
Abstract
Oxidative stress can impede maturation of the nucleus and cytoplasm of oocytes during in vitro maturation (IVM). Rhodiola sachalinensis, an herb commonly used in traditional Chinese medicine, conveys antioxidative effects to cryopreserved bovine sperm. Therefore, the aims of this study were to evaluate the effects of different concentrations of R. sachalinensis aqueous extract (RSAE) on IVM and subsequent in vitro embryonic development after parthenogenetic activation (PA), in vitro fertilization (IVF), and somatic cell nuclear transfer (SCNT). The results showed that RSAE supplementation (6 and 60 mg/L) significantly increased intracellular glutathione levels, but had no effect on maturation rates or reactive oxygen species. After in vitro culture, greater blastocyst formation was observed in PA embryos (6 mg/L RSAE), as well as in IVF and SCNT embryos (60 mg/L) matured in RSAE-supplemented IVM media. In conclusion, although there was no significant improvement in the maturation rate, RSAE supplementation conveyed an antioxidative effect during IVM, and improved subsequent embryonic development in vitro. Further studies are needed to explore gene expression pattern in oocytes and embryos treated with RSAE.
Introduction
Current methods for in vitro
Rhodiola sachalinensis is an herb commonly used in traditional Chinese medicine. Previous reports have found that R. sachalinensis extract increases RNA content in skeletal muscle, promotes the synthesis of adenosine triphosphate, and increases physical and mental energy in humans (Abidov et al., 2003; Zhang et al., 2007). R. sachalinensis has also been shown to promote oxidation resistance and prevent apoptosis (Yousef et al., 2006). Moreover, R. sachalinensis-derived saccharides are reported to increase antioxidant activities of frozen-thawed bovine semen (Cao et al., 2013). However, it remains unknown whether R. sachalinensis aqueous extract (RSAE) has a similar antioxidant effect on IVM of oocytes.
Therefore, the aims of the present study were (1) to investigate the effect of RSAE on the IVM and quality of porcine oocytes by detection of intracellular glutathione (GSH) and reactive oxygen species (ROS), and (2) to assess subsequent embryonic development after parthenogenetic activation (PA), in vitro fertilization (IVF), and somatic cell nuclear transfer (SCNT).
Materials and Methods
Chemicals
All chemicals were purchased from Sigma-Aldrich Corporation (St. Louis, MO), unless otherwise stated.
Preparation of RSAE
RSAE was a gift from Dr. Shushan Zhang (Shanghai Academy of Agriculture Sciences) and produced as described previously (Zhao et al., 2009). In brief, R. sachalinensis was dried and pulverized and soaked in distilled water at 80°C for 5 hours. After filtration, the filtrate was concentrated using a rotary evaporator and freeze-dried. RSAE was pulverized and maintained in dry storage at room temperature until used.
IVM of oocytes
Cumulus–oocyte complexes (COCs) with at least two to three layers of attached cumulus cells were selected before cultured at 38.5°C for 44 hours in maturation media (TCM-199; Gibco, Carlsbad, CA) supplemented with 10% (v/v) fetal bovine serum (FBS; Gibco), 10% (v/v) pig follicular fluid, 10 IU/mL of pregnant mare serum gonadotropin, 10 IU/mL of human chorionic gonadotropin, 10 ng/mL of epidermal growth factor, 0.1 mg/mL of glutamine, and 1% penicillin–streptomycin solution. Before treatment, RSAE was dissolved in IVM media to the final concentrations of 0.6, 6, and 60 mg/L. The maximum solubility of RSAE in water was 60 mg/L. The choice of RSAE concentrations was determined based on our preliminary results (unpublished data). Standard maturation media without RSAE was used as a control group. Oocytes with apparent first polar bodies were recorded as matured after IVM.
Evaluation of intracellular GSH levels, and ROS activities
Intracellular GSH levels were determined using GSH-GSSH assay kit (Beyotime Institute of Biotechnology, Shanghai, China) in accordance with the manufacturer's instructions. In brief, a standard curve of GSH was plotted of dilutions of 10 mM GSH stock solution at the concentrations of 0.5, 1, 2, 10, and 15 μM (Fig. 1). Then 10 μL of protein removal reagent M was added to each sample containing 30 denuded matured oocytes from each group, and all the samples were snap frozen in liquid nitrogen and thawed in a water bath at 37°C (repeated 4 times). The optical density of each sample at a wavelength of 405 nm was determined using an ELx808 Absorbance Microplate Reader (BioTek Instruments, Winooski, VT) to determine the concentration of GSH against the standard curve. Four replicates of each group were assayed.
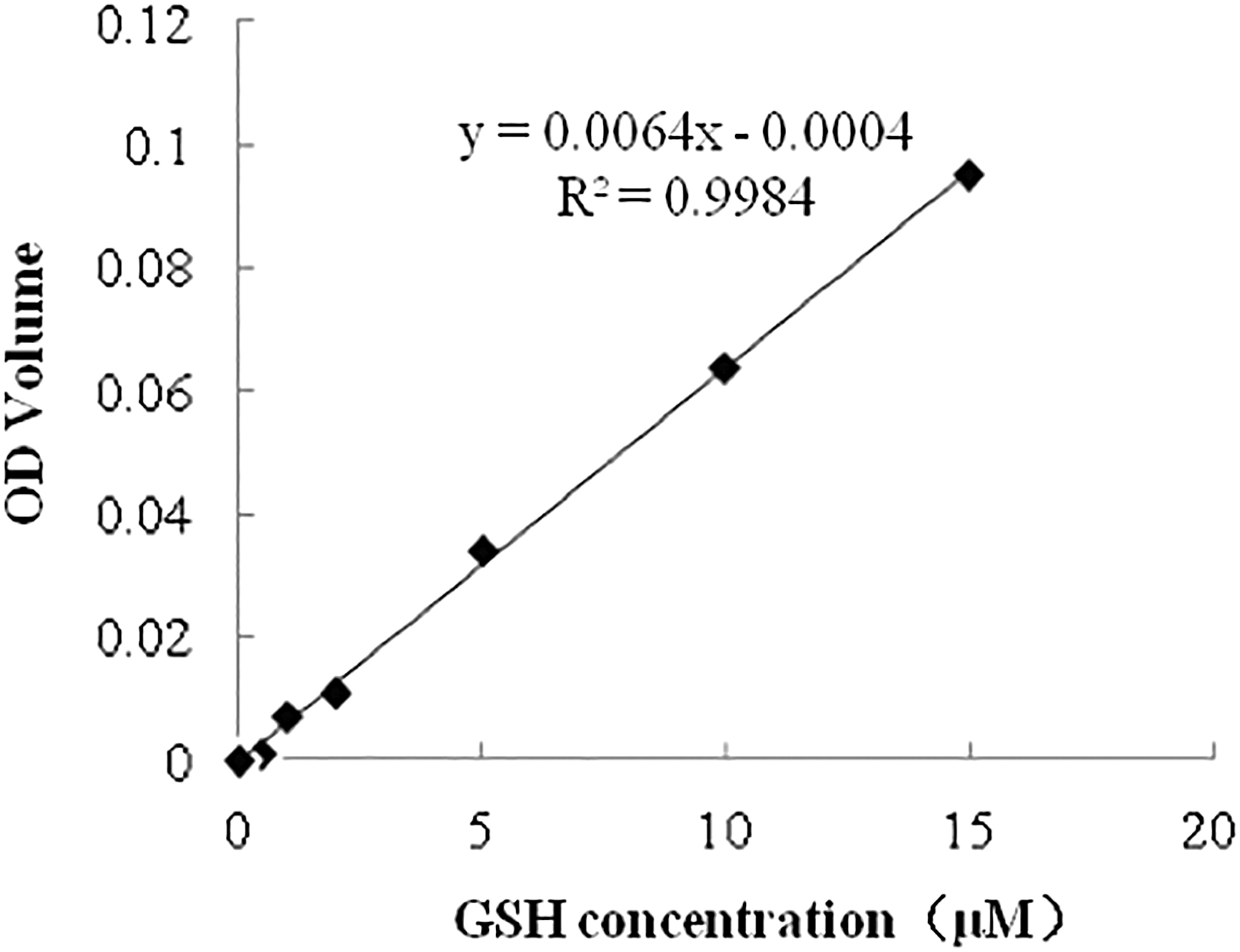
Standard curve to determine the concentrations of GSH in the oocytes from different groups. GSH concentrations (μM) are shown on x-axis and OD values on the y-axis. GSH, glutathione; OD, optical density.
Intracellular ROS activity of the oocytes was measured using the 2′,7′-dichlorofluorescein (DCF) fluorescence assay as previously described (Gupta et al., 2010). In brief, denuded matured oocytes were incubated with 10 mM DCF diacetate for 20 minutes at 38.5°C. After washing three times with phosphate-buffered saline (PBS), intracellular fluorescence of DCF was observed and images were captured under a fluorescent microscope (Nikon Corporation, Tokyo, Japan) at a wavelength of 450–490 nm. Fluorescence intensity was analyzed using Image-Pro Plus 6.0 Image Analysis Software (Media Cybernetics, Bethesda, MA). Representative images of ROS staining of the different groups are shown in Figure 2. This experiment was repeated 4 times with 30 oocytes per group in each replicate.
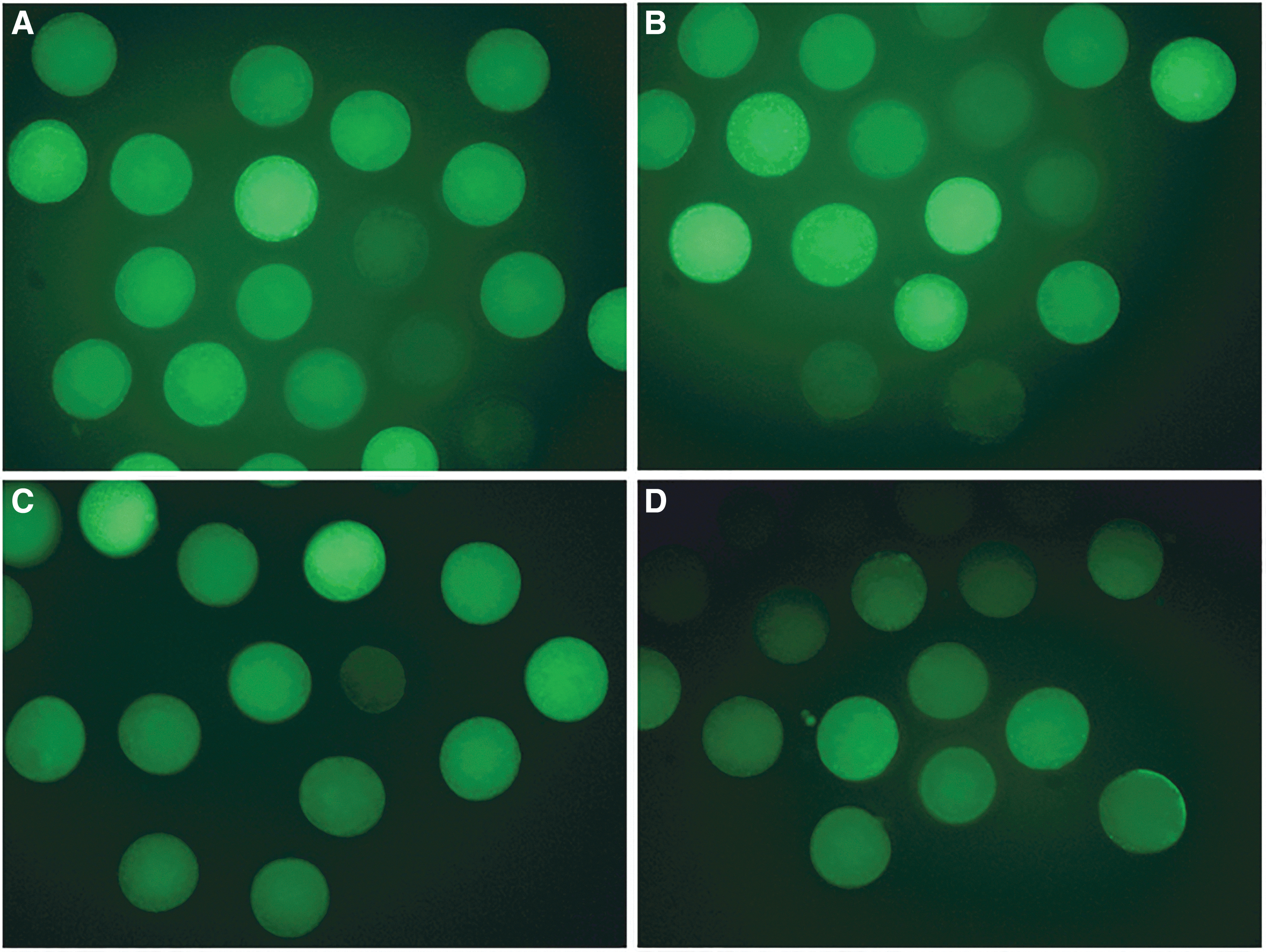
Representative images of fluorescence staining for ROS activity in matured porcine oocytes.
Parthenogenetic activation
After IVM, denuded oocytes were activated in fusion/activation medium (0.25 mM mannitol, 0.1 mM CaCl2, 0.1 mM MgCl2, 0.5 mM Hepes, 0.01% polyvinylpyrrolidone [w/v]) with a single direct current pulse of 1.2 kV/cm for 30 μs (BTX Instruments, Inc., San Diego, CA).
In vitro fertilization
The IVF protocol is described in our previous report (Wu et al., 2019). In brief, fresh semen was washed twice in PBS supplemented with 0.1% bovine serum albumin (BSA) at 700 g for 4 minutes, and once in IVF medium (0.11 M NaCl, 3 mM KCl, 7.5 mM CaCl2·2H2O, 0.11 M glucose, 5 mM sodium pyruvate, 2 mM caffeine, 1 g/L BSA), and then diluted to 1 × 107 sperm per milliliter. The sperm were resuspended with IVF medium and maintained in a 5% CO2 incubator at 38.5°C for 90 minutes. The COCs were washed and maintained in IVF drops for 45 minutes before adding sperm. The final concentration of sperm was 5 × 105 sperm per milliliter. After incubation for 6 hours, presumptive zygotes were pipetted to remove the cumulus cells before in vitro culture.
Somatic cell nuclear transfer
Before SCNT, porcine fetal fibroblasts reached at 80% confluence were starved by culturing in Dulbecco's modified Eagle's medium supplemented with 0.5% FBS for 1–2 days. SCNT was performed as described in our previous report (Wu et al., 2019). In brief, the nucleus and the first polar body of each oocyte were removed with a pipette under a polarized light microscope (PerkinElmer, Inc., Waltham, MA), and then the donor cell was injected into the perivitelline space. Reconstructed embryos were then fused in fusion/activation medium with a single direct current pulse of 1.2 kV/cm for 30 μs (BTX Instruments, Inc.).
Embryo culture, evaluation, and total cell number per blastocyst
The day on which PA, IVF, and SCNT were performed was considered to be day 0. The embryos were then cultured in PZM-3 (Yoshioka et al., 2002) for 7 days at 38.5°C with 5% CO2 in air with maximum humidity. The cleavage and blastocyst rates were evaluated at day 2 and 7. For the assessment of total cell number, the blastocysts (PA, SCNT, or IVF) at day 7 were stained with 10 μg/mL of Hoechst 33342 for 20 minutes and the number of cells was counted under the fluorescence microscope (Nikon).
Statistical analysis
One-way analysis of variance was used for data analysis. All statistical analyses were performed using PASW Statistics for Windows, version 18.0 (SPSS, Inc., Chicago, IL). A probability (p) value of <0.05 was considered statistically significant.
Ethics
No live animals were involved in this study. The pig fetus was obtained from a sow at slaughter. Ovaries were obtained from a local slaughterhouse and semen was purchased from a commercial company.
Results
Effect of RSAE on maturation, GSH levels, and ROS activities of porcine oocytes
As shown in Table 1, there were no statistically significant differences in the maturation rate or ROS activity among the three tested RSAE concentrations and control groups, although ROS activity tended to decrease with increased concentrations of RSAE in IVM media. GSH concentrations were significantly higher at RSAE concentrations of 6 and 60 mg/L compared with the control group (3.81 and 3.64 vs. 2.39, respectively, p < 0.05).
Effects of Rhodiola sachalinensis Aqueous Extract-Supplemented In Vitro Maturation Media on Maturation Rates, Intracellular Glutathione Concentrations, and Reactive Oxygen Species Activities of Porcine Oocytes
Different superscript letters in same column indicate significant differences (p < 0.05). Four replications were performed.
GSH, glutathione; ROS, reactive oxygen species; RSAE, Rhodiola sachalinensis aqueous extract; SEM, standard error of the mean.
Effect of RSAE on embryonic development after PA, IVF, and SCNT
With regard to the developmental competence of embryos, embryonic development after PA, IVF, and SCNT differed among the oocytes treated with different RSAE concentrations (Tables 2–4). For PA embryos, a higher cleavage rate was observed with 60 mg/L of RSAE, compared with the control group (p < 0.05), but the highest blastocyst rate was obtained with 6 mg/L of RSAE (p < 0.05).
Effect of Rhodiola sachalinensis Aqueous Extract-Supplemented In Vitro Maturation Media on Embryonic Development After Parthenogenetic Activation
Cleavage and blastocyst rates were calculated from the number of embryos in culture (four replicates).
Different superscript letters in same column indicate significant differences (p < 0.05).
Effect of Rhodiola sachalinensis Aqueous Extract-Supplemented In Vitro Maturation Media on Embryonic Development After In Vitro Fertilization
Cleavage and blastocyst rates were calculated from the number of embryos in culture (four replicates).
Different superscript letters in same column indicate significant differences (p < 0.05).
Effect of Rhodiola sachalinensis Aqueous Extract-Supplemented In Vitro Maturation Media on Embryonic Development After Somatic Cell Nuclear Transfer
Cleavage and blastocyst rates were calculated from the number of reconstructed embryos (three replicates).
Different superscript letters in same column indicate significant differences (p < 0.05).
For IVF embryos, the cleavage rates were increased at all three RSAE concentrations compared with the control group. However, only 60 mg/L of RSAE resulted in significantly greater blastocyst formation compared with the control group (p < 0.05), while there was no significant differences in blastocyst formation among the different RSAE concentrations. Enhanced cleavage and blastocyst rates were only observed in SCNT embryos treated with 60 mg/L of RSAE group (p < 0.05).
Moreover, there were no significant differences in cell number per blastocyst in the PA, IVF, and SCNT embryos among the different RSAE concentrations versus the control groups, suggesting that the quality of blastocysts was not affected by RSAE treatment.
Discussion
To the best of our knowledge, this study is the first to explore the antioxidative effect of RSAE on IVM of porcine oocytes and subsequent embryonic development.
Intracellular GSH is an established molecular marker to predict the maturation of cytoplasm (Wang et al., 1997). Increased GSH storage during oocyte maturation by antioxidant supplementation of IVM media may have further protected the embryo against oxidative aggressions during the early developmental stages (Khazaei and Aghaz, 2017). In the present study, GSH concentrations were increased both in oocytes matured in 6 and 60 mg/L of RSAE supplemented IVM media. Moreover, greater blastocyst formation was observed with two higher concentrations of RSAE groups after PA or IVF and SCNT, indicating that GSH concentrations in the oocytes might be positively correlated to embryonic development.
However, there were no significant changes in ROS activities in the oocytes among all the RSAE-supplemented groups, suggesting limited ability of tested RSAE concentrations to reducing oxidative stress.
Parthenogenetically activated embryos normally have been conventionally used as a model to determine optimal antioxidant conditions before SCNT (Jin et al., 2016; Qasim et al., 2019). Interestingly, in the present study, the highest blastocyst formation of PA embryos were obtained with 6 mg/L of RSAE, whereas the blastocyst rates of IVF and SCNT embryos were increased for oocytes treated with 60 mg/L of RSAE, compared with the control and the other two RSAE concentrations. The mechanisms of PA embryonic development and epigenetic status differ from IVF and SCNT embryos (Deshmukh et al., 2011; Paffoni et al., 2008). Therefore, the requirement of oocyte ability to support IVF and SCNT embryos may differ from that of PA. These results suggest that the optimal condition of antioxidants for PA embryos may not be the optimal condition for SCNT.
Since maximum solubility of RSAE in water is 60 mg/L, it was not possible to test whether higher RSAE concentrations would improve oocyte quality (e.g., reduce ROS levels) and subsequent SCNT embryonic development in the current study. Hence, further studies are needed to identify the active ingredients in RSAE that could protect porcine oocytes against ROS and further improve embryonic development after SCNT.
In conclusion, although the maturation rates of oocytes cultured in RSAE-supplemented IVM media did not significantly improve, RSAE conveyed antioxidative effects by increasing GSH concentrations in the oocytes during IVM. RSAE at higher concentrations was also beneficial for embryonic development (6 mg/L for PA and 60 mg/L for IVF and SCNT). Further studies are needed to explore gene expression pattern related to apoptotic and ROS in oocytes and embryos treated with RSAE.
Footnotes
Author Disclosure Statement
The authors declare they have no competing financial interests.
Funding Information
This study was supported by grants from Shanghai Committee of Science and Technology (Grant No. 18391903700) and Ningbo Science and Technology Innovation 2025 Major Project (Grant No. 2019B10023).