
Abstract
In our previous study, we constructed Schwann cells (SCs) that stably express Simian virus 40 T antigen (SV40T-SCs). SV40T-SCs functions and markers are similar to those of neural crest cells. There we used bone morphogenetic protein 9 (BMP9) to induce SV40T-SCs differentiation in vitro and in vivo and study possible related mechanism. SV40T-SCs differentiation was induced by BMP9 conditioned medium. The lipogenic differentiation of SV40T-SCs was assessed by Oil Red O staining. Alizarin red and Alcian blue staining, and alkaline phosphatase (ALP) assays were used to evaluate the SV40T-SCs osteogenic differentiation. The expression of adipocyte differentiation (c/EBPα and c/EBPβ) and osteoblast differentiation markers (OSX and RUNX2) were detected by quantitative polymerase chain reaction (qPCR). To study possible mechanism related to SV40T-SCs differentiation, the P53 and E2F1 activity were assessed by luciferase reporter plasmid, and Slug and E-cadherin expression by qPCR. In vivo, SV40T-SCs infected by Ad-BMP9 or Ad-GFP were injected under the skin of nude mice. After 4–6 W, the mice were euthanized and subcutaneously mass formed at injecting sites was collected for pathological analysis. After SV40T-SCs were cultured in BMP9 conditioned medium, lipid droplets were formed in the cytoplasm of these cells. Alizarin red and Alcian blue staining were positive, and ALP activity of SV40T-SCs increased significantly. The expression of adipocyte differentiation (c/EBPα and c/EBPβ) and osteoblast differentiation markers (OSX and RUNX2) in SV40T-SCs was upregulated by BMP9. SV40T significantly increased Slug expression and decreased E-cadherin expression. SV40T-SCs infected with Ad-BMP9 were able to differentiate into adipose tissue and form a small bone matrix under the nude mice skin. SV40T-SCs have the ability to differentiate into adipocytes and osteoblasts in vivo and in vitro. SV40T can upregulate the Slug expression and downregulate the E-cadherin expression to produce endothelial-to-mesenchymal transition (EMT). The multidirectional differentiation ability of SV40T-SCs may be related to EMT.
Introduction
Bone morphogenetic proteins (BMPs) are members of the TGFβ superfamily and play an important role in the stem cell proliferation and differentiation during development (Varga and Wrana, 2005; Zhang and Li, 2005). The deletions of BMPs results in skeletal and extraskeletal development abnormalities (Zhao, 2003). Several BMPs have been shown to regulate the osteoblast differentiation of mesenchymal stem cells (MSCs) (Deng et al., 2008; Hogan, 1996). In our previous study, we established a cell line that stably expresses a high level of active BMP9. This protein was shown to effectively induce osteogenic differentiation of MSCs (Li et al., 2016). Another study has shown that BMP9 has the ability to induce osteogenic and adipogenesis differentiation of reversibly immortalized stem cells from the dental apical papilla (Wang et al., 2014).
Schwann cells (SCs) in peripheral nerve sheaths of mice, as terminal cells for neural differentiation, have no differentiation ability. Previously, we constructed SCs that stably express Simian virus 40 T antigen (SV40T), which we named as SV40T-SCs. In this study, we observed the occurrence of significant adipogenesis and osteogenesis after culturing SV40T-SCs with BMP9 in vitro and in vivo. These results suggest that SV40T-SCs achieve transdermal differentiation, or transdifferentiation, and that SV40T induces SCs to be able to differentiate. Thus, we concluded that a reprogramming occurred in SV40T-SCs and these cells acquired differentiation ability similar to stem cells. Reprogramming can induce differentiated cells to revert to pluripotency cells, which in turn can differentiate into almost any cell type (Jopling et al., 2011).
Materials and Methods
Cell culture, chemicals, and animals
Primary SCs were obtained from the American Type Culture Collection (ATCC; Manassas, VA). All of cell lines were maintained in complete Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS, Hyclone, Logan, UT), 100 U/mL penicillin, and 100 μg/mL streptomycin at 37°C in 5% CO2. BMP9 conditioned medium (BMP9-cm) was prepared according to the description of our previous study (Li et al., 2016). Unless otherwise stated, all chemicals were purchased from Sigma-Aldrich (St. Louis, MO) or Thermo Fisher Scientific (Pittsburgh, PA). Sprague Dawley (SD) nude mice were purchased from the Animal Laboratory Center of the University of Chicago. Animal welfare, use, and care were carried out in accordance with the protocol approved by the Institutional Animal Care and Use Committee (IACUC) of the University of Chicago (protocol #71108).
Establishment of SV40T-SCs
To establish SV40T-SCs, early passage SCs (3–5 passages) were seeded in 25 cm2 flasks and cotransfected with piggyBac vector MPH86, which expresses SV40T flanked with FLP (Fig. 1A), piggyBac transposase expression vector, and Super PiggyBac (SBI, Mountain View, CA). SV40T-SCs were established by selecting the transfected cells with hygromycin B for 1 week. SV40T-SCs aliquots were preserved in liquid nitrogen tanks. Subsequently, SV40T-SCs were divided into two groups. The Ad-Flip group was infected with recombinant Flip adenovirus for 36 hours, whereas the Ad-GFP group was infected with recombinant adenovirus encoding green fluorescence protein (GFP) for 36 hours. SV40T expression was detected by quantitative reverse transcription polymerase chain reaction (qRT-PCR).
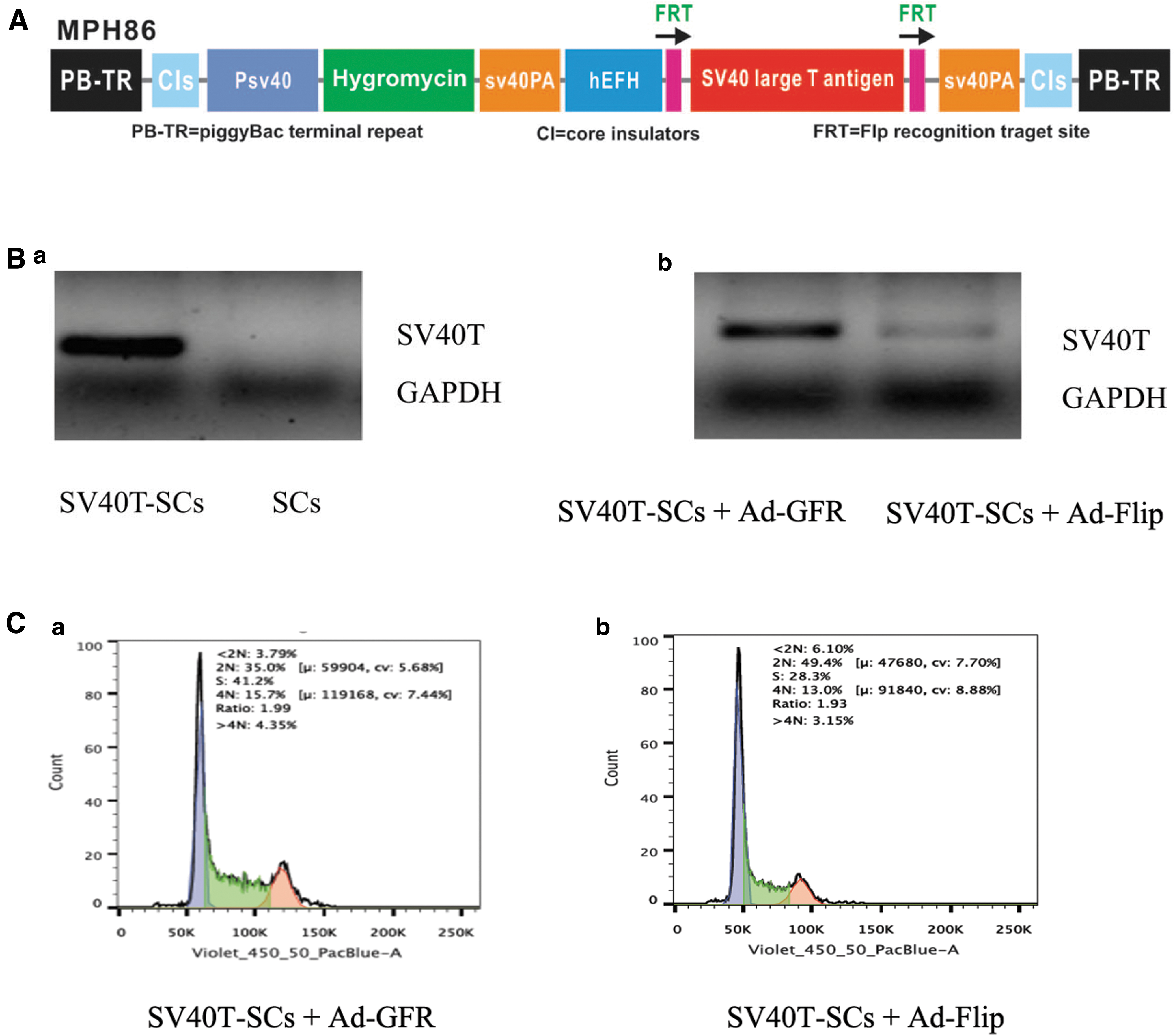
The piggyBac transposon-mediated SV40T expression effectively established SV40T-SCs and their cell cycle exhibits significant changes in comparison with primary SCs.
In brief, total RNA was isolated using TRIZOL reagents (Invitrogen, Carlsbad, CA) and subjected to the reverse transcription reaction with hexamer and M-MuLV Reverse Transcriptase (New England Biolabs, Ipswich, MA). The cDNA products were diluted 10–100 times and used as polymerase chain reaction (PCR) templates. The primers are shown in Table 1. PCR was performed as follows: 94°C for 2 minutes for 1 cycle; 92°C for 20 seconds, 68°C for 30 seconds, and 72°C for 12 cycles with a decrease in 1°C per cycle; and then at 92°C for 20 seconds, 57°C for 30 seconds, and 72°C for 20 seconds for 20–25 cycles, depending on the transcript abundance. All samples were normalized by the GAPDH expression level.
Quantitative Polymerase Chain Reaction (qPCR) Primer Sequences
Fluorescence activating cell sorter analysis
The cell cycle of SV40T-SCs in the Ad-Flip and Ad-GFP groups was detected by Fluorescence Activating Cell Sorter. The cells were harvested, fixed with 70% ethanol, washed with PBS, and stained with Hoechst 33342. The cell cycle was analyzed using the BD LSR II Flow Cytometer and the FlowJo software. All assays were performed in triplicate.
BMP9-induced differentiation of SV40T-SCs in vitro
In the BMP9 group, SV40T-SCs were cultured in 25% BMP9-cm, whereas in the control group SV40T-SCs were cultured in HEK293 medium that did not contain BMP9. Morphological changes in the cells were observed daily under a light microscope.
Evaluation of lipogenic differentiation in vitro
Red-Oil-O staining was used to assess SV40T-SCs lipid formation. In brief, cells were fixed with 10% formalin at room temperature for 20 minutes and washed with PBS. Then, the fixed cells were stained with freshly prepared Oil Red O solution (six parts saturated Oil Red O dye in isopropanol plus four parts water) at 37°C for 30–60 minutes, followed by washing with 70% ethanol and distilled water.
Evaluation of osteogenic differentiation in vitro
SV40T-SC osteogenic differentiation was evaluated by Alizarin red S stains and Alcian blue staining, and by alkaline phosphatase (ALP) assays. For Alizarin red S staining, cells were fixed with 0.05% (vol/vol) glutaraldehyde at room temperature for 10 minutes, washed with distilled water, incubated with 0.4% Alizarin red S for 5 minutes, and washed again with distilled water. The cells were then observed under a light microscope. For Alcian blue staining, the cells were washed with PBS, fixed with 4% formaldehyde solution for 30 minutes and washed again with PBS, stained with Alcian blue solution for 30 minutes and washed with distilled water. The cells were then observed under an optical microscope.
For ALP assays, SV40T-SCs seeded in 24-well culture plates were treated with 25% BMP9-cm or control medium. ALP activity was assessed qualitatively. At the indicated time points, ALP activity was measured quantitatively using the modified Great Escape SEAP Chemiluminescence assay kit (BD Clontech) and qualitatively with histochemical staining assay (using a mixture of 0.1 mg/mL naphthol AS-MX phosphate and 0.6 mg/mL Fast Blue BB salt). Each assay condition was performed in triplicate and repeated in three independent experiments. ALP activity was normalized by total cellular protein concentrations in all samples.
Detection of the adipogenic and osteogenic differentiation genes expression
After 3 days for differentiation in vitro, adipocyte differentiation was evaluated by the c/EBPα and c/EBPβ expression levels detected by qPCR, and the osteoblast differentiation by the OSX and RUNX2 expression levels also detected by qPCR. In brief, total RNA was isolated by using TRIZOL Reagents (Invitrogen, Carlsbad, CA) and subjected to reverse transcription with hexamer and M-MuLV reverse transcriptase (New England Biolabs). The cDNA products were used as PCR templates. qPCR primers were designed with the Primer3 program for the genes of interest. The primers are shown in Table 1. SYBR Green-based PCR analysis was carried out by using CFX-96 Connect (Bio-Rad, Hercules, CA). All PCRs were done in triplicate. GAPDH was used as a reference gene.
P53 and E2F1 activity detection by luciferase reporter assay
The P53 gene sequence was inserted into the p-MIR-reporter plasmid. For the luciferase reporter assays, SV40T-SCs were cultured in 24-well plates, and each well was transfected with 0.4 μg of P53 firefly luciferase reporter plasmid, 0.4 μg of E2F1 firefly luciferase reporter plasmid, or 0.4 μg of a β-galactosidase (β-gal) expression plasmid (Ambion) using Lipofectamine 3000 (Invitrogen). The β-gal plasmid was used as a transfection control. Twenty-four hours after transfection, cells were evaluated using a luciferase assay kit (Promega, Madison, WI). Subsequently, SV40T-SCs were divided into two groups. The experimental group was infected with Ad-Flip for 24 hours and the control group was infected with Ad-GFP for 24 hours. Then detection of P53 and E2F1 activity was detected by luciferase reporter assay.
Slug and E-cadherin expression detection
Slug and E-cadherin expression was detected by qPCR. In brief, total RNA was isolated by using TRIZOL Reagents and subjected to reverse transcription with hexamer and M-MuLV reverse transcriptase. The cDNA products were used as PCR templates. qPCR primers were designed with the Primer3 program for the genes of interest. The primers are also shown in Table 1. SYBR Green-based PCR analysis was carried out by using CFX-96 Connect. All PCRs were performed in triplicate and GAPDH was used as an internal reference.
In vivo differentiation assessment
SV40T-SCs were infected by Ad-Bmp9 or Ad-GFP. After 24 hours, the cells suspension density was 1 × 107. Two hundred microliters of Ad-BMP9-infected SV40T-SCs (experimental group) or Ad-GFP-infected SV40T-SCs (control group) were injected per point under the skin of male nude mice. The lumps on the back of mice were observed regularly. After feeding for 4–8 W, nude mice were euthanized. Differentiation of SV40T-SCs in the subcutaneous environment of nude mice was observed by hematoxylin and eosin (HE) and Alizarin red S staining.
Statistical analysis
All quantitative experiments were carried out in triplicate and/or repeated three times. The data were expressed as mean ± standard deviation. Statistical analysis was performed by one-way analysis of variance and the Student's t-test. p-Values <0.05 were considered statistically significant.
Results
PiggyBac transposon-mediated SV40 T expression effectively established SV40T-SCs and their cell cycle exhibits significant change in comparison with primary SCs
Cells cotransfected with the piggyBac vector MPH86 expressed SV40T, but primary SCs did not (Fig. 1B, panel a). This indicated that the transfection was successful. SV40T expression in the Ad-Flip group was significantly decreased, indicating that Ad-Flip can knock out the SV40T expression (Fig. 1B, panel b). The cell cycle distribution was detected by flow cytometry. In the Ad-GFP group 41.2% of the cells were found in the S-phase cells and 35.0% in the 2N-phase cells, whereas in the primary SCs 28.3% of the cells were found in S-phase and 49.4% in the 2N-phase (Fig. 1C, panel a). After the SV40T expression knockout by Flip adenovirus, the cell cycle was blocked in G1 phase, and the cell number that entered S phase decreased significantly (Fig. 1C, panel b). Therefore, SV40T can promote cell passage from G1 to S phase and accelerate the cell cycle.
Bmp9-induced lipogenic differentiation of SV40T-SCs in vitro
In the BMP9 group, oil droplets gradually appeared in the SV40T-SCs cytoplasm and were noticed significantly on third day. The oil droplets gradually fused and enlarged while the cell body gradually became round, without adhering to the wall. The cells were suspended in the culture medium, and became mature adipocytes. Red-oil-0 staining is shown in Figure 2B and C. BMP9 alone induced SV40T-SCs to differentiate into adipocytes in vitro. There were no oil droplets in the control group (Fig. 2A).
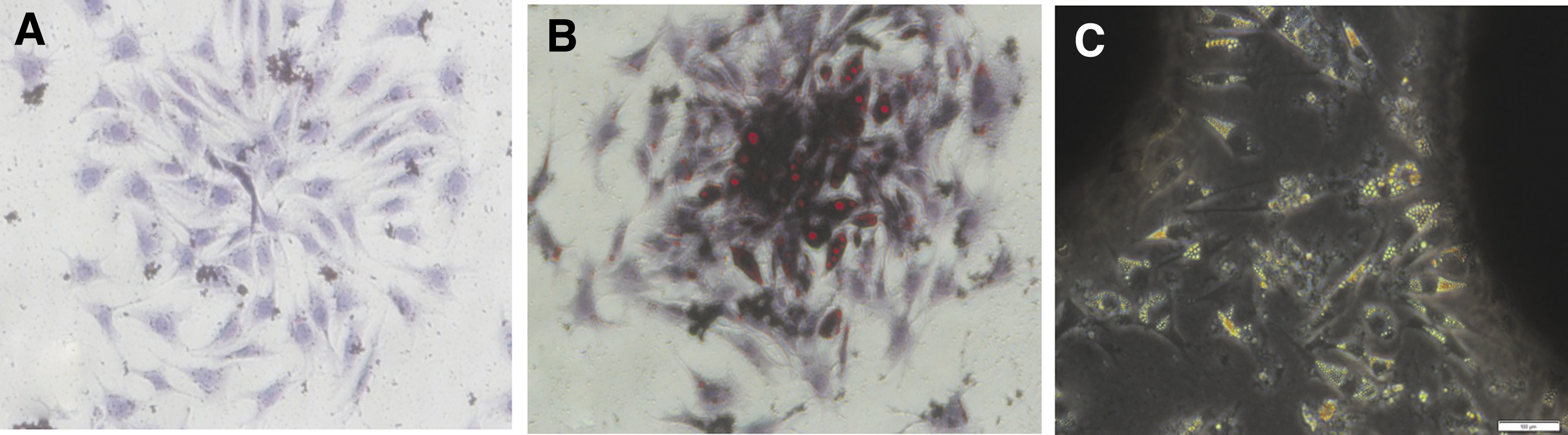
BMP9-induced lipogenic differentiation of SV40T-SCs in vitro.
Bmp9-induced osteogenic differentiation in vitro
SV40T-SCs were cultured in BMP9-cm for 5 days. Alizarin red staining was positive for calcium deposition (Fig. 3A) and Alcian blue staining was positive for collagen formation (Fig. 3B). ALP activity in BMP9 group was significantly increased compared with the control group. ALP activity in BMP9 group increased significantly on the fifth day compared with the second day (Fig. 3C).
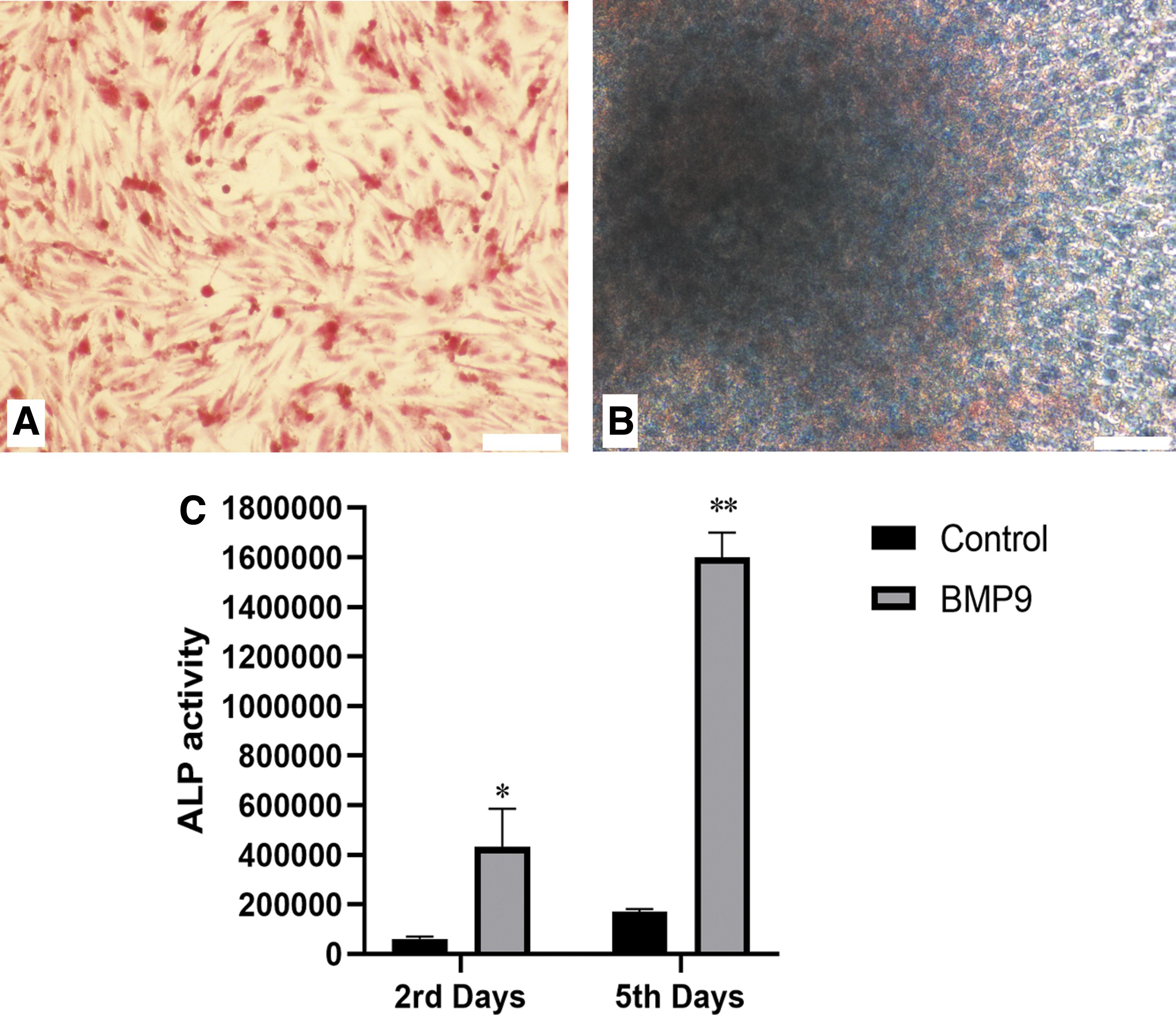
BMP9-induced osteogenic differentiation of SV40T-SCs in vitro.
Adipogenic and osteogenic differentiation genes expression
After cultivating SV40T-SCs in BMP9-cm for 3 days, the expression of c/EBPalpha, c/EBPbeta, OSX, and RUNX2 were significantly higher than in the control group (Fig. 4).

Adipogenic and osteogenic differentiation genes expression. The expression of c/EBPα, c/EBPβ, OSX, and RUNX2, and increased significantly in the SV40T-SCs cultured in BMP9 conditioned medium. *p < 0.05, **p < 0.01, ***p < 0.001 (vs. control groups).
P53 and E2F1 activity, and slug and E-cadherin gene expression
Luciferase reporter plasmid was used to detect P53 and E2F1 activity 24 hours after infection. After SV40T expression knockout in SV40T-SCs by Flip adenovirus, P53 activity increased and E2F1 activity decreased significantly (Fig. 5A). These results suggest that SV40T can inhibit P53 activity and enhance E2F1 activity. Slug expression decreased and E-cadherin expression increased significantly in Ad-Flip group compared with the Ad-GFP group (Fig. 5B). These data indicated that SV40T can upregulate the Slug expression and downregulate the E-cadherin expression.
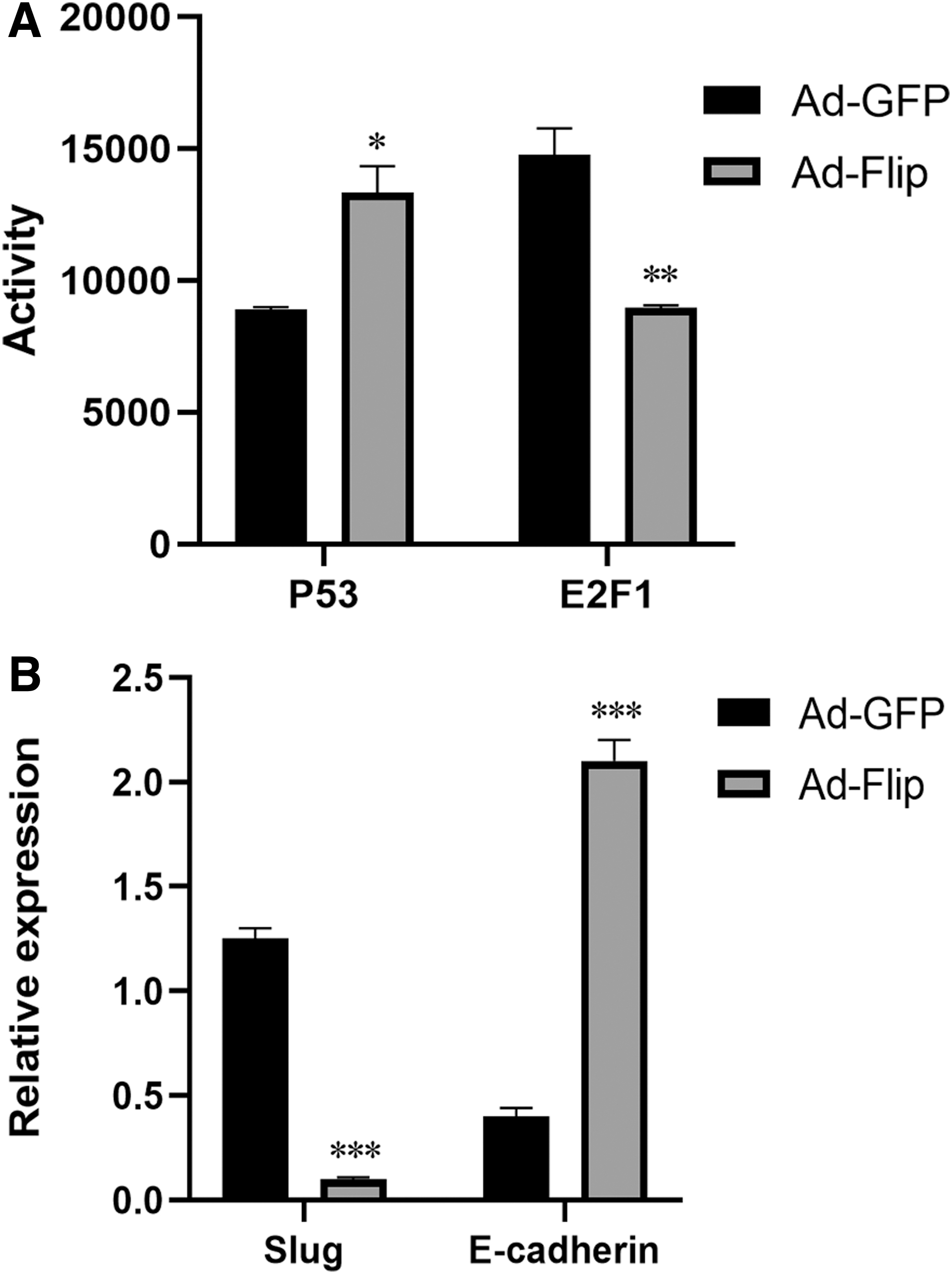
P53 and E2F1 activity and expression of the Slug and E-cadherin genes.
Bmp9-induced lipogenic and osteogenic differentiation of SV40T-SCs in vivo
Ad-BMP9-infected or Ad-GFP-infected SV40T-SCs were injected subcutaneously into nude mice. At 4 weeks, the mice were sacrificed and the samples were obtained. Pathological sections were stained with HE and Alcian blue staining. In the experimental group, SV40T-SCs were induced by BMP9 to differentiate into adipose tissue, including white and brown adipose tissue (Fig. 6A–C). A small bone matrix was observed, but osteogenesis is very weak relative to adipogenesis (Fig. 6D). In contrast, the cells were gradually absorbed and disappeared subcutaneously in the control group.

BMP9-induced lipogenic and osteogenic differentiation of SV40T-SCs in vivo.
Discussion
The addition of a small molecule substance named reversine to mouse myogenic lineage caused the cells to be dedifferentiated into mononuclear multipotent progenitor cells (C2C12 cells). These cells, in turn, can differentiate into adipocytes and osteoblasts under the action of adipogenic and osteogenic inducers (Chen et al., 2004). A more recent study on mouse fibroblasts screened four genes related to stem cell characteristics (Oct4, C-myc, Sox2, and Klf4). These genes proved to be enough to transform fibroblasts into embryonic stem-like cells, which have stem cells features, such as the expressing of specific embryonic stem cell genes and differentiating into three embryonic cell layers (Takahashi and Yamanaka, 2006).
In our study, we explored whether SV40T-SCs have differentiation capabilities. Using several differentiation-inducing conditions, we found that SV40T-SCs have the ability to differentiate into adipogenic and osteoblasts. The SV40T-SCs cultured in BMP9-cm showed fat granules in cytoplasm on third day of culture, and on the fifth day the cells differentiated into mature adipocytes. The cell ALP activity was significantly increased compared with control group. Alizarin red and Alcian blue staining was positive, indicating calcium deposition and bone collagen formation, respectively. The expression of adipogenic and osteogenic differentiation genes, such as c/EBPalpha, c/EBPbeta, OSX, and RUNX2, also increased significantly in SV40T-SCs cultured in BMP9-CM.
In vivo evaluation was performed by injection under the male nude mice of Ad-BMP9-infected SV40T-SCs and the results reinforced the data obtained by the in vitro experiments. BMP9 induced SV40T-SCs to differentiate into adipose tissue at sixth week, including white and brown adipose tissue. In addition, pathological sections stained by HE and Alizarin red showed a little bone matrix formation, although osteogenic differentiation is weak compared with adipogenic differentiation. Therefore, our experiments prove that SV40T-SCs have the ability to differentiate into adipose cells and osteoblasts.
Bone marrow osteoblasts and adipocytes are derived from MSCs, which can differentiate into mature adipocytes in two stages. Initially, MSCs differentiate into preadipocytes, which then differentiate into mature adipocytes under the action of a series of cytokines (Tang and Lane, 2012). The osteogenic and adipogenic differentiation of MSCs are regulated by a variety of cytokines and hormones, showing a balanced state of interrelationship and restriction. In osteoporosis, the osteogenic and adipogenesis differentiation of MSCs are unbalanced and excessive adipocytes are produced at the expense of osteoblast number, which ultimately leads to bone loss (Zhao, 2003). BMP9 can increase ALP activity, an early osteogenic differentiation indicator (Zhang et al., 2010). In addition, BMP9 can promote the expression of osteocalcin (OCN) and osteopontin (OPN), and promote the mineralization of extracellular matrix (Lin et al., 2016).
Flow cytometry showed that the SV40T expression knockout by Ad-Flip led to the blocking of the cell cycle in G1 phase and a decrease in the cell number in S phase. Thus, the SV40T can promote the passage of cells from G1 to S phase and accelerate the cell cycle. Luciferase reporter assays showed that SV40T can inhibit P53 activity and increase E2F1 activity. The upregulation of the p53 target gene mediates the G1 cell cycle arrest and the downregulation of E2F1 transcriptional activity has been associated with the inhibition of G1/S transition (Yang et al., 2011). Therefore, SV40T may regulate the cell cycle by interfering with the P53 and E2F1 activities.
SV40T antigen can bind to the ATPase domain of the wild-type p53 protein (wtp53) and to the C-terminus (351–708 region) of T antigen, forming a complex and inhibiting the binding of the p53 protein to the promoter. Thus, the SV40T prevents p53 from exercising its negative regulation function on cell growth, and induces increased DNA synthesis and cell transformation potential (Bocchetta et al., 2008). The SV40T contains a highly conserved LXCXG motif of the Rb retinoblastoma tumor suppressor (RB) family members, which can bind to the “pocket” structure of the pRb protein. The binding of pRB to E2F1 is a key step in the G1 phase of cell cycle, which can inhibit the E2F1 regulatory activity on the cell passage from G1 to S phase.
The combination of SV40T and pRB-E2F can separate E2F1 under the action of J-domain molecular chaperone. E2F1 promotes cells to enter S phase of the cell cycle (Harris et al., 1998). SV40 regulates the activity of cyclinD1 promoter by T antigen and increases cyclinD1 expression (Peterson et al., 1995; Watanabe et al., 1996). It can also upregulate cyclinD1 expression by Rb, p16, p21, cyclinD1, and CDK feedback regulation system. Other experiments have shown that excessive Cyclin D1 can also lead to decreased dependence on cell growth factor, leading to tumorigenesis (Peterson et al., 1995). These data are consistent with our findings.
Adipose and bone originate from the mesoderm (Kolios and Moodley, 2013; Lee et al., 1995). Our study found that SV40T-SCs have the ability to differentiate into osteoblasts and adipocytes. We revealed that SV40T-SCs have the ability to reverse differentiation. Therefore, we detected a process of transforming epithelial tissue to mesenchymal tissue. Endothelial-to-mesenchymal transition (EMT) refers to the process of transformation from epithelial cells into mesenchymal cells through specific biological behavior, which can modify the adhesion molecules expressed by the cells and reduce the cell adhesion ability (Gloushankova et al., 2018).
EMT was classified into three types, with the type I associated with embryonic development, type II with inflammation and wound healing, and type III with tumors (Tang et al., 2015). EMT occurs when multidirectional signaling factors induce the expression of transcription factors for specific EMT, including EMT-specific factors (EMT-TFs), epigenetic microRNAs, and post-translational regulatory factors, many of which are involved in embryonic development, damage repair, fibrosis, and tumor metastasis (Micalizzi and Ford, 2009).
SV40T can form a complex with p53 protein, resulting in inactivation of p53 protein. Wtp53 is a transcription inducer and target gene product of mouse diploid 2 (MDM2). Wtp53 upregulates MDM2 and forms a wtp53-MDM2-Slug complex, which promotes MDM2-mediated ubiquitination and Slug degradation. Furthermore, the E-cadherin expression was downregulated (Wang et al., 2009). Slug and E-cadherin are the key factors of EMT and mutation in p53 caused by SV40T prevented Slug degradation. The main molecular characteristics of EMT are the downregulation of E-cadherin, an epithelial marker, and the upregulation of nuclear transcription factors (Snail, Slug, and Twist) (Gloushankova et al., 2018). Snail, Slug, and Twist downregulate E-cadherin expression by binding to E-box of E-cadherin (Serrano-Gomez et al., 2016).
Thus, as SV40T leads to the upregulation of Slug, it results in the downregulation of E-cadherin expression, leading to EMT. This is the basis for SV40T-SCs to differentiate into fat and bone. We detected the Slug and E-cadherin expression by qPCR and the results showed that SV40T upregulates the Slug expression and downregulate the E-cadherin expression, proving the existence of EMT. In the differentiation inducing process of SV40T-SCs into osteoblasts and adipocytes by BMP9, first SV40T induces EMT and then BMP9 induces osteogenesis and adipogenesis.
SV40T effects on various cellular transformation are not consistent. The high SV40T expression in the transgenic mice eyes can induce the proliferation of nerve fiber cells and cone cells, and the formation of tumors (al-Ubaidi et al., 1992). SV40T-expressing cells were not always observed in pancreas, small intestine, prostate, and other organs of transgenic mice as cell proliferation, immortalization, or tumorigenesis (Furth, 1998). The neuroblasts in the midbrain and endocrine cells in the small intestine of the transgenic mice remained in their original state and did not proliferate (Asa et al., 1996). The reasons for these differing results are not yet clear at the moment. No tumorigenicity of SV40T-SCs was found in our in vivo experiments and, as described in this study, the high SV40T expression of is associated with cell cycle regulation, reverse differentiation, and EMT. In our study, different BMP9 systems were used to induce the differentiation of SV40T- SCs in vitro and in vivo, which is a limitation of the study.
In summary, SV40T regulates the cell cycle of SCs by interfering with P53 and E2F1 activity. The SV40T can upregulate the Slug expression and downregulate the E-cadherin expression to induce EMT. SV40T-SCs have the ability to differentiate into adipocytes and osteoblasts in vitro and in vivo, which is related to EMT.
Footnotes
Authors' Contributions
Z.l.-Z. conceived, designed, and supervised all aspects of the project with information from R.-F.L. R.-F.-L. performed most of the experiments. G.-X.N. performed, analyzed, and wrote the results of the cell culture experiments. D.W., C.G., and J.Y. participated in the writing of the article. R.-F.L. and Z.l.-Z. wrote the article with contributions from all authors. All authors read and approved the final article.
Author Disclosure Statement
The authors declare they have no conflicting interests.
Funding Information
This study is supported by the Health Commission of Hubei Province (WJ2017M150 and WJ2019M209).