 5).

YY1 promoted the EMT markers in A549 cells, whereas miR-142-5p reversed the function of YY1.
Discussion
The most notable finding of this study was that miR-142-5p can regulate the expression of EMT-related proteins by targeting YY1, thereby inhibiting the metastasis of the lung cancer in vitro and in vivo. This finding fills the gap in the impact of miR-142-5p on lung cancer. YY1 gene is located in the telomere region of human chromosome 14, which was first discovered by Shi et al. (1991). It can play a transcriptional activation part as a transcription factor by binding to adenovirus P5 promoter region, promoting the activation of adenovirus E1A. In addition, YY1 has been reported to play a dual role of transcriptional suppression or activation in a variety of diseases and biological processes by recruiting different protein factors (Liang et al., 2012).
In recent years, the role of YY1 in the development of tumors has been widely concerned. In addition to participating in biological processes such as cell proliferation regulation and apoptosis, YY1 can also simultaneously perform regulatory function on transcription levels including methylation, acetylation, and ubiquitination of target genes (Qu et al., 2017; Zhou et al., 2018). Moreover, YY1 has been widely reported to promote cancer in prostate cancer (Seligson et al., 2005), colon cancer (Yokoyama et al., 2010), ovarian cancer (Xia et al., 2016), and breast cancer (Wang et al., 2016). Although YY1 has been studied to be involved in the occurrence and development of lung cancer, the mechanism of its action is relatively rare.
In this study, the expression fold changes of YY1 and miR-142-5p genes in 7 human NSCLC cell lines and human normal lung epithelial cells were compared using qRT-PCR to analyze their correlation. Results showed that YY1 was highly expressed in 7 lung cancer cells to different degrees, whereas miR-142-5p was lowly expressed in 5 lung cancer cell lines, indicating that the expression levels of YY1 and miR-142-5p in different lung cancer cell lines were negatively correlated.
Previous studies have shown that miRNAs may play their biological functions by inhibiting target gene expression (Mohsen et al., 2013). Therefore, bioinformatics prediction method was performed to find possible target binding sequences of YY1 and miR-142-5p, suggesting that YY1 and miR-142-5p may exert their interaction mechanism in lung cancer cells through targeted binding. Dual-luciferase reporter assay results indicated that miR-142-5p mimic significantly inhibited pGL3-PIK3CA-YY1 WT luciferase activity, whereas it had no effect on the luciferase activity of pGL3-PIK3CA-YY1 MUT without the binding site of miR-142-5p after mutation. Meanwhile, overexpression of intracellular miR-142-5p or YY1 could inhibit the expression of YY1 and miR-142-5p, respectively, further verifying the regulatory effect of miR-142-5p on YY1.
Several studies have demonstrated that EMT of tumor cells induces the enhancement of tumor cells metastasis in vivo and in vitro (Yang and Weinberg, 2008). In this study, overexpression of YY1 remarkedly augmented the protein and mRNA expression of EMT-associated genes, N-cadherin and Snail 1 in A549 cells, whereas it suppressed the protein and mRNA expression of epithelial markers E-cadherin and ZO-1. Intracellular Snail 1 often inhibits E-cadherin expression by binding to E-cadherin promoter. Taken together, YY1 could enhance the metastasis ability of lung cancer cells in vitro and in vivo by promoting EMT. Conversely, miR-142-5p could constrain the expression of mesenchymal markers by targeting YY1, reverse the differentiation of cells into mesenchymal cells, and weaken the metastasis ability of tumor cells in vitro and in vivo.
In addition to the induction of EMT to promote the metastasis of tumor cells into liver tissue, brain metastasis and bone metastasis of NSCLC patients are also common clinical sites of advanced lung cancer metastasis (Song et al., 2019). This study only observed the formation of metastatic nodules in liver tissues after 8 weeks of establishment of transplantation tumor model in nude mice, but no tumor formation occurred in the brain or bone tissue regions. It was speculated that it might be related to the duration of model modeling in vivo, and whether miR-142-5p can regulate brain metastasis and bone metastasis of advanced lung cancer by targeting YY1 remains to be further explored.
In addition, the occurrence of bone metastases in lung cancer patients has been found to be associated with the abnormal expression of multiple circulating miRNAs (Lei and Dian-Ming, 2017; Song et al., 2019), and plasma miR-142-5p has been reported to be of great significance in the early diagnosis of colorectal cancer in previous study (Ghanbari et al., 2015), suggesting that we can further explore the role and significance of YY1 and miR-142-5p in bone metastasis and early diagnosis of NSCLC.
Conclusion
In summary, this study reveals that YY1 is highly expressed in NSCLC cells, which is reversed by miR-142-5p by targeting YY1, and eliminates the promotion effect of YY1 on lung cancer cell metastasis by promoting EMT in vivo and in vitro. These findings indicate that miR-142-5p is a novel therapeutic target for lung cancer.
Footnotes
Author Disclosure Statement
The authors declare they have no competing financial interests.
Funding information
This study was supported by the National Natural Science Foundation Project (81760014), Xinjiang Uygur Autonomous Region Municipal Natural Science Foundation (2020D01C207), and Youth Medical Science and Technology Talents Special Scientific Research Project of the Health and Family Planning Commission in Xinjiang Uygur Autonomous Region (WJWY-201907).