
Abstract
The establishment of an in vitro culture system for complete oocyte maturation from the early stages of ovarian follicles is still a challenge. The aim of the present study was to assess the effect of different matrix with different culture media on the developmental growth of ovarian follicles in vitro. An ovarian histoarchitectural study was carried out to identify the primordial (0.027–0.039 mm), primary (0.041–0.079 mm), small preantral (0.085–0.131 mm), large preantral (0.132–0.294 mm), small antral (0.387–0.589 mm), and large antral (1.188–1.366 mm) follicles. Thus, large preantral follicles (0.2–0.3 mm) were mechanically isolated and cultured subsequently in different microconditions such as Dulbecco's modified Eagle's medium, Tissue Culture Medium-199 (TCM-199) and Opti-minimum essential medium, with same supplements where control (without matrix) was compared with matrix (coculture and encapsulation), which includes (1) buffalo fetal fibroblast cells, (2) cumulus cells, (3) ovarian mesenchymal cells, (4) collagen, (5) gelatin, and (6) Matrigel, cultured for 7 days in CO2 incubator at 38.5°C (5% CO2 in air). Cultured follicles were evaluated for growth rate (107.88% ± 10.24%), maturation rate (51.06% ± 6.53%), survivability rate (56.52% ± 3.42%), and antioxidant (catalase; CAT [1.58 ± 0.04 U/mg], superoxide dismutase; SOD [4.63 ± 0.05 U/mg], lactate dehydrogenase; LDH [1.48 ± 0.01 U/mg]) enzymatic activities, which showed significantly (p < 0.05) positive results in growth model with media TCM-199 than other studied groups. Furthermore, the development of large preantral follicles augmented significantly (p < 0.05) for growth rate (248.54% ± 9.51%), maturation rate (75.81% ± 7.07%), survivability rate (81.82% ± 3.02%), antioxidant (CAT [2.05 ± 0.03 U/mg], SOD [3.13 ± 0.12 U/mg], LDH [2.55 ± 0.51 U/mg]), and estradiol (175.83 ± 5.92 pg/mL) activities when they were encapsulated in Matrigel with nutritional requirements fulfilled by media TCM-199. These results provide better insight for the optimization of culture conditions for in vitro follicular development in the water buffalo, which will eventually assist in resolving the limitation of obtaining fewer competent oocytes for the embryo production in the species.
Introduction
Various advanced assisted reproductive techniques have been adapted to produce healthy offspring, but with low reproducibility rate. The main cause of this fact is inability to produce a large number of usable oocytes. Hence, in vitro follicular growth is serving as a hope to improve the possibility to retrieve the sufficient number of oocytes from a single ovary.
Historically, it has been stated that the capacity of reproduction in buffalo species is poor in comparison to other livestock animals. The reason being, buffaloes have a less number of primordial follicles and higher atresia rate (Bharadwaz and Roy, 1999), which successively reduces the availability of preantral follicles and subsequently oocytes. Various other factors influencing the growth of buffalo preantral follicles include the presence of corpus luteum, seasonal recovery, and presence of dominant follicles, at the time of recovery (Nandi et al., 2009).
One of the major difficulty level to mimic the in vivo environment accurately, originates from its unique architecture, where an oocyte is surrounded by follicular cells, consists of a vascular system, and creates a three-dimensional (3D) structure. As oocyte growth critically depends on the bidirectional autocrine and paracrine signaling between the theca cells, granulosa cells, and other constituents, due to this mutualism, follicular architecture should be maintained during its growth process for the correct acquisition of competent oocytes. Achievements in the preantral follicle culture system with live birth have been reported in mice (Eppig and Schroeder, 1989), but to replicate the same in large mammals is still a challenge. This may be due to the technical difficulty of recovering a large number of follicles from the dense and fibrous cortex of ovaries.
Another reason, perhaps is the two-dimensional (2D) follicular culture system, which is not sufficient to maintain the normal architecture of in vitro follicles, and new technological advancements have emerged in the form of 3D follicle culture system, which is able to retain the structure of normal follicles. To acquire the 3D integrity of in vitro follicle, three different ways have been known: (1) in situ/organ culture; (2) isolated follicles cultured on 3D system, which includes different extracellular matrix; and (3) 3D printing microporous scaffolds (Rossi et al., 2019).
As the microenvironment of in vitro growth of follicles is still unclear, to provide a better understanding and simplifying this procedure, a comparison between various coculture and encapsulation matrix, along with different commercially available basal media, which supports differently to different cell types. Therefore, the present work is designed to standardize the culture conditions for in vitro growth of preantral buffalo follicles, to obtain oocytes that can be considered developmentally competent for embryo production in the future. The frequency of follicular growth is also estimated by biochemical assays that include catalase (CAT), superoxide dismutase (SOD), lactate dehydrogenase (LDH), reactive oxygen species (ROS), and estradiol variation during the growth model.
Materials and Methods
All the culture media and chemicals were supplied from Sigma Chemicals (St. Louis, USA), if not otherwise stated. Plastic wares used were acquired from Nunc (Roskilde, Denmark). Fetal bovine serum was obtained from Gibco Life Technologies (Massachusetts, USA). Buffalo ovaries were collected from Delhi abattoir (in accordance with institutional ethics guidelines).
Histological characterization of ovarian follicles
Follicles from 10 buffalo ovaries were characterized by histomorphology using standard protocol (Bhardwaj and Sharma, 2017). Ovarian tissues were fixed, dehydrated, embedded, rehydrated, and finally stained with Hematoxylin and Eosin. Digital images of histological structures were obtained using an inverted microscope (Nikon Eclipse-Ti, Japan) at 40 × . Follicles were characterized on the basis of their sizes, as earlier mentioned by Palma et al. (2012).
Isolation of preantral follicles
Cortical pieces of 1 mm thick, from 434 buffalo ovaries, were sliced and subjected to mechanical dissection of large preantral follicles (0.2–0.3 mm). Follicles with circular outline, uniformly distributed granulose cells, and prominent oocyte were isolated in isolation medium (minimum essential medium [MEM] supplemented with 0.3% bovine serum albumin, 2 mM
Culture of preantral follicles
Isolated preantral follicles were washed and incubated in neutral red (15 μg/mL) for 30 minutes. Preantral follicles, which have taken red color, were considered as viable and further cultured in Dulbecco's modified Eagle's medium (DMEM), Tissue Culture Medium-199 (TCM-199), Opti-MEM (reduced-serum medium) supplemented with 5% follicular fluid (FF), 10% fetal bovine serum, 2 mM
The day of culture was considered as day 0 and medium was replenished at day 3 and 5. Growth of follicles was assessed by measuring the distance between the two ends, horizontally and vertically, by NIS-element BR software on day 0, 3, 5, and 7. At day 7, cultured follicles were again stained by neutral red to check the viability of growing follicles. The growth and survival rate was measured by parameters reported by Nandi et al. (2017).
Conditions for in vitro culture
Somatic cell coculture system
Primary cultures for buffalo fetal fibroblast cells (BFFC), cumulus cells (CC), and ovarian mesenchymal cells (OMC) were established and cryopreserved, as described earlier (Mehta et al., 2018).
After isolation, these cells were subsequently treated with mitomycin-C to form a uniform and nongrowing feeder layer, which provides a matrix for the coculture of preantral follicles.
Encapsulation system
Collagen type I (COL), Gelatin (2%) (GEL), and Matrigel (1:1) (MAT) (Corning, Life Science, USA) matrix was used for encapsulation.
Oocyte recovery and in vitro maturation
Oocytes were mechanically harvested after the culture of day 7, under a zoom stereo microscope (Nikon SMZ745T, USA). Oocytes with a compact, homogeneous granular ooplasm, and unexpanded cumulus mass having >2 layers of CCs, were selected for in vitro maturation (Fig. 3A). Isolated oocytes were matured for 24 h at 38.5°C, as mentioned by Dua et al. (2019). An oocyte with expanded cumulus layers and extruded polar body, stained with Hoechst 33342 nuclear stain (10 μg/mL), was considered as matured (Fig. 3B, C).
Chromosomal analysis
Matured oocytes obtained were thought to be arrested at the metaphase stage, which was directly subjected to karyotyping, mentioned by Yadav et al. (1997). The chromosome spreads obtained, were observed under the compound microscope (Leica Mikroskopie & Systeme GmbH, Germany) at 1000 × .
Biochemical assays
Homogenate of cultured preantral follicles for 7 days from different treatments in vitro was used separately as a test sample. CAT assay and SOD assay were performed according to the methods mentioned by Aebi (1984) and Marklund and Marklund (1974), respectively. Enzymatic extract of follicles from different culture conditions was collected separately at day 0, 3, 5, and 7, which was used to measure LDH cytotoxicity and levels of ROS by Watts (1979) and spectrofluorimetric method (Nandi et al., 2017), respectively. Estradiol estimation was determined by the ELISA Kit (Cayman Chemicals, MI, USA) and performed as per the manufacturer's instructions.
Experimental design
To investigate the effect of different matrix on the growth of follicles in vitro, six different matrices (BFFC [experiment 1], CC [experiment 2], OMC [experiment 3], COL [experiment 4], GEL [experiment 5], and MAT [experiment 6]) were compared with control (without matrix), which was subsequently combined with three different culture media (DMEM, TCM-199, and Opti-MEM). The above experiments were performed in triplicates, containing 20–25 follicles per trial per culture media.
Statistical analysis
A randomized study was conducted to evaluate the effect of each combination on follicular growth and survival rate of follicles; maturation rate of oocytes; biochemical assays (CAT, SOD, LDH); ROS, and estradiol production. Analysis was performed using GraphPad Prism software (San Diego, USA) by ANOVA followed by Tukey's test to compare the mean of different groups between the viability rate, biochemical activities, ROS, and estradiol production rate. Whereas, the significant differences in growth rate was tested by t-test to compare the mean diameter on day 0 and 7. The values obtained from each experiment were normalized, wherever required. Significant values were considered at p < 0.05. Data are presented in the form of mean ± SEM.
Results
Histological assessment
Ovarian follicles were characterized on the basis of their size, observed in histological sections. Follicles, ranging from 0.027 to 0.039 mm (0.033 ± 0.001), were classified as primordial follicles; 0.041–0.079 mm (0.051 ± 0.003) as primary follicles; 0.085–0.131 mm (0.107 ± 0.004) as small preantral follicles; 0.132–0.294 mm (0.216 ± 0.009) as large preantral follicles; 0.387–0.589 mm (0.482 ± 0.038) as small antral follicles, and 1.188–1.366 mm (1.277 ± 0.089) as large antral follicles (Fig. 1).
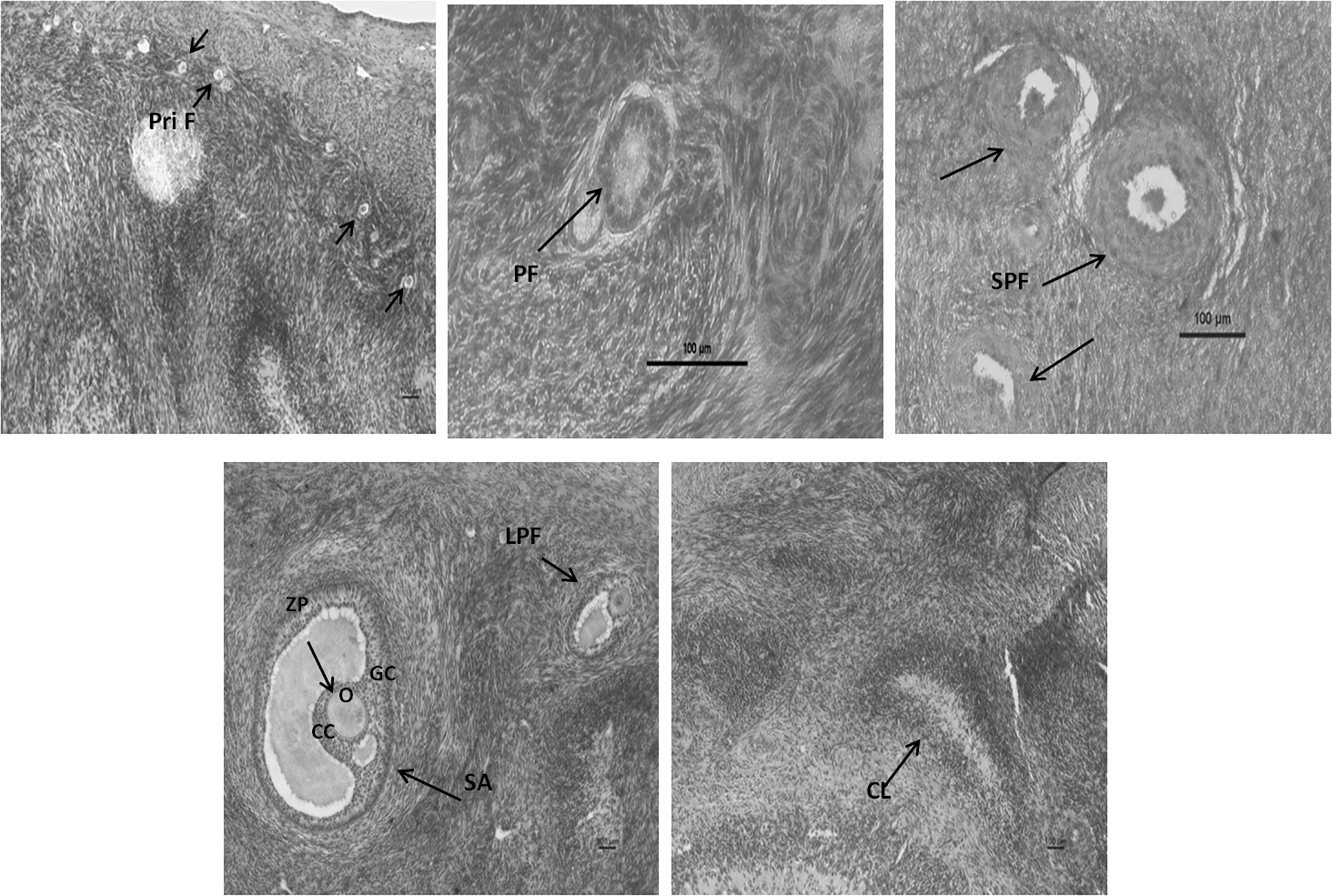
Histoarchitecture of buffalo ovary sections stained with Hematoxylin and Eosin showing: primordial follicle (Pri F); primary follicle (PF); small preantral follicle (SPF); large preantral follicle (LPF); small antral (SA); corpus luteum (CL); oocyte (O); cumulous cells (CC); granulosa cells (GC); zona pellucida (ZP). Scale bars represent 100 μm.
In vitro growth rate of large preantral follicles in different media and matrix
Large preantral follicles, ranging from 0.2 to 0.3 mm, were mechanically isolated from ovarian cortical tissue (Fig. 2A, B). Isolated follicles were stained with neutral red for viable assessment (Fig. 2C) and cultured for 7 days (Fig. 2D–G). When the follicles were grown in different media, without any matrix, the percentage change in follicular growth of preantral follicles (n = 192) after day 7, was significantly (p < 0.05) higher in TCM-199 (248.54% ± 9.51%) followed by DMEM (195.87% ± 9.02%) and Opti-MEM (127.00% ± 11.28%). Furthermore, follicles (n = 1331) were cultured to evaluate the growth rate in different basal media, in combination with different somatic cell coculture system and encapsulating agents. Follicles developed in TCM-199, with MAT matrix, showed a significantly (p < 0.05) double growth rate (Table 1).
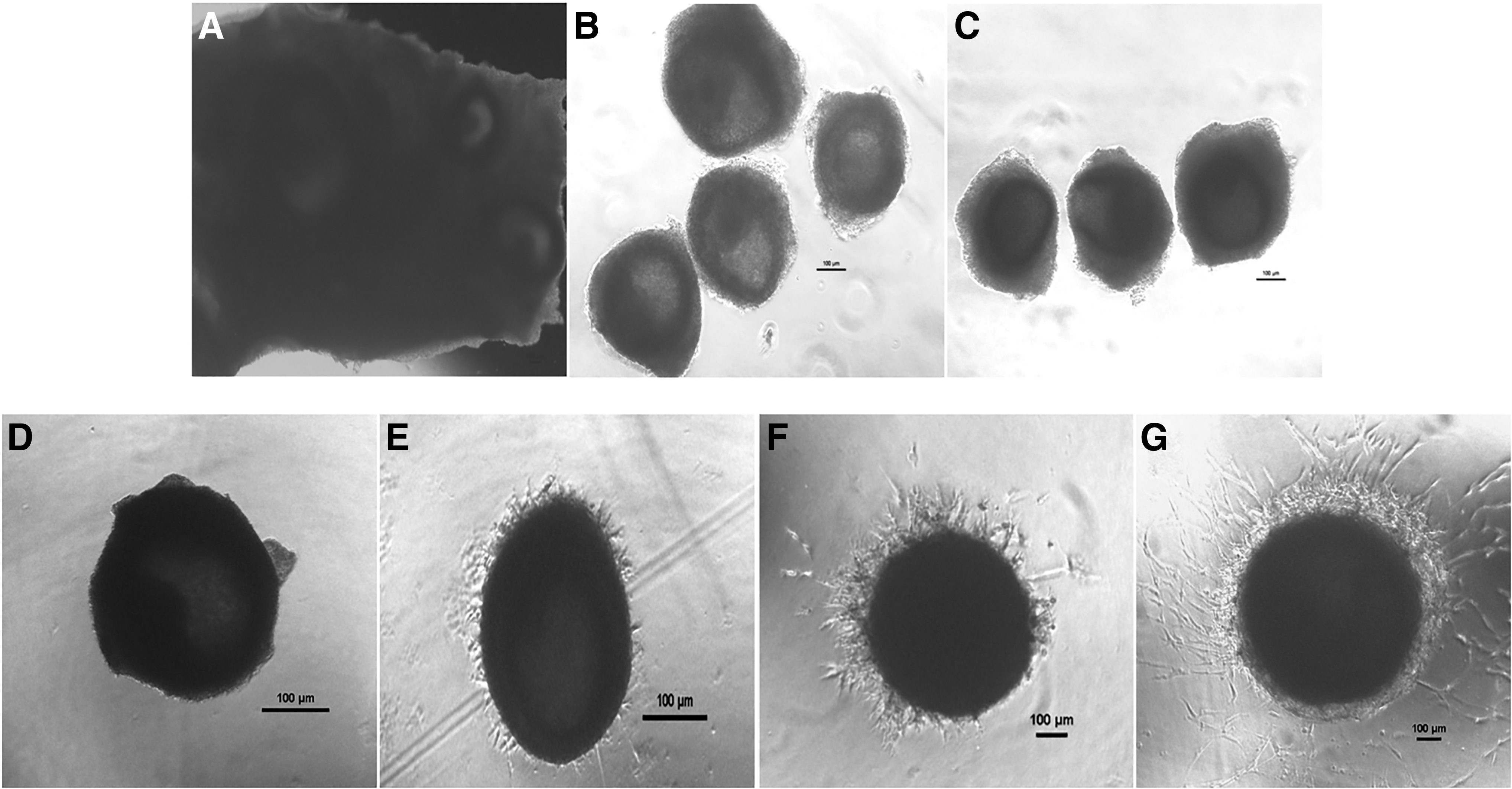
Isolation and culture of buffalo ovarian follicles in TCM-199 media encapsulated in Matrigel: Ovarian buffalo cortical tissue
Effect of Basal Medium with Different Matrix on Follicular Growth of Preantral Follicles on Day 7 and Maturation Rate of Oocytes Obtained from In Vitro Cultured Follicles
Values are mean ± SEM. Values with different superscripts differ significantly (p < 0.05).
Single media with single matrix cultured on different days (rows).
Single media with different matrix (columns).
Single matrix with different media (columns).
CC, cumulus cell; COL, collagen type I; DMEM, Dulbecco's modified Eagle's medium; GEL, gelatin; MAT, Matrigel; MEM, minimum essential medium; OMC, ovarian mesenchymal cell; TCM-199, Tissue Culture Medium-199; SEM, standard error of the mean.
Effect of different culture system on the viability rate of preantral follicles
The percentage viability of preantral follicles cultured in vitro for 7 days in TCM-199 showed significantly (p < 0.05) increased survivability rate than follicles cultured in DMEM and Opti-MEM basal media (Table 2). When compared within basal media, nearly a similar pattern was observed that affected the viability rate in each matrix, that is, MAT, COL, CC, OMC, BFFC, GEL, and control (Table 2).
Viability (%) of Preantral Follicles Cultured for 7 Days in Different Culture Conditions
Values with different superscripts differ significantly (p < 0.05).
Single matrix with different media (rows).
Single media with different matrix (columns).
Oocytes recovered and chromosomal analysis
A total of 1073 oocytes were recovered, after the culture of preantral follicles for 7 days, out of which 630 were matured (Table 1; Fig. 3A, B), stained (Fig. 3C), and subjected to chromosomal analysis that showed normal haploid chromosome (n = 25) complementation (Fig. 3D).
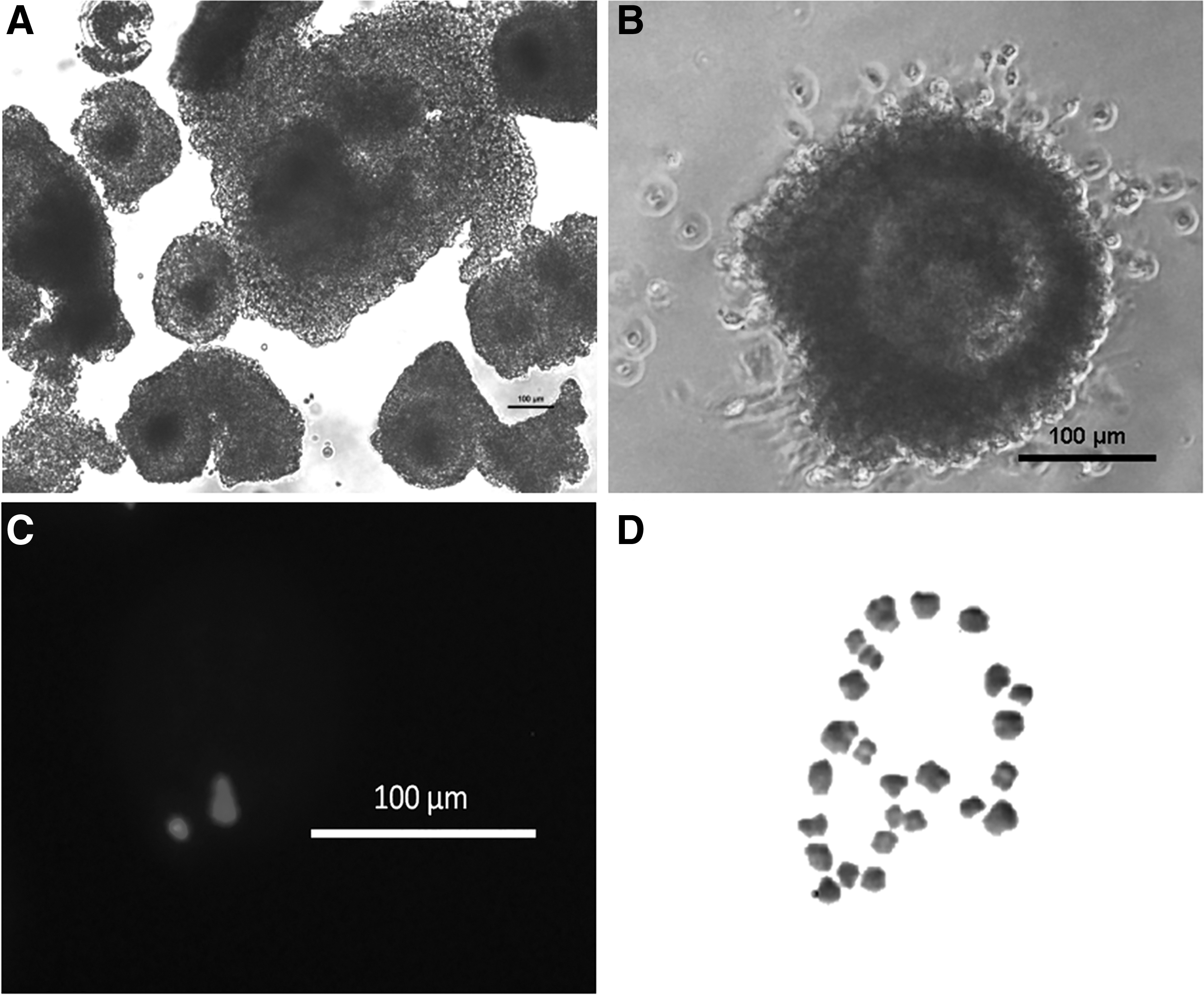
Immatured oocytes isolated after 7 days culture
Effect of different culture systems on CAT, SOD, LDH activity, ROS, and estradiol production
The CAT activity increased significantly (p < 0.05), whereas SOD activity decreased in preantral follicles cultured for 7 days when compared with follicles on day 0, which were proportional to a subsequent change in the follicular size. LDH activity showed significantly (p < 0.05) increased level in TCM-199 with MAT matrix; whereas other combinations did not show a significant difference (Table 3).
Measurement of Catalase, Superoxide Dismutase, and Lactate Dehydrogenase Activity of Preantral Follicles Cultured for 7 Days in Different Culture Conditions
Values are mean ± SEM. Values with different superscripts differ significantly (p < 0.05).
Day 0 compared with day 7 culture conditions (columns).
Single media with different matrix (columns).
ROS production was decreased significantly (p < 0.05) approximately threefold in TCM-199 with MAT and COL matrices, whereas ROS production remained constant in cell coculture system (Table 4). Estradiol production was 175.83 ± 5.92 pg/mL, when cultured in TCM-199 with MAT at day 7, which showed a significant (p < 0.05) increase as compared with other groups (Table 5).
Measurement of Reactive Oxygen Species Production (Fluorescent Unit) of Preantral Follicles Cultured for 7 Days in Different Culture Conditions
Values are mean ± SEM. Values with different superscripts differ significantly (p < 0.05).
Single media with single matrix cultured on different days (rows).
ROS, reactive oxygen species.
Determination of Estradiol Production on Day 7 for Preantral Follicles Developed in Different Culture Conditions
Values are mean ± SEM. Values with different superscripts differ significantly (p < 0.05).
Single matrix with different media (rows).
Single media with different matrix (columns).
Discussion
An efficient procedure for in vitro culture of buffalo preantral follicles was optimized in the present study. The study included the comparison of various matrices along with the affect of basal media. Initially, large preantral follicles were cultured in basal media alone, which was considered as control (without matrix), to observe the sole affect of media on growth pattern and further it was considered as standard to compare the affect of different matrices used in the study.
As a result, TCM-199 media have shown approximately fourfold better results in comparison to DMEM and Opti-MEM media. It could be primarily because of the variation in glucose content, that is, DMEM/F-12 (17.5 mM), TCM-199 (5.5 mM), and Opti-MEM (22.6 mM), which explains the diverse effect of each basal media on follicular development. Its variation, in bovine, promotes the downregulation of certain proteins, which are responsible for the transportation of sugar that may result in intracellular glucose deprivation, instable energy reservoirs, and ultimately, can lead to oxidative stress (Fladeby et al., 2003).
Additionally, TCM-199 has higher concentration of few amino acids, such as arginine, glutamine, leucine, and tyrosine, in comparison to DMEM and Opti-MEM, which helps in regulating the intracellular pH and osmotic pressure. Our study also included a combination of some important factors such as EGF, ITS, FSH, FF, which maintains the morphology of preantral follicles. It impacts majorly, the proliferation of granulosa cells, deceleration in apoptosis, and promotion of antrum formation (Devesa and Caicedo, 2019).
So far, biological matrix is used frequently in follicular culture system, so the present study was performed to coculture isolated follicles with different types of somatic cells, where CC cocultured with follicles favored over OMC and BFFC. This could be explained as CC secretes certain factors, that is, oocyte growth-promoting factor, cumulus expansion-enabling factor, meiosis-activating factor, and few unknown proteins, which benefit the growth rate of follicles (Ramesh et al., 2010). Despite the positive effect of fibroblast cells on nuclear maturation of oocytes, its coculture with follicles could not induce cytoplasmic maturation in oocytes, which has reported to decrease the follicular development (Malekshah et al., 2014).
Besides the advantages of 2D culture system with somatic cells, which was considered to secrete certain factors that assist the growth of preantral follicles to the antrum formation, certain facts hinder its consistent development during the follicular growth. First, follicles attach and spread on to the surface that disrupt the 3D cell architecture, which is necessary for the exchange of gases and metabolic precursors to the oocyte and second, media will be consumed in case to maintain the somatic cells, which may slow down the growth of follicles. Moving on, extracellular matrix (ECM) is often being used for 3D encapsulation culture system, which promotes cell survival and proliferation over 2D coculture system.
During our study, follicles embedded in MAT showed significantly higher follicular growth percent as well as viability rate as compared with collagen and gelatin in all basal media types. This variation can be explained by the composition of the ECM used, Matrigel, a combination of laminin, collagen, and fibronectin, rich in protein, providing multilayer development, more competent than collagen and gelatin (a hydrolyzed collagen), having reduced protein fibrils that will lead to less integrity mediation adhesions. These comparisons clearly indicate the physical attributes of different matrices tailored, which improves the architecture of growing follicles.
Antioxidants, including CAT and SOD, are responsible for the improvement of cell viability potentially by reducing oxidative stress (Park et al., 2016). In the present study, CAT activity increases in the follicles, which showed a higher growth rate, similar to the reports in goat ovarian granulosa and theca cells during ovarian development (Behl and Pandey, 2002). This could be explained by the exceptional behavior of CAT in folliculogenesis that has been found to be different throughout estrous phases but its concentration in FF is not significantly different among different follicular stages (Shahat and Kandil, 2012). It is also evidenced that less SOD activity in FF is related to high rates of fertilization in humans (Sabatini et al., 1999), which is in similar pattern with our results where decreased SOD activity is observed in follicles with higher follicle size.
This decrease in SOD activity was necessary to ensure that ROS levels have reached threshold value in preovulatory follicles that may exert oxidative stress to oocytes. Therefore, development of follicles from primordial to antral stage involves several physiological activities which lead to a large amount of ROS production, inducing apoptosis process (Wang et al., 2017). But our results have shown a decreased ROS pattern, which may be due to the replenishment of media every alternate day that might have reduced the oxidative stress in the microenvironment of growing follicles.
Furthermore, LDH assay predicts the release of mitochondrial toxicants during apoptosis, but our results are showing an increase in LDH activity with increasing follicle size. This may be explained by an increase in carbohydrate metabolism, which utilizes lactate and pyruvate linked to the energy production during oocyte maturation (Fotakis and Timbrell, 2006).
In our study, induced E2 production was also observed with the accompanied follicular antrum formation, which also evaluates the effect of various gonadotropins and growth factors on differentiation of follicles. Intrafollicular endocrine and autocrine compounds, which contain some steroids, particularly estrogens, is an essential feature indicating the granulosa cell functionality during antral formation (Baufeld and Vanselow, 2018).
In conclusion to the present study, it formulates new information that correlates media composition with different matrices and the regulation of their growth. Individually, MAT encapsulation and CC coculture, provide the best environment for growing large preantral follicles along with TCM-199 basal media as compared with other combinations. The results of our study will contribute significantly to enhance the knowledge regarding the maintenance of microenvironment to get competent oocytes from the in vitro culture system of preantral follicles.
Footnotes
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This work was supported by the National Agriculture Science Fund (NASF/GTR-7004/2018-19/79), Indian Council of Agriculture Research, New Delhi and Department of Biotechnology (BT/PR8725/AAQ/1/556/2013), New Delhi.