
Abstract
Wnt/β-Catenin signaling pathway plays an important role in maintaining self-renewal and pluripotency of human and mouse embryonic stem cells (ESCs). Activation of Wnt/β-Catenin signaling pathway by glycogen synthase kinase-3 (GSK3) inhibitor, the Wnt signaling agonist, could maintain the pluripotency of human and mouse ESCs in the presence of serum. However, the role of signaling pathway in the derivation of buffalo ESCs remains unclear. In this study, we used GSK3 inhibitors (6-bromoindirubin-3′-oxime [BIO] and CHIR99021) and investigated the effect of Wnt/β-Catenin activation on colony formation, proliferation, self-renewal, and pluripotency of Chinese swamp buffalo (buffalo) embryonic stem cell-like cells (ES-like cells), which were isolated from blastocysts. The results showed that buffalo ES-like cells displayed typical morphological characteristics of pluripotent stem cells: positive for alkaline phosphatase staining, expression of pluripotent markers, including OCT4, SOX2, SSEA-1, SSEA-4, LIN28, CH1, NANOG, and the proliferative markers, PCNA and C-MYC. Furthermore, activation of Wnt/β-Catenin signaling pathway by GSK3 inhibitors could promote colony formation and proliferation of buffalo ES-like cells and maintain their undifferentiated state, and upregulate the expression levels of pluripotent-related genes and proliferation-related genes. These results indicated that Wnt/β-Catenin signaling pathway plays an important role in the derivation and pluripotency of buffalo ES-like cells.
Introduction
Chinese swamp buffalo is an important domestic animal, most distributed in the south of China. It is very popular for its nutrient-rich milk which is richer in fat, protein, calcium, and vitamin compared with dairy cow. However, milk yield of buffalo is low which cannot meet the needs of people. Buffalo pluripotent stem cell-based gene modification was considered to be an efficient way to breed high-milk yield buffalo.
Embryonic stem cells (ESCs) are derived from the inner cell mass (ICM) of early embryo at the blastocyst stage, and possess the ability of self-renewal and maintenance pluripotency. Embryonic stem cell-like cells (ES-like cells) were generated from a large number of mammalian species, including many farm animals, such as pig (Notarianni et al., 1990), sheep (Notarianni et al., 1991), rabbit (Graves and Moreadith, 1993), bovine (First et al., 1994), and equine (Saito et al., 2002). Buffalo ES-like cells have been established in our and other laboratories (Huang et al., 2010; Sharma et al., 2011; Verma et al., 2007; Zandi et al., 2014, 2015).
In these studies, leukemia inhibitory factor (LIF) and basic fibroblast growth factor (bFGF) were considered to be important for pluripotency of buffalo ES-like cells. But no bona fide buffalo ESC lines had been established (just called ES-like cells), because the mechanism underlying pluripotency maintenance in buffalo ESCs remains not clear. Meanwhile, the culture conditions, especially the chemical supplements maintaining pluripotency and self-renewal of buffalo ESCs, need further investigations.
Different intracellular signaling pathways and transcription factors have different roles in pluripotency maintenance of different mammal ESCs. For example, LIF can be used to maintain pluripotency of 129 mouse ES cells on gelatin in the absence of feeders, but not for non-129 mouse ESCs (Sato et al., 2009). Studies also demonstrated that LIF treatment has no or little effect on maintaining self-renewal of human and rabbit ESCs (Honda et al., 2009; Sato et al., 2003; Thomson et al., 1998). Besides, Wnt/β-Catenin is another important signaling pathway in maintaining self-renewal of pluripotent stem cells. Activation of Wnt signaling pathway was sufficient to maintain self-renewal of both human and mouse ES cells with glycogen synthase kinase-3 (GSK3) inhibitor: 6-bromoindirubin-3′-oxime (BIO) (Sato et al., 2003).
GSK3, a serine/threonine kinase, existing in two isoforms (named GSK3α and GSK3β), is a key regulator of numerous signaling pathways, especially Wnt pathways (de Jaime-Soguero et al., 2018). The inhibition of GSK3 results in dephosphorylation of β-Catenin and leads to an accumulation of β-Catenin in the nucleus to activate the canonical Wnt signaling pathway (Caspi et al., 2008).
GSK3 inhibitors BIO or β-Catenin inhibitors IWR1 combining with stem cell factor (SCF), LIF, and bFGF could significantly contribute to the establishment of porcine and bovine ESCs, and maintain their undifferentiated status (Bogliotti et al., 2018; Li et al., 2020; Wen et al., 2010; Xu et al., 2019).
CHIR99021, another GSK3 inhibitor, also promoted self-renewal of ESCs in C57BL/6 (B6) and BALB/c mouse strains by stabilizing of β-Catenin protein level (Cooney et al., 2012), enhanced KLF4 expression through β-Catenin signaling and miR-7α regulation in J1 mouse ESCs (Ai et al., 2016), and sustained ovine ES-like cells in semidefined medium without feeder cells (Zhao et al., 2011). Zandi et al. (2015) used BIO combining with WNT3A to activate WNT signaling pathway that upregulated the expression of pluripotent-related genes in buffalo ES-like cells. Indicating the Wnt/β-Catenin pathway influences the pluripotency of buffalo ES-like cells. However, the effect of Wnt/β-Catenin activation on primary colony formation and proliferation of buffalo ES-like cells is still not very clear.
In this study, the GSK3 inhibitors, BIO and CHIR99021, were used to culture the buffalo ES-like cells, to explore the effects of Wnt/β-Catenin signaling pathway on the derivation, proliferation, and maintenance of pluripotency in buffalo ES-like cells. These efforts will provide a chemical screening platform to optimize the culture condition for establishing buffalo ES cell lines.
Materials and Methods
Reagents and medium
Cell and embryo culture medium with supplements were obtained from Gibco (Carlsbad, CA). Cell and embryo culture dishes were obtained from Corning (Steuben County, NY). The primary antibodies of cell surface antigens SSEA-1, SSEA-4; pluripotency markers Oct4, Sox2, and β-Catenin; and the secondary antibodies were obtained from Cell Signaling Technology (Danvers, MA). Reverse transcription–polymerase chain reaction (RT-PCR) and quantitative real-time PCR (qRT-PCR) reagents were obtained from TaKaRa Bio, Inc. (Kusatsu, Japan). The GSK3 inhibitor (BIO: Catalog No. 361550, and CHIR99021: Catalog No. 361571), the cytokines (LIF, bFGF, and SCF), and other chemical reagents were obtained from Sigma-Aldrich (St, Louis, MO), unless otherwise indicated.
Preparations of buffalo blastocysts
All experiments about animals were performed in State Key Laboratory for Conservation and Utilization of Subtropical Agro-Bioresources, and were conducted in accordance with its guidelines for the care and use of laboratory animals. Buffalo ovaries were collected from local slaughterhouse and transported to the laboratory within 3 hours, washed three times with 0.9% sterile saline at 37°C. Cumulus–oocyte complexes (COCs) were aspirated in the washing medium (9.5 g TCM199, 1.2 g Hepes, 0.06 g Penicillin, 0.9 g NaCl, 0.1 g Streptomycin, 20 mL newborn calf serum [NCS], 1 L H2O) using 10-mL injector with pinhead and washed twice. COCs were cultured in the maturation medium (9.5 g TCM199, 26.2 mmol/L NaHCO3, 5.0 mmol/L Hepes, 5% fetal bovine serum [FBS], 0.1 μg/mL FSH, 0.1 g/L Streptomycin, 0.06 g/L Penicillin) under a humidified atmosphere of 5% CO2 in air at 38.5°C for 24 hours.
In vitro-matured oocytes were placed in 25 μL droplet of fertilization medium (modified Tyrode's medium without lactate, 50 μg/mL heparin, 2.5 mmol/L caffeine). The frozen semen straw was stayed 10 seconds in the air, and gently shook for 10 seconds in a 37°C water bath. The spermatozoa were suspended in 1.5 mL fertilization medium for 30 minutes, and the superstratum liquid was taken out for washing by centrifugation. Processed spermatozoa and in vitro oocytes were incubated in a 25 μL droplet of fertilization medium covered with mineral oil for 24 hours in the same conditions as before described. After 24 hours of incubating, the presumptive zygotes were transferred to 25 μL droplets of CM (TCM199, 26.2 mmol/L NaHCO3, 5.0 mmol/L Hepes, 3% FBS, 0.1 g/L Streptomycin, and 0.06 g/L Penicillin), and cultured to blastocyst stage.
Isolation and culture of ES-like cells
First, the zona pellucida of the embryo was removed by the pinhead of the 1-mL injector under a microscope, ICMs of the blastocyst and hatched blastocyst were isolated in the same way, and seeded on buffalo embryonic fibroblast feeder layers, which were treated with mitomycin-C in a four-well multidish. After 5–6 days of culture, the buffalo ICM-derived primary ES-like colonies were passaged by mechanical dissociation. The detailed isolation and subculture protocols could refer to our previous report (Huang et al., 2010). The ESC culture medium (ECM) was composed of high-glucose Dulbecco's modified Eagle's medium, 20% knockout serum replacement (KSR), 0.1 mmol/L non-essential amino acids (NEAA), 0.1 mmol/L β-mercaptoethanol, 1 mmol/L pyruvate, 1 mmol/L
Alkaline phosphatase and immunofluorescence staining
Buffalo ES-like cells were subjected to alkaline phosphatase (AP) staining for characterization, for which the medium was removed from the cultures, and the cells were washed with phosphate-buffered saline (PBS) twice, then fixed in 4% paraformaldehyde for 20 minutes. After fixation, the cells were washed three times with PBS and incubated for 30 minutes at room temperature in BCIP/NBT (Amresco, Solon, OH). The stained ES-like cells were examined under the inverted microscope (Nikon, Tokyo, Japan). The expression of cell surface antigens, SSEA-1 and SSEA-4; pluripotency markers OCT4 and SOX2; and the β-Catenin were examined by immunofluorescence staining.
The putative buffalo ES-like cells were fixed with 4% paraformaldehyde in PBS for 30 minutes, washed three times in PBS, and then permeabilized in 1% Triton X-100 for 30 minutes. After thorough washing with PBS, ES-like cells were incubated with the blocking solution (1% bovine serum albumin in PBS) for 30 minutes at room temperature, and then incubated with the primary antibody at a dilution of 1:200 overnight at 4°C.
After washing three times with PBS, samples were incubated with the FITC-labeled appropriate secondary antibody at a dilution of 1:200 for 90 minutes at room temperature in the dark. Cells were washed twice with PBS, then costained with Propidium Iodide (PI; 3 μmol/L), and examined under a fluorescence microscope (Nikon). Fluorescence intensity was analyzed using Image Pro-plus 6.0 software to measure the relative expression of β-Catenin protein. The detailed procedure could refer to our previous report (Shen et al., 2017).
5-Bromo-2-deoxyuridine incorporation assay
5-Bromo-2-deoxyuridine (BrdU) incorporation assay was used to detect the proliferation of primary buffalo ES-like colonies. The day on which isolated ICM was used for seeding was considered as day 1. Primary buffalo ES-like colonies were treated with BrdU (0.3 μg/μL) for 24 hours, and subjected to BrdU immunostaining as follows.
After washing with PBS twice for 5 minutes, cells were fixed in 4% paraformaldehyde for 40 minutes at room temperature, and washed with PBS twice, permeabilized in 1% Triton X-100 for 40 minutes, washed with PBS twice, incubated in 4 M HCl for 30 minutes to break the DNA structure of the cells. Then, cells were washed with PBS twice, incubated with the blocking solution for 1 hour, incubated with the medium containing 2 μg/mL anti-BrdU overnight at 4°C. Cells were washed with PBS twice and incubated with the secondary antibody at a dilution of 1:200 for 90 minutes. After washing twice with PBS, the cells were costained with PI, visualized under a fluorescent microscope, the BrdU-positive cells were counted.
RT-PCR and qRT-PCR
RT-PCR and qRT-PCR were used to detect the pluripotent-related gene expression in passage 5 of buffalo ES-like colonies; the expression of PCNA gene (proliferating cell nuclear antigen) in buffalo ES-like colonies after treating with GSK3 inhibitors for 24 hours. RNA was extracted from buffalo ES-like colonies using the Cells-to-CT Kit. The first-strand cDNA was synthesized by the SuperScript II Reverse Transcriptase Kit (Thermo Fisher Scientific, Waltham, MA).
Briefly, the colonies were washed with ice-cold PBS and then heated in cell lysis buffer (Cells-to-cDNA™ II Kit; Ambion, Huntingdon, United Kingdom) at 75°C for 15 minutes. The cell lysate was treated with 1.3 μL 10 × RDD buffer, 1 μL DNase-I at 37°C for 30 minutes to digest genomic DNA; and treated with 1 μL EDTA, 1 μL dNTP, 2 μL random primer at 65°C for 10 minutes; then was treated with 2 μL DTT, 4 μL 5 × FS buffer, 0.25 μL reverse transcriptase, 0.5 μL RNase inhibitor RRI, 25°C for 5 minutes, 42°C for 60 minutes, 95°C for 10 minutes, and 4°C forever.
PCR was conducted with Taq DNA polymerase under the following conditions: 1 cycle with denaturing at 94°C for 5 minutes, subsequent 35 cycles of denaturation at 95°C for 30 seconds, at annealing temperature for 30 seconds, elongation at 72°C for 30 seconds, and final extension at 72°C for 10 minutes.
qRT-PCR was carried out by the ABI PRISM 7500 Real-Time System (Applied Bio-Systems, Foster City, CA). The qRT-PCR mixture (20 μL) contained 10 μL SYBR, 8 μL ddH2O, 1 μL primer, and 1 μL cDNA. Reaction conditions were as follows: 95°C for 10 minutes, followed by 40 cycles at 95°C for 30 seconds, 60°C for 30 seconds, and 72°C for 30 seconds. Each detection was repeated three times using different samples for each gene. The relative expression levels were normalized to β-actin gene. The relative expression level of target gene was determined by using the formula 2−▵▵CT. The details of primers, and reaction conditions for RT-PCR and qRT-PCR are listed in Supplementary Table S1.
Statistic analysis
The expression profiles of target genes were analyzed by chi-square analysis. The other data were using one-way ANOVA in a statistical analysis systems program (SPSS 17.0) to determine differences between the experimental groups. Differences were considered significant when p-values were <0.05.
Results
Characterization of buffalo ES-like cells
The primary colonies of buffalo ES-like cells were formed at 5–7 days after seeding of ICMs. The established colonies showed round and tightly packed morphology (Fig. 1A). Meanwhile, ES-like cells exhibited a high AP activity by AP staining and expressed OCT4, SOX2, SSEA-1, and SSEA-4 by immunofluorescence (Fig. 1B). RT-PCR also revealed that ES-like cells expressed pluripotent markers, including OCT4, SOX2, C-MYC, LIN28, E-Cadherin (CH1), and NANOG (Fig. 1C). These results suggest that buffalo ES-like have the characteristics of pluripotency.

Isolation, culture and pluripotency identification of buffalo ES-like cells.
The effect of GSK3 inhibitor on primary colony formation of buffalo ES-like cells
Effect of GSK3 inhibitor on primary colony formation of buffalo ES-like cells was investigated. The percentage of ICM attachment and primary colony formation were observed in Table 1. The results showed that there was no significant difference in the ICM attachment rate among the 0.5 μg/mL BIO group, 5 μmol/L CHIR99021 group, and the control group (91.18% and 92.98% vs. 94.59%, p > 0.05). However, the rate of primary colony formation in both CHIR99021 and BIO-treated groups was significantly improved compared with the control group (77.71% and 72.06% vs. 55.41%, p < 0.05). These results suggest that inhibition of GSK3 promotes the primary colony formation of buffalo ES-like cells.
Effect of Glycogen Synthase Kinase-3 Inhibitor on Formation of Buffalo Embryonic Stem Cell-Like Cell Primary Colonies
ES-like cells were isolated from ICMs of the blastocysts, the percentage of ICM attachment and primary colony formation were calculated.
Values are mean ± SD, the different superscripts in the same column indicate significant difference (p < 0.05).
BIO, 6-bromoindirubin-3′-oxime; ES-like cells, embryonic stem cell-like cells; ICMs, inner cell masses; SD, standard deviation.
The effect of GSK3 inhibitor on the proliferation of primary buffalo ES-like colonies
The effect of GSK3 inhibitor on the proliferation of primary buffalo ES-like colonies was studied. First, the day on which isolated ICM was used for seeding was considered as day 1, and the mitosis index of primary buffalo ES-like cells was detected by BrdU incorporation assay (Fig. 2A). As shown in Table 2, the proliferation rate of primary colonies in the group treated with CHIR99021 was significantly higher compared with the control group on day 1, day 3, day 4, and day 5 (p < 0.05); the group treated with BIO was remarkably higher compared with the control group on day 1 (p < 0.05), but there was no significant difference in other days (p > 0.05). Second, the average diameter of primary buffalo ES-like colonies was measured.
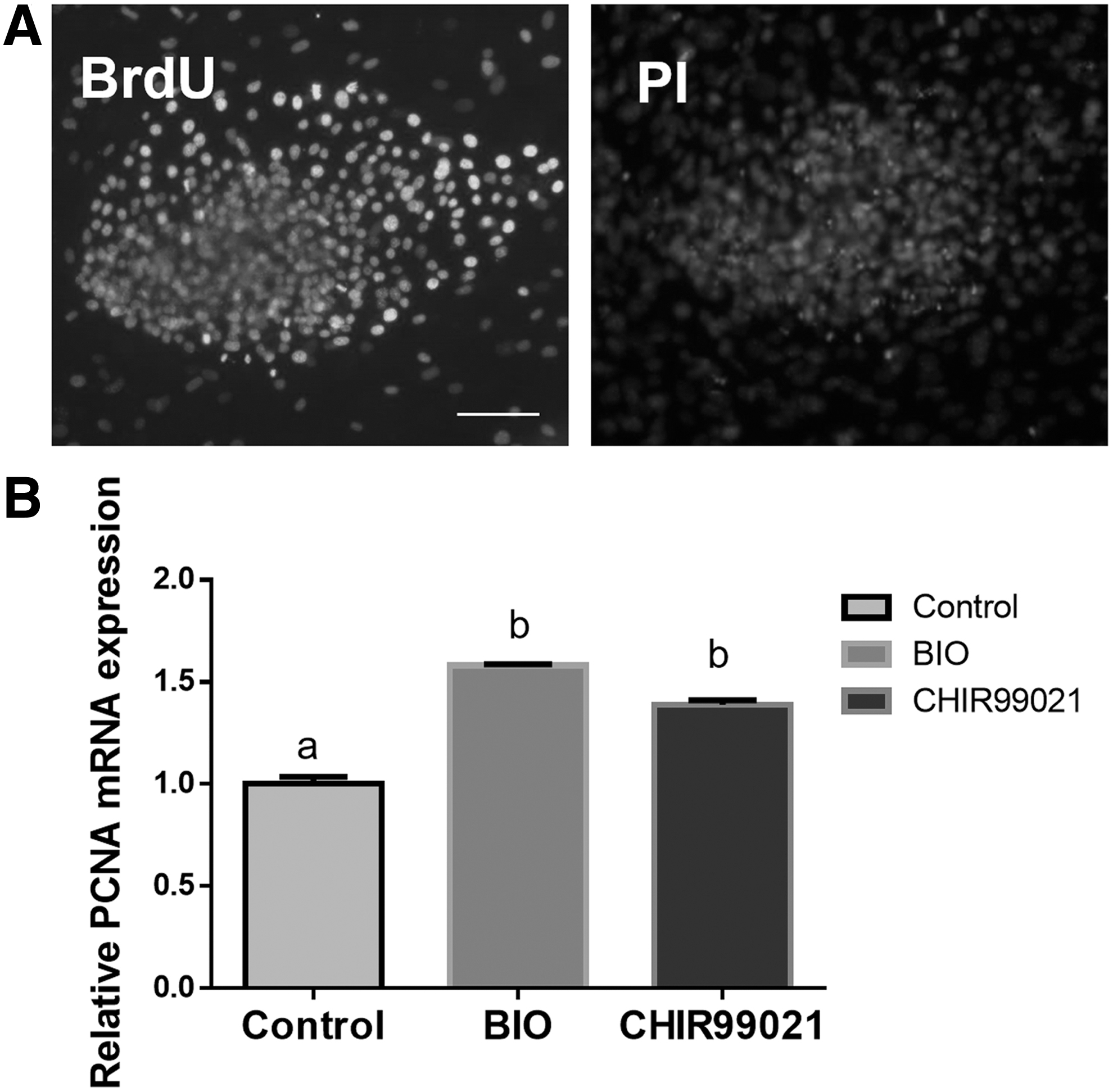
BrdU immunostaining analyzed the proliferation of buffalo ES-like cells
The Proliferation Rate of Buffalo Embryonic Stem Cell-Like Cells from Day 1 to 5
BrdU incorporation assay was used to detect the proliferation of buffalo ES-like cells' primary colonies in the culture from day 1 to 5.
Values are mean ± SD, data with different superscripts in the same row differ significantly (p < 0.05).
BrdU, 5-bromo-2-deoxyuridine.
As shown in Table 3, the average diameter of primary colonies in the group treated with CHIR99021 was evidently bigger compared with the control group (203.41 μm vs. 128.01 μm, p < 0.05), but there was no significant difference between the group treated with BIO and the control group (166.64 μm vs. 128.01 μm, p > 0.05). Furthermore, qRT-PCR result showed that the expression PCNA, a distinct marker for proliferative cells, was significantly upregulated in both CHIR99021 and BIO groups (p < 0.05, Fig. 2B). These results suggest that inhibition of GSK3 promotes proliferation of primary buffalo ES-like colonies.
Effect of Glycogen Synthase Kinase-3 Inhibitor on Average Diameter of Buffalo Embryonic Stem Cell-Like Cells’ Primary Colonies
After 5–6 days of culture, the ES-like cell morphology colony formed, diameter of primary colonies was measured by microscope.
Values are mean ± SD, the different superscripts in the same column indicate significant difference (p < 0.05).
The effect of GSK3 inhibitor on pluripotency maintenance of buffalo ES-like cells
The pluripotency maintenance is an important characteristic of stem cells. Buffalo ES-like cells could be subcultured for 15 passages in GSK3 inhibitor-treated medium, and continuously expressed pluripotency markers, OCT4, SOX2, SSEA-1, SSEA-4, LIN28, CH1, and NANOG (data not shown, the identification was the same as Fig. 1B,C). However, without GSK3 inhibitor, buffalo ES-like cells could only be subcultured for eight passages.
On the other hand, the effect of GSK3 inhibitor on relative expression level of pluripotency markers was detected by qRT-PCR. The results showed that the expression of OCT4, SOX2, and NANOG in buffalo ES-like cells treated with CHIR99021 or BIO was significantly upregulated compared with the control group (p < 0.05). Moreover, the expression levels of these genes in buffalo ES-like cells treated with CHIR99021 were significantly higher than those of the BIO-treatment group (p < 0.05, Fig. 3).

qRT-PCR analyzed the expression level of pluripotency marker genes in GSK3-inhibited buffalo ES-like cells. Bars labeled with different letters mean significantly different (p < 0.05). Control, ESCs culture medium-derived ES-like cells.
Meanwhile, the key downstream target genes of the Wnt pathway, β-Catenin, and C-MYC (also a pluripotency marker gene), were significantly upregulated with treating with CHIR99021 or BIO (p < 0.05, Figs. 3 and 4A,B). These results suggested that inhibition of GSK3 can prolong the in vitro culture time of buffalo ES-like cells and promote their pluripotency.

Immunofluorescence analyzed the expression
Discussion
As for the agricultural and biomedical purposes, the derivation of stable ESCs from domestic animals is important for reproduction, genetic breeding and improvement, and human disease model. Buffalo is one of the most important domestic ungulates that are usually used for food, labor, and bioreactors. However, it remains a challenge to establish stable buffalo ESC lines. Screening chemical supplements to optimize the culture condition will be an effective way to establish buffalo ES cell lines. Previous study reported that Wnt/β-Catenin plays a critical role in pluripotent stem cells. Wnt/β-Catenin pathway acted synergistically with LIF/JAK-STAT pathway to prevent differentiation of mouse ESCs (Hao et al., 2006).
Wnt/β-Catenin pathway promoted self-renewal and inhibits the primed state transition in both naive mouse and human ESCs (Xu et al., 2016). BIO and WNT3A combined to activate Wnt/β-Catenin signaling pathway could upregulate the expression of pluripotent-related genes in buffalo ES-like cells (Zandi et al., 2014, 2015). Our study further demonstrated that buffalo ES-like cells could maintain pluripotency and self-renewal in the stem cell culture medium with GSK3 inhibitor BIO or CHIR99021. Inhibition of GSK3 not only upregulated the pluripotency markers expression in buffalo ES-like cells, but also promoted the primary colony formation and proliferation.
Proliferation is a main characteristic of in vitro cultured pluripotent stem cells. Many reports demonstrated that regulating Wnt/β-Catenin pathway affected the proliferation of pluripotent stem cells, including the stem cells of domestic animals (Bogliotti et al., 2018; Fan et al., 2017; Li et al., 2020; Peng et al., 2014; Zhu et al., 2012). In this study, BrdU incorporation assay showed the proliferation rate of primary buffalo ES-like colonies in the CHIR99021- or BIO-treated group was significantly higher compared with the control group. Activation of Wnt/β-Catenin by BIO or CHIR99021 stimulated mitosis of buffalo ES-like cells. Correspondingly, the upregulated expression of proliferation marker genes, PCNA (Wrobel et al., 1996) and C-MYC (Denis et al., 1991) provided additional evidence that CHIR99021 and BIO could promote proliferation of buffalo ES-like cells.
How does β-Catenin affect stem cell proliferation is unclear. According to the previous studies about the role of Wnt/β-Catenin pathway, the main feature of this pathway is the regulation of cytoplasmic β-Catenin protein stability. In the absence of Wnt, β-Catenin is phosphorylated by a “destruction complex” consisting of adenomatous polyposis coli gene (APC), Axin, CK1, and GSK3β. Phosphorylated β-Catenin is degraded by the ubiquitin-proteasome system, thereby the level of cytoplasmic β-Catenin is low (Okita and Yamanaka, 2006).
In the presence of Wnt or treating with GSK3 inhibitor, Wnt binds to the Frizzled and LRP5/6 receptors to activate Dishevelled, to inhibit GSK3β from phosphorylating β-Catenin. As a result, β-Catenin accumulates in the cytoplasm and travels to the nucleus, where it associates with LEF (lymphoid enhancer factor)/Tcf (T cell factor) to activate transcription of Wnt target gene, including C-MYC, cyclin D1, and p53, which are associated with promoting proliferation (He et al., 1998; Peng et al., 2014; Tetsu and McCormick, 1999). These are the present data to explain why GSK3 inhibitors promote ESCs proliferation.
Some important pluripotency markers OCT4, SOX2, and NANOG were upregulated in buffalo ES-like cells when supplemented with GSK3 inhibitors. These results are in agreement with the previous report (Zhu et al., 2012). Tcf, a terminal component of the canonical Wnt pathway, which provides an important clue to explain the mechanism that Wnt pathway regulates stem cell state by the upregulated expression of OCT4, SOX2, and NANOG. TCF is a kind of transcription factor with two-way regulation function. It can inhibit gene transcription by combining with Groucho, and promote gene transcription by combining with β-Catenin.
Tcf3, a member of the Tcf/Lef family, occupies more than 1000 murine promoters (Cole et al., 2008), including those of the known Wnt targets, Axin2 and C-MYC, and important for maintaining pluripotency genes OCT4, SOX2, and NANOG (Pereira et al., 2006; Tam et al., 2008; Wray et al., 2011). Tcf3 promotes ESC differentiation by repressing the expression of OCT4, SOX2, and NAONG. Tcf3 knockdown results in the upregulation of OCT4, SOX2, and NAONG (Cole et al., 2008).
Therefore, depleting Tcf increases Wnt signaling, and GSK3 inhibitors alleviate Tcf3 repression of the pluripotency marker genes and increase ESC resistance to differentiation (Wray et al., 2011). GSK3 inhibitors that could reinforce pluripotency of ESCs may also be contributed by their role of stabilizing the protein level of β-Catenin. Stabilized β-Catenin can form a complex with CREB-binding proteins (CBP) and activate the expression of pluripotency marker genes, OCT4 and NANOG (Kelly et al., 2011; Takao et al., 2007; Takemaru and Moon, 2000). The regulation of β-Catenin/CBP complex plays a critical role in the maintenance of pluripotency in ESCs (Miki et al., 2011; Miyabayashi et al., 2007).
Especially, C-MYC is a key downstream target gene of Wnt/β-Catenin signaling pathway, and is also a marker gene of proliferation and pluripotency. C-MYC activation results in many biological effects, including driving cell proliferation, regulating cell population growth, apoptosis, differentiation, and self-renewal of stem cells. The upregulated expression of C-MYC implicated that CHIR99021 and BIO could inhibit GSK3β from phosphorylating β-Catenin, and activate Wnt pathway. GSK3β phosphorylation of C-MYC on T58 has been implicated in the LIF/STAT3-mediated regulation of mouse ESC pluripotency, since overexpression of a T58A mutant of C-MYC in mouse ESCs promotes their self-renewal and pluripotency in the absence of exogenous LIF (Zandi et al., 2015). Suppression of GSK3β activity, leads to elevated C-MYC levels, and hence enhances buffalo ES-like cells and stem cell derivation, stability, and self-renewal.
However, these Chinese swamp buffalo ES-like cells were subcultured 15 times in the ECM supplemented with GSK3 inhibitor, under our experimental conditions. We did not test for chimerism, which is considered the stringent test for pluripotency and developmental potency of pluripotent stem cells. So, our buffalo ESCs can only be called ES cell-like cells. On the basis that GSK3 inhibitor promotes the derivation of buffalo ES-like cells, further study will continue to optimize buffalo ESC culture conditions, and focus on the mechanism of Wnt/β-Catenin signaling pathway on self-renewal and pluripotency of buffalo ESCs.
Conclusion
In conclusion, GSK3 inhibitors BIO and CHIR99021 enhance the derivation of Chinese swamp buffalo ES-like cells, promote their self-renewal and pluripotency, and maintain the undifferentiated state in vitro. Those findings provide a guidance for screening more signaling pathways that have effective role in regulating the proliferation and pluripotency of buffalo ESCs.
Footnotes
Author Contributions
F.L., D.S., and Y.D. conceived and designed the study. Y.D., Y.L., and C.L. performed the experiments and drafted the article. Y.L., Q.R., and J.Z. collected and analyzed data. All authors read and approved the final article.
Author Disclosure Statement
The authors declare they have no competing financial interests.
Funding Information
This work was supported by the Natural Science Foundation of China (31560633, 31760666 and 31760334); Natural Science Foundation of Guangxi Province (2018JJA130074, 2018JJA130044); State Key Laboratory for Conservation and Utilization of Subtropical Agro-Bioresources (SKLCUSA-a201908), and Guangxi training project funded by the new century academic and technical leaders of special funds; Science and Technology Major Project of Guangxi (AA17204051), and Nanning Scientific Research and Technological Development Foundation (20192087).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.