
Abstract
Lymphedema is a chronic, progressive disease that causes pain as well as heavy economic burdens to patients. Reconstruction of the impaired lymphatic system is the key to treat lymphedema. Currently, there is no cure, but mesenchymal stromal cells show promising potential for lymphatic endothelial regeneration. Adipose-derived stromal cells (ADSCs) have been proved to support lymphangiogenesis both in vivo and in vitro. However, the mechanism in vascular endothelial growth factor C-induced (VEGF-C-induced) lymphatic endothelial transdifferentiation of ADSCs remains unknown. In this study, we show a novel link between the Wingless and int-1 (Wnt) pathway and the lymphatic endothelial differentiation process. We used LiCl to activate Wnt and DKK-1 to inhibit Wnt. Compared with the Wnt inhibition group and the control groups, the Wnt activation group produced more lymphatic endothelial cell (LEC)-related mRNA and proteins. Besides, Wnt-activated ADSCs formed longer tubes in two-dimensional culture and promoted the growth of lymphatic vessels in a three-dimensional transwell ADSC-LEC co-culture system. Our results demonstrated that activation of Wnt during the lymphatic endothelial transdifferentiation of ADSCs would enhance the efficacy of VEGF-C treatment. We anticipate our assay to expand our knowledge of Wnt in cell transdifferentiation and lay a foundation for future efforts to explore a novel and effective ADSC-based therapy for lymphedema.
Introduction
Lymphedema is a progressive chronic disease, caused by the destruction of lymphatic system (Rockson, 2001). Secondary lymphedema is often caused by lymphadenectomy, radiation, trauma, and infection (Deng et al., 2012; Szuba et al., 2002). Accumulation of protein-rich fluid, chronic inflammation, hypertrophy of adipose tissue, and fibrosis are common pathological changes in the swelling area (Bjork and Hettrick, 2019). Although several therapy strategies have been developed to reduce symptoms, there is no cure currently (Bjork and Hettrick, 2019). The surgical treatments have many shortages; in endovascular stent placement cases, displacement and deformation of the stents are widely seen (Raju et al., 2001), auto-transplantation of lymph nodes often cause damage to other parts of the body (Nguyen and Suami, 2015), and lymphovenous shunt operation is technically difficult, which could not be done without the help of microscope (Koshima et al., 2000). Thus, stimulating lymphatic regeneration by regenerative medicine seems the best way.
Mesenchymal stromal cells (MSCs) have long been recognized as a wonderful regenerative medicine (Sargent and Miller, 2016). Basic functions of MSCs are composed of cell differentiation, modulation of immune responses by secreting cytokines, and promotion of angiogenesis (Viswanathan et al., 2019). These properties make the MSCs literally appropriate for treating lymphedema. Fortunately, there have been some studies that strongly advocate MSC-based therapy is a promising treatment for lymphedema (Chen et al., 2020; Shimizu et al., 2012).
Bone marrow MSCs (BM-MSCs) and adipose-derived stromal cells (ADSCs) are two major sources of MSCs. ADSCs have attracted greater attention for the advantages of easy accessibility and easy to culture. In some studies, ADSCs have been proved to be of great potential to cure lymphedema: ADSCs support lymphangiogenesis in vitro by secreting vascular endothelial growth factor C (VEGF-C) and some other key factors (Ahmadzadeh et al., 2020; Strassburg et al., 2016); implantation of ADSCs could reduce lymphedema in a mouse secondary lymphedema model (Shimizu et al., 2012). However, recent studies showed ADSCs carry an intrinsic program that restricts their differentiation to adipogenic lineage cells (Chinnadurai et al., 2019; Roson-Burgo et al., 2016; Wagner et al., 2005). The swelling area of lymphedema patients is an adipogenic microenvironment, where ADSCs more easily differentiate into adipocytes (Levi et al., 2013; Xiang et al., 2020). This raises the concern that simple injections of ADSCs may aggravate adipose accumulation and further impair the lymphatic circulation.
Several studies have shown that pretreatment of MSCs before implantation could exhibit better therapeutic effects (Fan et al., 2012; Liu et al., 2015). Besides, under VEGF-C pretreatment, lymphatic endothelial cell (LEC)-related genes were upregulated significantly in the ADSCs (Yan et al., 2011; Yang et al., 2015). Therefore, inducing ADSCs toward LECs before transplantation might be a promising therapy strategy in the future. Understanding the mechanism of lymphatic endothelial differentiation of ADSCs is important for its future use in clinical therapy.
Wingless and int-1 (Wnt) signaling pathway is a highly conserved pathway in tissue development and differentiation (Nusse and Clevers, 2017); its role in angiogenesis is well-established (Dejana, 2010). Wnt controls vascular sprouting and remolding by integrating with Notch signaling (Phng et al., 2009), regulates human endothelial cell proliferation and migration (Cheng et al., 2008), and determines the endothelial progenitor cell fate through metabolic profile regulation (Shao et al., 2019). Activation of Wnt in ADSCs would promote angiogenesis in a ADSC-endothelial cell co-culture system (Cai et al., 2017). However, the role of Wnt in lymphatic development is poorly investigated.
In this study, we tried to explore how Wnt signaling regulates the lymphatic endothelial differentiation of ADSCs and lay a foundation for further cell-based therapy researches in vivo.
Materials and Methods
This animal experiment was registered and approved by the Ethics Committee of Sichuan University.
Cell culture
Adipose tissue was isolated from the inguinal white adipose tissue of postnatal 7-day wild-type mice. Harvested adipose tissue was digested by collagenase I (Sigma) for 1 hour and centrifuged at 3000 rpm for 5 minutes to separate the pellets. The pellets were suspended in a complete medium (α-MEM [minimum essential medium α] supplied 10% fetal bovine serum [FBS]). The medium was changed every other day; cells were passaged when 60%–80% confluent and used at passage 3. Drug concentration used in this experiment is as follows: VEGF-C (100 ng/mL, Proteintech), LiCl (0.5 mg/mL, Sigma), and DKK-1 (100 ng/mL, R&D).
Quantitative polymerase chain reaction
RNA was extracted from cultured ADSCs with TAKARA kit (Bio-medicals) according to the manufacturer's instructions. Nanodrop (Thermo) was used to determine the concentration and purity. PCR was performed in a Quant Studio 3 system (Applied Biosystems) and the SYBR Green (Qingke) was used as the detection dye. Relative expression levels were normalized against GAPDH transcript abundance. Primers are listed below (Table 1).
Primers Used in This Experiment
Western blot
The samples were collected at indicated times and then lysis with RIPA (Sigma-Aldrich). Then the extracts were run on 10% gels (Bio-Rad) and transferred to nitrocellulose membranes (Bio-Rad). After incubating with primary antibodies VEGFR-3 (ab273148, Abcam), PDPN (ab11936, Abcam), Prox-1(21409, SAB), Lyve-1 (49343, SAB), β-catenin (ab32572, Abcam), p-β-catenin (12114, SAB), and α-tubulin (C49624, SAB) at 4°C overnight, the membrane was incubated with secondary antibodies at room temperature for 1 hour. Chemiluminescence images were taken by a ChemiDoc™ MP (Bio-Rad).
Immunofluorescence staining
Cell immunofluorescence staining
Medium was removed and cells were washed with phosphate-buffered saline thrice; then the cells were fixed with 4% paraformaldehyde for 30 minutes. Then cells were permeabilized with 0.5% Triton x100 (T8200, Solarbio) and blocked with 2% bovine serum albumin (Thermo) for 30 minutes at room temperature. The cells were then incubated with primary antibodies β-catenin (ab32572, Abcam) and Lyve-1 (49343, SAB) overnight at 4°C followed by a 1-hour incubation with secondary antibody, 1-hour incubation with phalloidine (Thermo), and 1-minute incubation with 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI) at room temperature. Images were taken by a confocal microscope (N-STORM & A1, Nikon).
IF staining in the gel
We followed the steps as described before (Bruyere et al., 2008). The overall steps are the same with cell immunofluorescence (IF), but keep in mind that Matrigel is fragile and the melting point is 4°C, do not incubate below 4°C for too long.
Two-dimensional tube formation assay
First, we coated the 24-well plate with Matrigel (BD Bioscience), then the gel was allowed to polymerize at 37°C for 30 minutes. After induction for 7 days, the ADSCs were seeded on the gel with a concentration of 1 × 105 per well. Images were taken 12 hours after seeding. The total length of the tubes was measured by Image-Pro (Media Cybernetics), five fields were randomly selected for each well, and mean length was calculated.
Three-dimensional tube formation assay
In this experiment, we used the transwell apparatus (Corning) to investigate how ADSCs affect lymphatic vessels growing in a paracrine way. After 7 days of induction, ADSCs were seeded in the upper chamber and 1-mm pieces of thoracic duct were embedded in the Matrigel in the lower chamber. The medium in the upper chamber was α-MEM supplemented with 10% FBS and the lower chamber was MCDB131 supplemented with 10% FBS. The thoracic duct was provided by the State Key Laboratory of Oral Diseases, Sichuan University. The diameter of vessel occupation area was used to evaluate the tube growth. The identity of the tube network was identified by IF (Fig. 1).

Thoracic duct rings were embedded in Matrigel and cultured for 7 days, then the branches were identified as lymphatic vessel by IF. Lyve-1 is a specific LEC marker. IF, immunofluorescence; LEC, lymphatic endothelial cell. Color images are available online.
Statistical analysis
Data were presented as mean ± standard deviation. Independent sample t-test was performed to compare the difference using SPSS 16.0 (Chicago, IL). Differences with p < 0.05 were considered statistically significant.
Results
VEGF-C successfully induced lymphatic endothelial transdifferentiation of ADSCs
Primary ADSCs were isolated from postnatal 7-day C57/B6 mice, passage 3 ADSCs were used in this experiment, ADSCs showed typical spindle-like morphology, and osteogenic and adipogenic differentiation ability were confirmed by alizarin red staining and oil red staining (Fig. 2a). To test the possibility that Wnt is activated during lymphatic endothelial differentiation of ADSCs, we performed this experiment. To do this, first, we tested whether VEGF-C (100 ng/mL) alone could induce the lymphatic endothelial differentiation of ADSCs. Using IF staining, we recognized that an LEC-related marker lyve-1 started to emerge after 4 days of induction (Fig. 2b). Consistent with these results, we then used q-PCR to confirm the upregulation of LEC-specific genes, LYVE1, VEGFR3, PROX1, and PDPN (Fig. 2c–f). Western blot further confirmed this (Fig. 2g).
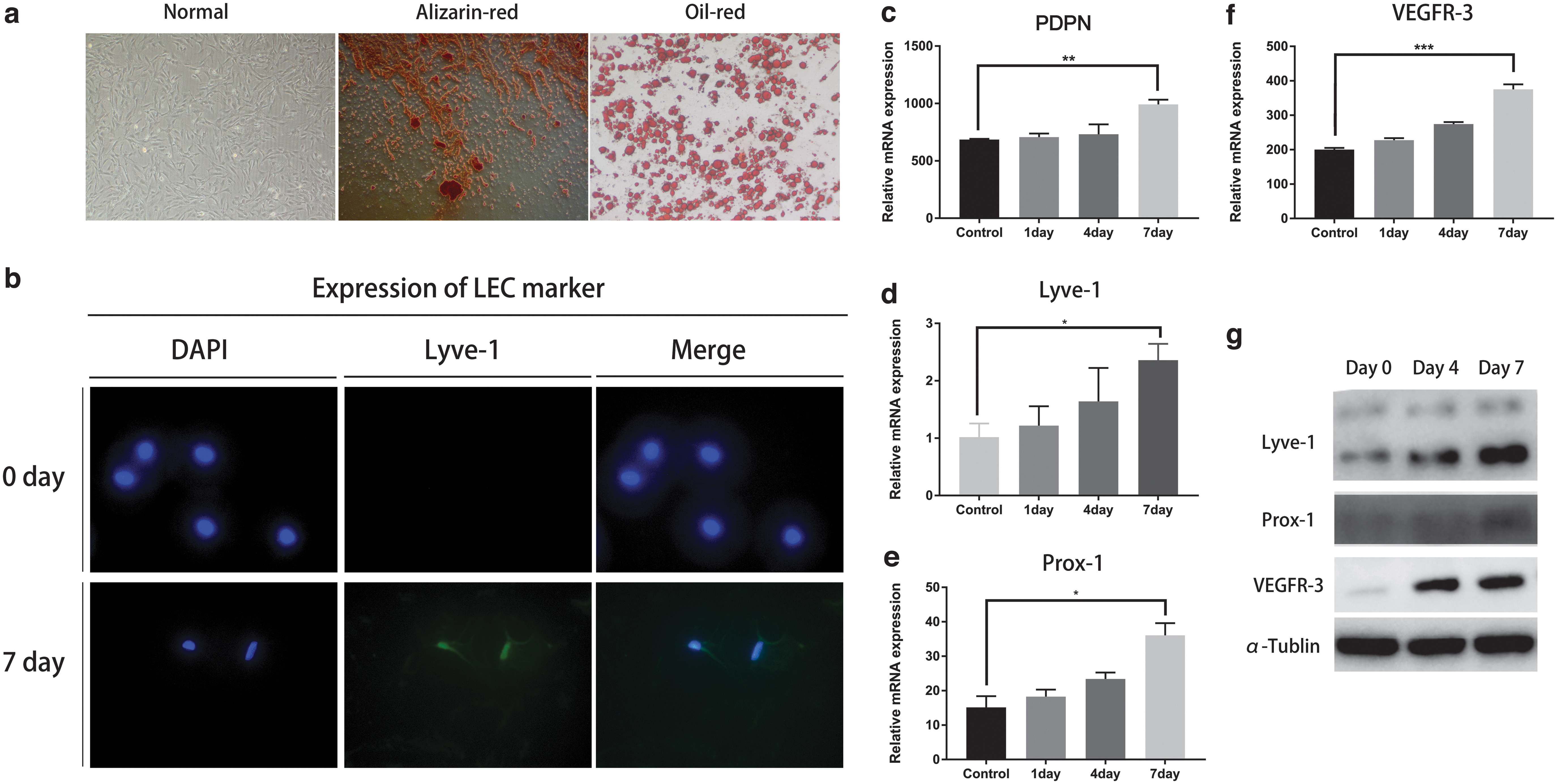
Successful lymphatic endothelial transdifferentiation of ADSCs. Isolated ADSCs were successfully induced to lymphatic endothelium differentiation under the complete medium supplemented with 100 ng/mL of VEGF-C for a week, cells were collected at the indicated times. LYVE1, PROX1, PDPN, and VEGFR3 are LEC-specific markers.
Canonical Wnt pathway was activated during the lymphatic endothelial transdifferentiation of ADSCs
Next, we explored whether Wnt was activated during the transdifferentiation. Since the translocation of β-catenin into nucleus is a symbol for Wnt activation, we examined the localization of β-catenin by IF. DAPI-stained nucleus and F-actin represented cytoplasm. Before induction, β-catenin distributed in the cytoplasm and nucleus equally; during the next 12 hours, β-catenin accumulated in the nucleus (Fig. 3a). Western blot further confirmed this (Fig. 3b). These data supported our hypothesis that Wnt was activated during lymphatic endothelial transdifferentiation of ADSCs.
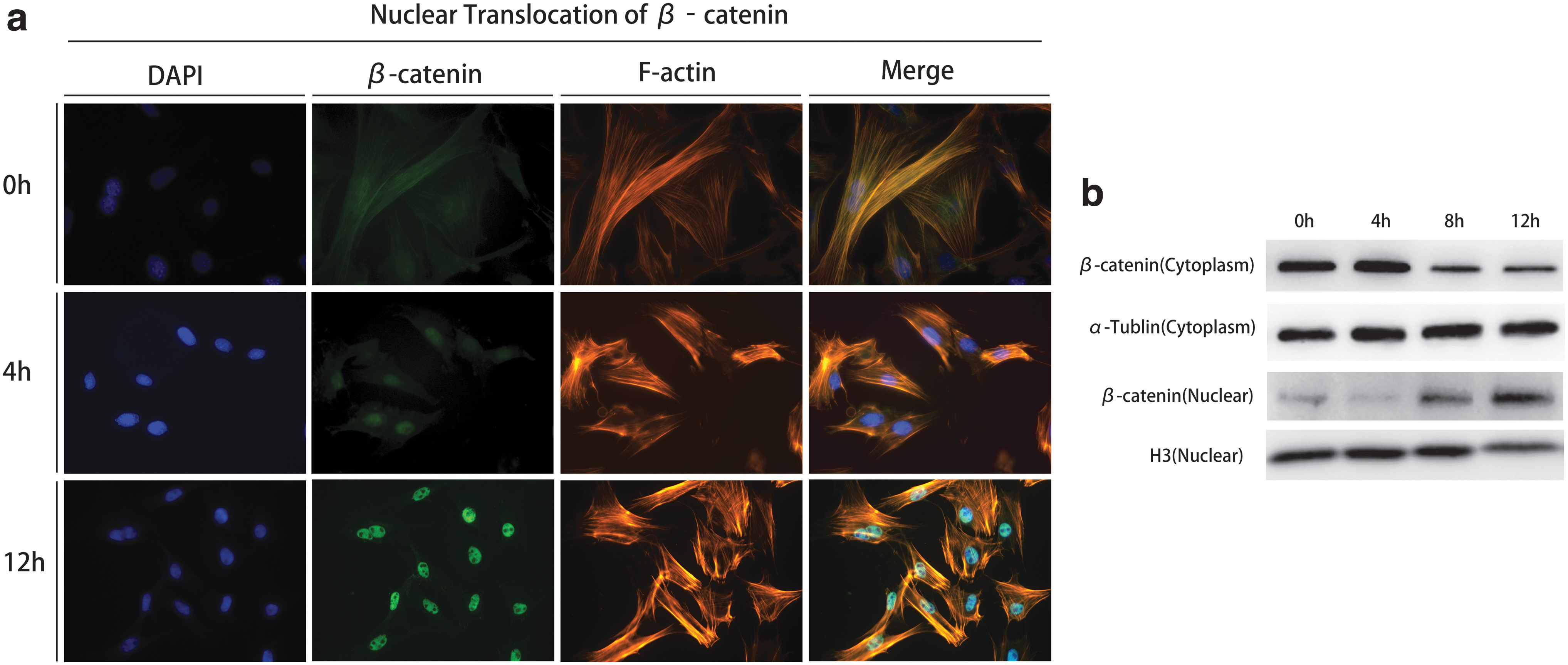
Nuclear translocation of β-catenin during the lymphatic endothelial transdifferentiation of ADSCs.
Effect of LiCl and DKK-1 on Wnt manipulation
LiCl and DKK-1 are well-established drugs to manipulate Wnt signaling in ADSCs (Cai et al., 2017). So we tried to use them to manipulate Wnt in this experiment. First, we confirmed LiCl could activate Wnt and DKK-1 could inhibit Wnt in ADSCs. The ratio of β-catenin/p-β-catenin represents the activation level of Wnt signaling. Our results showed a significant increase of β-catenin/p-β-catenin ratio in LiCl group and significant decrease in DKK-1 group compared with the control group (Fig. 4a). Then we tested whether LiCl and DKK-1 have the same influence in the presence of VEGF-C. Western blot showed VEGF-C+LiCl group produced significantly more β-catenin accumulation and more Wnt downstream proteins Lef1 and CyclinD1 than the other groups (Fig. 4b). These results verified our hypothesis that LiCl treatment could activate Wnt and DKK-1 treatment could suppress Wnt during the lymphatic endothelial transdifferentiation of ADSCs.
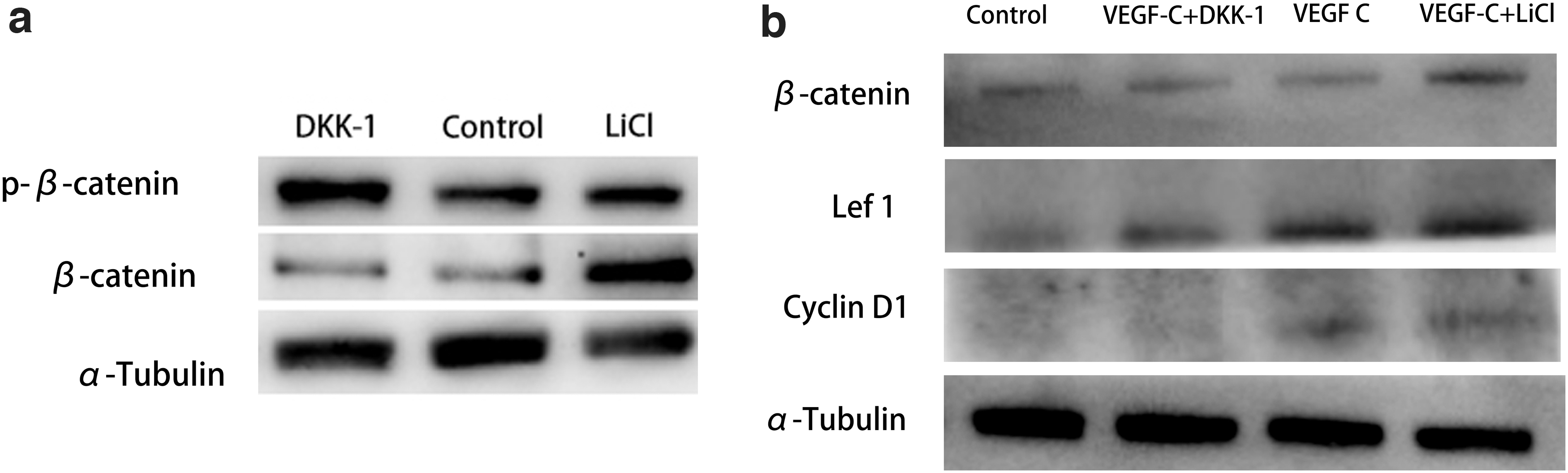
Effect of LiCl and DKK-1 on Wnt manipulation.
Effect of Wnt manipulation on the key factors during the lymphatic endothelium transdifferentiation of ADSCs
Base on the data, we the hypothesized that activation of Wnt signaling could accelerate the transdifferentiation process. To do this, we used quantitative polymerase chain reaction (qPCR) and Western blot to detect the protein expression levels of LEC-related genes. Overall, Western blot showed the VEGF-C+LiCl group produced the most LYVE1, PDPN, PROX1, and VEGFR-3 at 7 days postinduction. However, at 7 days, the accumulation of PROX1 seemed close between the VEGF-C+LiCl group and the VEGF-C group, although the VEGF-C+LiCl group produced most PROX1 at 4 days (Fig. 5e).

Effect of Wnt manipulation on the expression of LEC-related markers.
Consistent with these results, we then used qPCR to investigate the gene transcription levels of the LEC-related genes (Fig. 3a–d). At 7 days, compared with the DKK-1+VEGF-C group, mRNA expression levels of LYVE1, PDPN, PROX1, and VEGFR-3 increased by 137%, 199%, 490%, and 58%, respectively, in the VEGF-C+LiCl group. However, when comparing the VEGF-C+LiCl group and the VEGF-C group, there were no significant differences in the mRNA expression level of Pod at 7 days postinduction. This is in coincidence with the Western blot results. These data suggested that activation of Wnt could accelerate lymphatic endothelium differentiation of ADSCs and produce more LEC-related proteins and mRNA.
Effect of Wnt manipulation on lymphangiogenesis of lymphatic endothelial trandifferentiated ADSCs in vitro
Lymphangiogenesis, in other words: the ability to form tubes and promote the growth of LECs, is the golden standard to evaluate the success of lymphatic endothelial transdifferentiation of ADSCs.
To determine the effect of Wnt on lymphangiogenesis, we performed tube formation assays. To do this, we designed two-dimensional (2D) culture assay to test the tube formation activity of ADSCs, and three-dimensional (3D) transwell co-culture with LECs to assess how ADSCs stimulate the growth of LECs in a paracrine-dependent way. The network in the gel was composed of lymphatic vessels (Fig. 1). For 2D culture, after 7 days of induction, 1 × 105 cells were seeded on the Matrigel, and 12 hours later, images were taken (Fig. 6a). VEGF-C+LiCl-treated ADSCs formed many more tubes than the other groups (Fig. 6b). In the 3D transwell co-culture, we seeded ADSCs in the upper chamber and LECs (primary thoracic duct rings) in the lower chamber. There were no ADSCs in the VEGF-C group and the LEC group, the VEGF-C group was selected as the positive control and the LEC group was the negative control. The diameter of vessel occupation area was used to evaluate the lymphangiogenic ability.
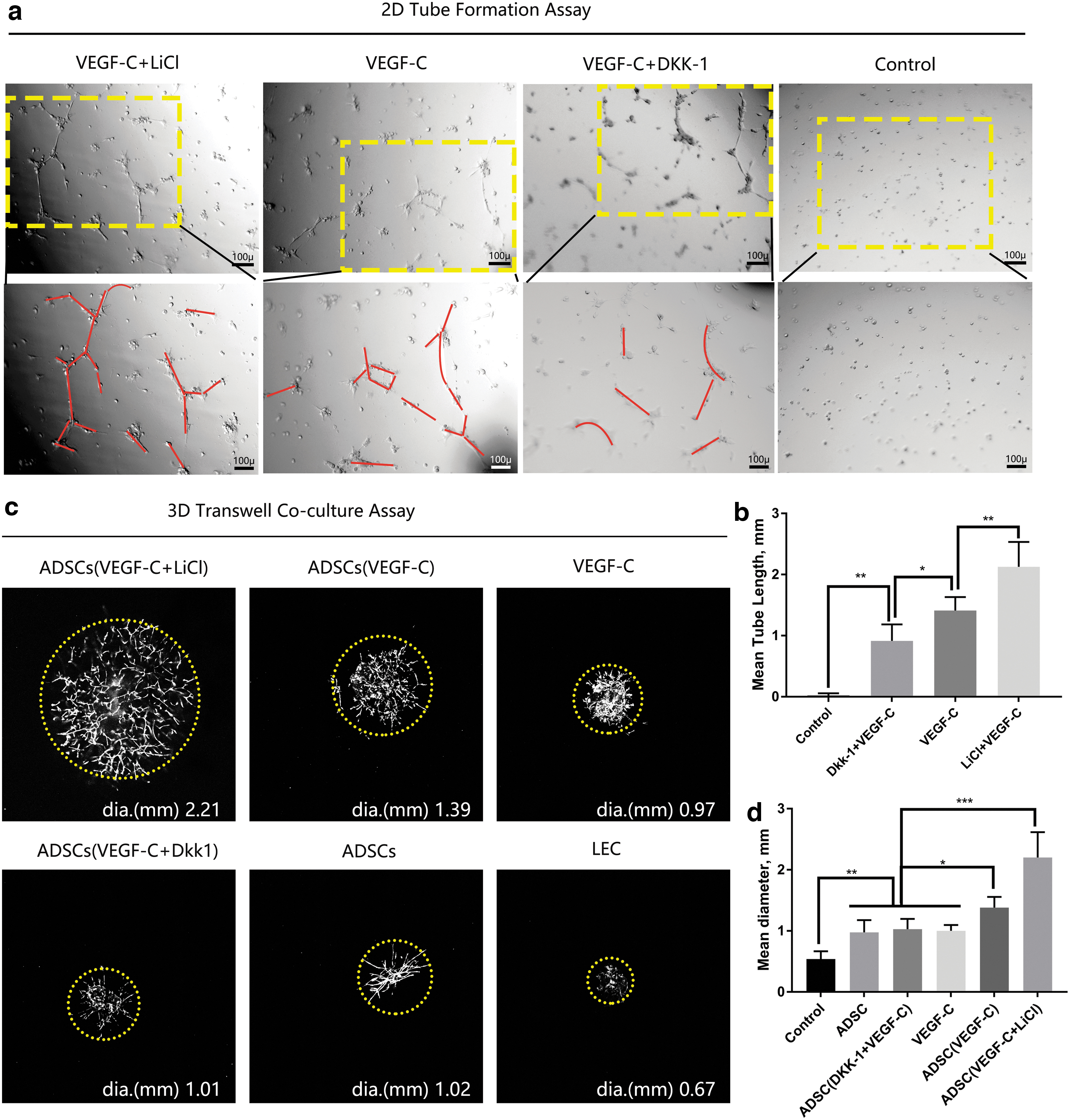
Effect of Wnt manipulation on tube formation assays. For 2D culture, after 7 days of induction, ADSCs were seeded on Matrigel, tube formation was evaluated at 12 hours postseeding. For 3D culture, we used transwell apparatus, induced ADSCs, were seeded in the upper chambers, Matrigel containing a small fraction of thoracic duct was placed in the lower chambers, images were taken 7 days later.
In the VEGF-C+LiCl-treated ADSC group, the lymphatic sprouts were longer and more disorganized (Fig. 6c). Comprehensively analysis (Fig. 6d) showed there were no significant differences among the ADSC group, the ADSC (VEGF-C+DKK-1) group, and VEGF-C group. Taken these data together, Wnt activation during the lymphatic endothelial transdifferentiation of ADSCs not only promoted tube formation ability of ADSCs but also promoted the growth of LECs in a paracrine way.
Discussion
Secondary lymphedema is frequently seen after lymph node dissection to treat head and neck cancer
For a long time, the accumulation of interstitial fluid caused by the destruction of lymphatic vessels is considered the primary problem. However, recent studies have shown that scar tissue progressively replaces functional parenchyma and wraps lymphatic vessels in the lymphedema region. As a result, the lymphatic system is replaced by excessive deposition of fibroadipose tissue, which also contributes significantly to the development of this disease (Mihara et al., 2012). So the ideal medicine for treating lymphedema should possess the two abilities: promoting the growth of existing lymphatic vessels and attenuating fibrosis.
The MSC therapy seemed promising for their potential to modulate the immune response and secrete growth factors (Ahmadzadeh et al., 2020; Regmi et al., 2019). Several clinical trials also showed MSC treatment significantly reduced the volume and decreased symptomatology (Hou et al., 2008; Maldonado et al., 2011; Toyserkani et al., 2016). However, the mechanisms by which MSCs therapy may function were unclear until most recently.
Ogino et al. (2020) found ADSC transplantation could recover the scar fibers, accelerate LEC proliferation, and produce more lymphatic vessels. Especially, they observed the intussusceptive lymphangiogenesis and administration of ADSCs promoted this process (Ogino et al., 2020). Intussusceptive lymphangiogenesis is a complementary form of classic sprouting lymphangiogenesis. In this kind of lymphangiogenesis, opposing endothelial cells protrude into the capillary and create separated zones (Diaz-Flores et al., 2019).
In short, intussusceptive lymphangiogenesis creates more branches, while sprouting lymphangiogenesis creates longer vessels (Konerding et al., 2010). The advantage of intussusceptive lymphangiogenesis lies in expanding the lymphatic network quickly with minimal cell proliferation. However, in Ogino's research, the authors also observed the proliferation of LECs, indicating the event of sprouting lymphangiogenesis (Ogino et al., 2020). This is also in accordance with our 3D lymphangiogenesis data. So we can reasonably hypothesize that both sprouting and intussusceptive lymphangiogenesis take place during the lymphedema treatment, but which one dominates the repairing process needs further research.
In recent years, the concept “preprogrammed MSCs” has attracted great attention, which means the MSCs isolated from specific tissue carry an intrinsic differentiation program to develop into the original tissue-specific cell types (Gokbuget and Blelloch, 2019; Rauch et al., 2019; Yianni and Sharpe, 2019, 2020). For example, BM-MSCs are more likely to differentiate into osteogenic lineage cells, while ADSCs are more likely to differentiate into adipocytes (Mohamed-Ahmed et al., 2018; Shen et al., 2019). Therefore, when we consider ADSC therapy outside making adipose, we need to ensure that the adipogenic preprograms are changed into new differentiation programs. Precious studies have suggested VEGF-C induction as a well-established way to guide lymphatic endothelial transdifferentiation of MSCs (Wang and Xue, 2019; Yan et al., 2011). However, the mechanism in this transdifferentiation process is poorly studied.
In this research, we found that Wnt signaling pathway regulated the lymphatic endothelial transdifferentiation of ADSCs, and activation of Wnt could accelerate this process. β-catenin is the key protein in Wnt signaling pathway. It appears a phosphorylated state (p-β-catenin) and resides along cell skeleton within the cytosol. Upon Wnt activation, the p-β-catenin was dephosphorylated into β-catenin, followed by translocation into the nucleus (Nusse and Clevers, 2017). In this experiment, we observed the neural translocation of β-catenin happened very quickly, only several hours after VEGF-C induction, indicating Wnt pathway was the trigger by VEGF-C directly.
Then we tried to manipulate Wnt with LiCl and DKK-1. Luckily, both of them worked well in the presence of VEGF-C, VEGF-C+LiCl-upregulated Wnt signaling, and VEGF-C+DKK-1-downregulated Wnt. Molecularly, activation of Wnt signaling pathway during the induction could result in more VEGFR-3, Lyve-1, Prox-1, and Pod, while inhibition of Wnt resulted in significantly less. Then we further investigated the effect of Wnt manipulation on lymphangiogenesis. When we consider the transdifferentiation of ADSCs, the most important is whether the induced ADSCs provide specific lymphangiogenesis functions. So we designed 2D tube formation assay and 3D co-culture assay to assess. Phenotypically, Wnt-activated ADSCs formed longer tubes and promoted the growth of lymphatic vessels than the other groups, while Wnt-inhibited ADSCs formed shorter tubes as well as lacked the ability to promote the growth of lymphatic vessels.
Taken these data together, we concluded that Wnt signaling pathway regulated the lymphatic endothelial transdifferentiation of ADSCs and activation of Wnt during the transdifferentiation promoted lymphangiogenesis in vitro.
Footnotes
Acknowledgments
The research was performed at the State Key laboratory of Oral Disease (State Key Laboratory of Oral Diseases, National Clinical Research Center for Oral Diseases, Department of Oral and Maxillofacial Surgery, and West China Hospital of Stomatology, Sichuan University, No. 14, Section 3, South Renmin Road, Chengdu, 610041, China).
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This research is supported by Research and Develop Program, West China Hospital of Stomatology Sichuan University, No. LCYJ2019-1, and Department of Human Resources and Social Security of Sichuan Province, No. 2017-A.