
Abstract
Zygotic epigenetic reprogramming is the major initial event in embryo development to acquire a totipotent potential. However, the patterns of epigenetic modifications in bovine zygote were not well clarified, especially in the first cell cycle of bovine somatic cell nuclear transfer (SCNT) embryos. This study was conducted to examine the patterns of DNA methylation (5-methylcytosine [5mc] and 5-hydroxymethylcytosine [5hmc]) and histone H3 lysine 9 methylation (H3K9m2 and H3K9m3) in the first cell cycle of bovine in vitro fertilization (IVF) and SCNT embryos. In bovine zygotic development, the 5mc in the paternal pronucleus (pPN) undergoes partial demethylation from PN1 to PN3, and remethylation from PN4 to PN5, while 5hmc exhibits absolutely different patterns. The 5mc in SCNT embryos underwent much more dramatic demethylation and much earlier de novo methylation compared with their IVF counterparts, while 5hmc stayed stable from PN1 to PN4, and significantly increased at PN5, which made significantly higher level of 5mc and 5hmc at the end of the first cell cycle in SCNT embryos. Different H3K9m2 and H3K9m3 patterns were also observed between IVF and SCNT embryos. H3K9m2 and H3K9m3 asymmetrically distributed in parental genomes in IVF zygote, highly present in the maternal pronucleus, whereas faintly stained in the pPN. H3K9m2 and H3K9m3 in the somatic cell genome were gradually demethylated from PN1-PN4, and significantly increased at the end of the first cell cycle. TET3 dioxygenase was highly present in the first cell cycle of embryos compared with TET1 and TET2. Our results showed that SCNT embryos underwent aberrant epigenetic reprogramming in the first cell cycle; much more dramatic demethylation and significant higher remethylation were observed compared with IVF counterparts.
Introduction
Somatic cell nuclear transfer (SCNT) is a remarkable process in which the highly differentiated somatic nuclei can be converted into a totipotent state in the oocyte through a mechanism that depends on the reprogramming of epigenetic modifications. During the early development of SCNT embryos, epigenetic reprogramming of the injected somatic cell is a key event, with abnormal reprogramming of the epigenetic marks contributing to the inefficiency of somatic cell cloning (Dean et al., 2001; Santos et al., 2003; Tian et al., 2009).
DNA methylation is the most studied DNA epigenetic modification. During early mammalian development, reprogramming of genomic DNA methylation is observed both before and after the formation of the zygote, which was considered essential for establishing zygotic totipotency (Mayer et al., 2000; Reik et al., 2001; Santos et al., 2002). The DNA methylation pattern of zygotes was well clarified in mouse, and the 5-methylcytosine (5mC) content of the paternal chromosomes is actively reduced before the first cell division, while maternal pronuclei (mPN) maintain its 5mC status during the same period of time (Iqbal et al., 2011).
A good complementarity between 5mC and 5-hydroxymethylcytosine (5hmC) levels was observed at mouse zygote, the level of 5hmC increase in the paternal pronucleus (pPN) when 5mC decreases (Inoue and Zhang, 2011; Iqbal et al., 2011). The 5hmc is a newly discovered DNA methylation modification, which serves as an intermediate during DNA demethylation (Bhutani et al., 2011; Kriaucionis and Heintz, 2009). The epigenetic balance between 5mC and 5hmC plays an important role in epigenetic reprogramming of fertilized and SCNT embryos (Nie et al., 2017; Wossidlo et al., 2011; Zhang et al., 2016).
Although the reprogramming pattern of DNA methylation during mouse in vitro fertilization (IVF) and SCNT one-cell stage embryos was studied, the changes that occur during embryonic development are not strictly conserved between species (Beaujean et al., 2004). Therefore, it is important and necessary to investigate the reprogramming patterns of DNA methylation in bovine SCNT one-cell stage embryos to expand our understanding of the mechanism responsible for the abnormal development of bovine SCNT embryos.
Compared to mouse, partial demethylation of pPN is observed in bovine IVF zygote (Park et al., 2007). However, the dynamic patterns of 5mc were studied only in bovine fertilized zygotes; little is known about the dynamic patterns of 5mc in one-cell stage embryos derived from SCNT. And the studies on 5hmc patterns in bovine focus on preimplantation embryos (from two-cell stage to blastocysts). Thus, in this study, we aimed to gain further insight into the dynamic patterns of 5mC and 5hmC during the first cell cycle of bovine IVF and SCNT embryos.
Previous studies on mouse suggested a connection between DNA methylation and H3K9 methylation (Santos et al., 2005), it was reported that the presence of methylated H3K9 in the mPN protects the maternal genomic DNA from active demethylation, while the absence of H3K9 methylation in pPN allows active DNA demethylation to occur, The level of H3K9me2 in the chromatin might be critical in the DNA methylation reprogramming of zygote. Therefore, we also detect the levels of H3K9me2 and H3K9m3 in the first cell cycle of bovine SCNT embryos to further investigate the mechanism underlying the aberrant epigenetic reprogramming.
Recently, the conversion of 5mc to 5hmc in zygotes and implantation embryos was demonstrated to be catalyzed by mammalian Ten-Eleven Translocation (TET) proteins, including TET1, TET2, and TET3 (Gu et al., 2011; Huang et al., 2014; Ito et al., 2010; Shen et al., 2014; Tahiliani et al., 2009). The expression patterns of TET enzymes were also studied in IVF zygotes, implantation embryos, and SCNT implantation embryos (Cao et al., 2014; Lee et al., 2014; Nie et al., 2017; Page-Lariviere and Sirard, 2014; Wossidlo et al., 2011), but little is known about the expression patterns of TET enzymes in bovine SCNT one-cell stage embryos. In the first cell stage, embryos undergo dramatic DNA demethylation; therefore, we investigated the expression patterns of TET enzymes to evaluate the potential role of the TET enzymes in bovine SCNT one-cell stage embryos.
Materials and Methods
Materials
Unless otherwise noted, all chemicals were purchased from the Sigma Chemical Company (St. Louis, MO). Disposable, sterile plasticware was purchased from Nunclon (Roskilde, Denmark). All procedures were approved by the Animal Care and Use Committee of Northwest A & F University and Shaanxi University of Chinese Medicine, and performed in accordance with animal welfare and ethics.
Oocyte collection and in vitro maturation
Bovine ovaries were obtained from local abattoirs and transported to the laboratory (within 4 hours after death) in sterile 0.9% NaCl saline at 15°C to 20°C (Wang et al., 2011b) in a vacuum bottle. Cumulus oocyte complexes (COCs) were aspired from antral follicles (2 to 8 mm in diameter) using a 12-gauge disposable syringe. The COCs with evenly granulated cytoplasm and more than three layers of compact cumulus cells were selected and washed in maturation medium, comprising TCM-199 supplemented with 10% (v/v) fetal bovine serum (FBS), 1 μg/mL 17β-estradiol, and 0.075 IU/mL human menopausal gonadotropin (TCM-199 and FBS; Gibco, Grand Island, NY; HMG, Livzon, Zhuhai, Guangdong, China). Then, the COCs were transferred into maturation medium and incubated at 38.5°C in a humidified incubator of 5% CO2 in air for ∼20 hours.
In vitro fertilization
After maturation, the COCs were transferred into several microdrops of fertilization media (Fert-Talp medium supplemented with 5 mg/mL heparin sodium, 1.6 mg/mL hypotaurine, 0.3 mg/mL epinephrine, and 100 IU/mL penicillamine), and the frozen-thawed spermatozoa from a Holstein bull were used to inseminate the oocytes. Motile spermatozoa were selected by a swim-up technique and inseminated in a concentration of 1 × 106 mL−1. Oocytes and spermatozoa were incubated together for 24 hours. Presumptive zygotes were denuded by treatment with 0.1% bovine testicular hyaluronidase in phosphate-buffered saline (PBS), and then embryos were washed thrice in culture medium and cultured in modified synthetic oviductal fluid (mSOF) medium [supplemented with 8 mg/mL fatty acid-free of bovine serum albumin (BSA), 1% minimum essential medium (MEM) nonessential amino acid solution and 2% basal medium eagle (BME) essential amino acid solution] under mineral oil. All incubations were done at 38.5°C in a humidified incubator of 5% CO2 in air.
Somatic cell nuclear transfer
SCNT was performed essentially as previously described (Wang et al., 2011a). Briefly, matured oocytes were denuded of cumulus cells. Only oocytes with a first polar body were selected and used for SCNT. Enucleation was performed with a 20 μm (internal diameter) glass pipette by aspirating the first polar body and a small amount of surrounding cytoplasm. Enucleated oocytes were subsequently reconstructed by injecting a donor cell into the perivitelline space. The second to fifth passage of fibroblast cells from ear skin of a 1-week-old Holstein heifer were used as nuclei donors.
Oocyte-cell couplets were put in a droplet of fusion medium and vertically sandwiched between a pair of platinum microelectrodes connected to the micromanipulator. A double electrical pulse (35 V for 10 μs) was applied for oocyte-cell fusion. Successfully reconstructed embryos were kept in mSOF (containing 5 μg/mL cytochalasin B) for 2 hours until activation. All fused embryos were further activated in 5 μM ionomycin for 5 minutes, followed by exposure to 1.9 mM 6-Dimethylaminopurine in mSOF for 4 hours. Then embryos were washed thrice in culture medium and cultured in mSOF medium (supplemented with 8 mg/mL fatty acid-free of BSA, 1% MEM nonessential amino acid solution and 2% BME essential amino acid solution) under mineral oil. All incubations were done at 38.5°C in a humidified incubator of 5% CO2 in air.
Immunofluorescent staining of 5mC and 5hmC
One-cell stage embryos were classified into different stages (PN1 to PN5) according to size, position, and conformation of pronuclei. For each group, five embryos at PN1, PN2, PN3, PN4, or PN5 in the same batch were randomly chosen for immunofluorescent (IF) staining. Embryos were washed with PBS containing 0.2% (w/v) polyvinylpyrrolidone (PVP) for 5 minutes and fixed for 2 hours in 4% paraformaldehyde at room temperature (RT). Then embryos were washed thrice in PBS/PVP (2 minutes each), and permeabilized with 0.2% Triton X-100 in PBS for 30 minutes.
After permeabilization, embryos were incubated with RNaseA (50 μg/mL) for 1 hour at 37℃ in the dark, then washed with wash buffer for 5 minutes, and incubated at 37℃ for 30 minutes with 3M HCL/PVP to denature DNA. Then embryos were neutralized with 100 mM Tris-HCL for 15 minutes at RT, and washed for 5 minutes and blocked in 2% BSA in PBS overnight at 4℃.
After blocking, embryos were transferred to a solution of 1 μg/mL rabbit monoclonal to 5mC antibody (ab214727; Abcam, UK), 1 μg/mL rat monoclonal to 5hmC antibody (ab106918; Abcam), and 1 μg/mL mouse monoclonal to ssDNA antibody (MAB3868; Sigma-Aldrich, St. Louis, MO) diluted in blocking solution overnight at 4℃. Then embryos were washed thrice (5 minutes each) with PBS/PVP and transferred to a solution of 1 μg/mL 488 Alexa Flour labeled anti-rat IgG (ab150157; Abcam), 1 μg/mL 555 Alexa Flour labeled anti-rabbit IgG (Beyotime, NanTong, China), and 1 μg/mL 350 Alexa Flour labeled anti-mouse IgG (Beyotime) diluted in blocking solution and incubated for 2 hours in the dark at RT. After extensive washing thrice (5 minutes each), embryos were mounted with cover slips on microscope slides.
IF staining of H3K9m2 and H3K9m3
One-cell stage embryos were classified into different stages (PN1 to PN5) according to size, position, and conformation of pronuclei. For each group, five embryos at PN1, PN2, PN3, PN4, or PN5 in the same batch were randomly chosen for IF staining.
Embryos were washed in 0.2% PVA-PBS, fixed for 2 hours in 4% paraformaldehyde in PBS, and permeabilized with 0.2% Triton X-100 in PBS for 30 minutes at RT and then blocked in 2% BSA in PBS overnight at 4°C. After blocking, embryos were transferred to a solution of 1 μg/mL mouse monoclonal to histone H3 dimethyl K9 (ab1220; Abcam) and 1 μg/mL rabbit polyclonal to H3 trimethyl K9 (ab8898; Abcam) diluted in blocking solution overnight at 4°C. Then embryos were washed thrice for 5 minutes each in 0.2% PVA-PBS and transferred to a solution of 1 μg/mL 488 Alexa Flour labeled anti-rabbit IgG (Beyotime) and 1 μg/mL 555 Alexa Flour labeled anti-mouse IgG (Beyotime) diluted in blocking solution and incubated for 2 hours in the dark at RT. After extensive washing thrice (5 minutes each), embryos were transferred to DAPI solution for chromatin visualization. Then, embryos were mounted on slides to detect the fluorescence.
Quantitative real-time polymerase chain reaction (PCR)
For each group, 20 one-cell embryos at PN1, PN3, or PN5 in the same batch were randomly chosen and pooled for total RNA extraction. Total RNA was isolated using the Cells-to-Signal™ Kit (Invitrogen, Carlsbad, CA) according to the manufacturer's protocol. cDNA was produced and amplified using a cDNA synthesis kit (Takara, Dalian, China) according to the manufacturer's instructions, oligo dT and random primers were used together. Primers for all genes were designed cross-intron by primer 5.0 software (Premier Biosoft International, PaloAlto, CA), and were based on bovine RNA sequences found in Genbank (Table 1). The specificity of all primers was tested using a BLAST analysis against the genomic NCBI database.
Primer Sequences
All qRT-PCR were performed in triplicate in a 10 μL reaction volume on the quantitative real-time PCR StepOne plus system (ABI, Carlsbad, CA) using the SYBR Premix Ex TaqTM Ⅱ (Takara), containing 5 μL 2 × SYBR Green premix, 0.4 μL of forward and reverse primers (20 pmol/mL), respectively, 1 μL embryonic cDNA, 0.2 μL Rox, and 3 μL RNAase-free and DNAase-free water. The PCR was initiated with 30 seconds at 95°C to activate hot-start Taq polymerase, followed by 40 cycles of denaturation at 95°C for 5 seconds, and annealing and extension at 60°C for 30 seconds. During each cycle, fluorescence was measured, and a melting curve analysis was produced to confirm a single gene-specific peak and to detect primer/dimer formation.
A negative template control of diethylpyrocarbonate (DEPC) water was used to ensure that there was no nucleotide contamination. The relative amount of each target gene was determined using the 2−△△Ct method (Rutledge and Cote, 2003) and the amount of transcripts of target genes were normalized to transcripts of housekeeping gene H2a or 18sRNA.
Immunofluorescence intensity analysis
Immunofluorescence was examined by epifluorescence using a Nikon eclipse Ti-S microscope equipped with a Nikon DS-Ri1 digital camera (Nikon, Tokyo, Japan). Images were individually captured and transported to computer with Nikon NIS element software. All images were captured using the same settings and without any adjustment of constant or brightness.
Fluorescence was measured by analysis of the images with Image-Pro Plus 6.0 software (Media Cybernetics, Silver Spring). The average fluorescence intensity of the stained area of nucleus in the embryo represents the DNA methylation, hydroxymethylation, and H3K9m2 or H3K9m3 levels of the embryos before the two-cell stage. Images were first converted to grayscale and inverted. After correcting optical density (average cytoplasmic intensity was measured for normalization to background), the male and female pronuclei or pseudo-pronucleus were outlined, and integrated optical density (IOD) and area of those nuclei were measured; the average normalized fluorescence intensity for a single embryo was represented by “sum IOD/sum area.”
Statistical analysis
Experiments were repeated four times, each replicate of the experiments was carried out using oocytes matured on the same day to remove any batch effect of oocytes. Data of relative fluorescence intensity and gene expression were analyzed by one-way analysis of variance and Tukey's LSD test using SPSS 18.0 software. Differences were considered significant at p < 0.05.
Results
Dynamic appearance of 5mC and 5hmC during bovine IVF zygotic development
First, bovine IVF pronuclear embryos were analyzed for 5mC and 5hmC expressions at 2 to 5 hours, 6 to 9 hours, 10 to 13 hours, 14 to 17 hours, and 18 to 21 hours after fertilization, which represented five different developmental stages of pronuclear progression (PN1 to PN5). The zygotes were classified into different stages according to size, position, and conformation of pronuclei.
The global DNA methylation pattern of bovine IVF zygotes was analyzed throughout pronuclear development by IF staining for 5mC and 5hmC. All antibodies show a strong reactivity in advanced zygotes (Fig. 1). At the earliest developmental stages after insemination (PN1), both the mPN and the pPN showed similar intensity of 5mc and 5hmC. On further development, the fluorescence intensity of 5mC was significantly higher in the mPN at PN2, PN3, and PN4 than that in the pPN, whereas the fluorescence intensity of 5hmC was significantly lower in the mPN at PN3, PN4, and PN5 than that in the pPN.

5hmc and 5mc patterns in the first cell cycle of bovine IVF and SCNT embryos.
The dynamic pattern of the normalized 5mC fluorescence (5mC/DNA) of the mPN and the pPN during pronuclear development was similar. The 5mC signal significantly decreased from PN1 to PN4, and then moderately increased to PN5 at pPN. Similar, but much less pronounced changes were detected in the mPN. The dynamic pattern of 5hmc was different for the mPN and pPN. The 5hmc signal significantly increased from PN1 to PN3, and then slightly decreased to PN5 at pPN. At mPN, the 5hmc signal moderately increased from PN1 to PN2, and then decreased to PN5 with no significant alteration of 5hmc from PN3 to PN5 being observed.
Dynamic appearance of 5mC and 5hmC in SCNT pronuclear stage embryos
We next examined the dynamics of 5mC and 5hmC in chromosomes of bovine one-cell SCNT embryos at 2 to 5 hours, 6 to 9 hours, 10 to 13 hours, 14 to 17 hours, and 18 to 21 hours postactivation. The embryos were classified into five stages of pronuclear progression (PN1 to PN5) according to size, position, and conformation of pronuclei.
Cloned embryos only have one pronucleus derived from injected somatic cell. It is interesting to investigate the methylation and demethylation pattern of pronuclear stage cloned embryos, and the different methylation and demethylation patterns existed among IVF and cloned pronuleus. As shown in Figure 1B, the dynamic pattern of the normalized 5mC and 5hmC fluorescence (5mC/DNA and 5hmC/DNA) of the injected somatic cell during pronuclear development was different. The 5mC signal significantly decreased from PN1 to PN3, and then significantly increased to PN5. Conversely, the 5hmc levels stayed stable from PN1 to PN4, and then increased to a significantly higher extent at PN5.
Compared to IVF zygotes, the intensities of 5mC and 5hmC in SCNT one-cell embryos were significantly higher at the earliest developmental stage (PN1), and then 5mc underwent dramatic demethylation to a low level similar to pPN at PN3, while 5hmc stayed stable. At the end of the first cell cycle, the intensities of 5mc and 5hmc in SCNT embryos increased to significant higher levels compared to IVF counterparts.
Dynamic appearance of H3K9m2 and H3K9m3 in bovine IVF and SCNT pronuclear stage embryos
The presence of H3K9m2 and H3K9m3 during the development of bovine pronuclear embryos was also demonstrated by immunofluorescence. We examine H3K9m2 and H3K9m3 signals in IVF and SCNT embryos at 2 to 5 hours, 6 to 9 hours, 10 to 13 hours, 14 to 17 hours, and 18 to 21 hours postfertilization or activation. The zygotes were classified into five stages of pronuclear progression (PN1 to PN5) according to size, position, and conformation of pronuclei.
As shown in Figure 2, H3K9m3 signal asymmetrically distributed in parental genomes during zygotic development. H3K9m3 signal was highly present throughout pronuclear development in the mPN, whereas faintly stained in the pPN. Compared to IVF embryos, H3K9m3 signal exhibited a different pattern in the SCNT embryos. At the earliest developmental stages after activation (PN1), H3K9m3 mark was significantly higher than that in IVF embryos, both in mPN and pPN. Then H3K9m3 underwent dramatic demethylation to a low level (PN1 to PN2), and stayed stable from PN2 to PN4. At these stages (PN2 to PN4), H3K9m3 signal in SCNT embryos was significantly higher than that in pPN of IVF embryos, but significantly lower than that in mPN of IVF embryos. At the end of the first cell cycle, the intensities of H3K9m3 in SCNT embryos increased to significantly higher levels compared to IVF counterparts.
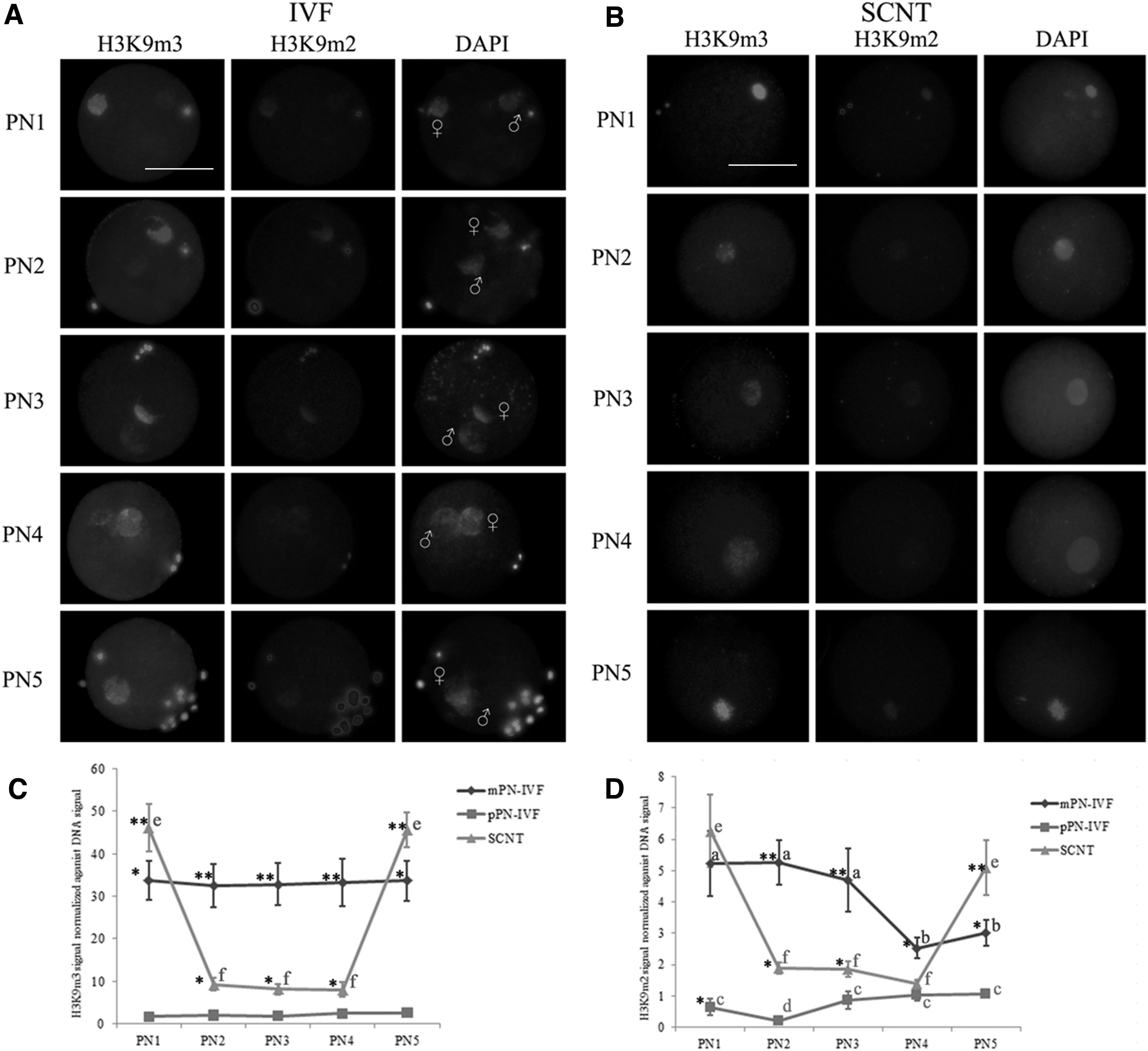
H3K9m3 and H3K9m2 patterns in the first cell cycle of bovine IVF and SCNT embryos.
Similar to the pattern of H3K9m3, H3K9m2 was also asymmetrically distributed in parental genomes of IVF embryos. H3K9m2 signal persisted at a low level at PN1, PN2, and PN3 in the mPN, and decreased to a lower level at PN4 and PN5, whereas an almost invisible H3K9m2 signal was detected in the pPN at PN1 to PN5, the lowest signal exhibited in PN2. H3K9m2 signal were significantly higher in mPN than that in pPN from PN1 to PN5.
In the SCNT embryos, H3K9m2 signal exhibited a different pattern from their IVF counterparts. At the earliest developmental stages after activation (PN1), H3K9m2 mark exhibited a similar level with mPN, which was significantly higher than that in pPN. Then H3K9m2 underwent dramatic demethylation to a low level (PN1 to PN2), and stayed stable from PN2 to PN4. At PN2 and PN3, H3K9m2 signal in SCNT embryos was significantly higher than that in pPN, while significantly lower than that in mPN. At the end of the first cell cycle, the intensities of H3K9m3 in SCNT embryos increased to significantly higher levels compared to IVF counterparts.
Expression levels of TET enzymes in bovine IVF and SCNT pronuclear stage embryos
The results (Fig. 3) showed that transcripts for TET1, TET2, and TET3 were differentially expressed in IVF and SCNT one-cell stage embryos. TET1 expression could barely be detected, and TET2 was only moderately expressed, while TET3 showed extremely high levels of expression. From PN1 to PN5, the expression of TET3 increased in both IVF and SCNT one-cell stage embryos. Compared to IVF zygotes, the expression of TET3 was significantly lower in SCNT embryos at PN1 and PN3 stage.
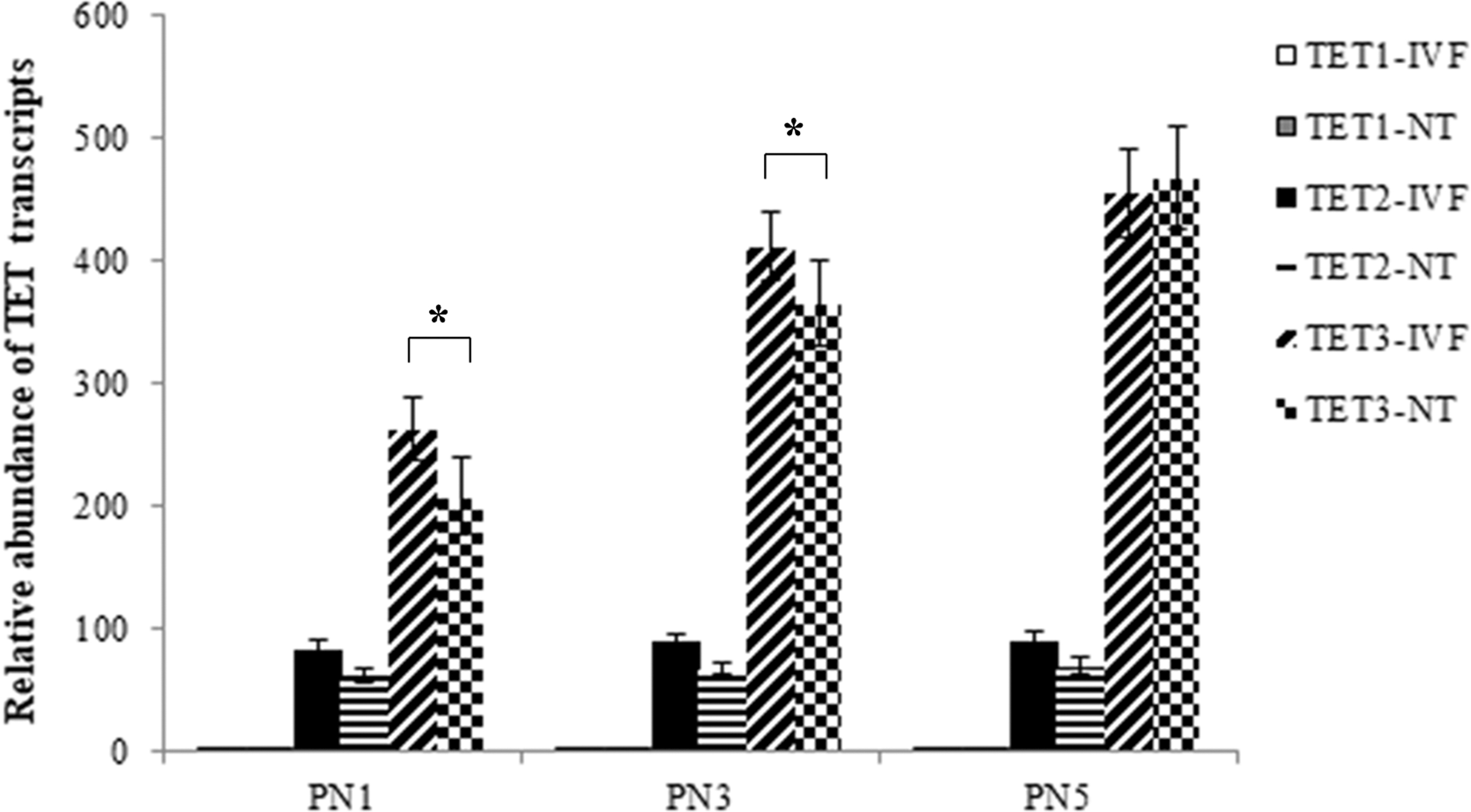
Relative abundance of TET1, TET2, and TET3 transcripts in PN1, PN3. and PN5 of the first cell cycle of bovine IVF and SCNT embryos. The values are mean ± SD. Asterisks show significant changes of TET3 expression between IVF and SCNT embryos (p < 0.05). TET, Ten-Eleven Translocation.
Discussion
DNA methylation reprogramming of one-cell stage embryos plays important roles in mammalian embryogenesis. A clear understanding of dynamic 5mc and 5hmc in one-cell stage SCNT embryos may yield more knowledge to clarify the mechanism of epigenetic reprogramming and improve somatic cloning efficiency. However, the DNA methylation reprogramming patterns in one-cell stage bovine SCNT embryos are not fully elucidated. Thus, in this study, we provided a complete description of 5hmC and 5mC reprogramming in first cell cycle of bovine IVF and SCNT embryos. We also analyzed the dynamics of H3K9m2 and H3K9m3 distribution, and the expression pattern of TET enzymes during the first cell stage in bovine IVF and SCNT embryos. To achieve this, we classified the zygotes into five PN stages (PN1 to PN5) according to size, position, and conformation of pronuclei.
Wossidlo et al. (Wossidlo et al., 2011) found that in mouse one-cell stage SCNT embryos, a strong demethylation of 5mc happened quickly after activation, and persisted into 12 hours postactivation (hpa), which was similar with the pattern of their IVF counterparts. In porcine one-cell stage SCNT embryos, a constant high level of 5mc signal was observed up to 16hpa, and then a demethylation of 5mc happened, which is similar with the pattern in pPN (Cao et al., 2014; Nie et al., 2017). In addition, a demethylation of the 5mc was also found in pronuclei of rabbit SCNT embryos at 4hpa, which is 1 to 2 hours faster compared with IVF zygotes (Lepikhov et al., 2008).
Our results on bovine were similar with the previous results in mouse and rabbit, but with some differences. At the early developmental stages (PN1 to PN3, up to 13hpa) of bovine SCNT one-cell stage embryos, a dramatic demethylation of 5mc was observed. However, after PN3, a rapid de novo methylation of 5mc happened and up to a higher level at PN5, which was not reported in other species. Compared to IVF zygotes, SCNT embryos undergo much more dramatic demethylation and much earlier de novo methylation. At the end of one-cell stage, the 5mc level of SCNT embryos was significantly higher than their IVF counterparts, and this hypermethylation pattern persists into the two-cell stage (Dean et al., 2001; Santos et al., 2003). It was well known that majority of cloned bovine embryos exhibit DNA hypermethylation at two-cell stage compared with IVF embryos (Dean et al., 2001; Santos et al., 2003; Zhang et al., 2016).
The study of one-cell stage SCNT embryos on mouse and porcine showed two different 5hmc patterns. In mouse, the 5hmC signal decreased during the first few hours of development and is followed by an increase reaching high levels at 12hpa (Wossidlo et al., 2011), while a constant high level of 5hmc was observed through the entire pronuclear stage in porcine, which was similar to the 5hmc pattern of male pronucleus in porcine IVF zygotes (Nie et al., 2017).
Unlike mouse and porcine, our study on bovine SCNT embryos showed that the 5hmc stayed stable from PN1 to PN4, and then increased to a significantly high extent at PN5, which indicated that the 5hmc patterns in the earliest stage of SCNT embryos were not strictly conserved between species. Compared to their IVF counterparts, the 5hmc pattern of SCNT embryos was absolutely different. The 5hmc level was higher in SCNT embryos at the earliest stage, and then stayed stable when IVF zygotes first increased and then decreased; at the end of the one-cell stage, SCNT embryos exhibited a significant increase in 5hmc, while IVF zygotes showed a significantly lower level. These results showed that aberrant reprogramming of 5mc and 5hmc existed at bovine one-cell stage SCNT embryos, and significantly higher levels of 5mc and 5hmc were observed at the end of the one-cell stage.
Our results of 5mc pattern in bovine IVF zygotes showed that the paternal DNA undergoes partial demethylation compared to mouse, and then followed by de novo methylation within a short period during the pronucleus stage. These results were similar with the previous observations in cattle (Park et al., 2007; Wossidlo et al., 2011). The global changes in 5hmc observed in this study showed similar pattern with the previous studies in mouse (Wossidlo et al., 2011).
Double fluorescent staining of 5mc and 5hmc in this study showed that the dynamic changes between 5mc and 5hmc were completely inversed in the pPN; an increase of 5hmc was accompanied with a decrease of 5mc. The changes in the mPN were much more moderate, and were not completely inversed. This inverse relationship between 5mC and 5hmC in pPN was also observed in mouse and rabbit (Lepikhov et al., 2008; Reis Silva et al., 2011; Salvaing et al., 2012; Wossidlo et al., 2011). However, this inverse relationship was not observed in horse (Heras et al., 2017), instead, both parental genomes displayed sustained and similar levels of methylation and hydroxymethylation in horse zygotes. This strong association between the patterns of 5mc and 5hmc was also not observed in bovine one-cell stage SCNT embryos.
We further studied the expression patterns of TET enzymes in the first cell cycle of bovine embryos. Our results on IVF zygotes showed that only TET2 and TET3 were detected with a significantly higher level of TET3. This expression pattern was similar to the previous reports in bovine (Page-Lariviere and Sirard, 2014) and mouse (Gu et al., 2011; Wossidlo et al., 2011). SCNT embryos showed similar dynamics of TET expression during the first cell stage. Therefore, we hypothesized that TET3 was predominantly responsible for the dynamic conversion of 5mc to 5hmC in bovine SCNT one-cell embryo, which was demonstrated in bovine and mouse zygotes earlier (Gu et al., 2011; Page-Lariviere and Sirard, 2014; Wossidlo et al., 2011). However, when compared with IVF zygotes, the levels of the TET3 were significantly lower at PN1 and PN3 of SCNT embryos. The abnormal 5mc and 5hmc levels in the first cell stage of SCNT embryos might be attributed to its abnormal TET3 expression to some extent.
H3K9 methylation plays an important role in early embryo development and epigenetic reprogramming (Ancelin et al., 2016; Torres-Padilla et al., 2007; Wang et al., 2018). In this study, H3K9m2 and H3K9m3 in the somatic cell genome were gradually demethylated in the SCNT embryos following activation, and significantly increased at the end of the first cell cycle. This pattern was different from their IVF counterparts; no remarkable remethylation of H3K9m2 and H3K9m3 was observed at the end of the first cell cycle in IVF zygotes. In mouse, different H3K9m2 and H3K9m3 patterns were also observed between IVF and SCNT embryos (Wang et al., 2007), a different chromatin structure between somatic cell genome and oocyte chromatin was thought to be the cause.
In mouse, H3K9m2 and H3K9m3 were asymmetrically distributed in parental genomes, the mPN exhibited H3K9m2 and H3K9m3, while the pPN never developed such methylation marks on H3K9 (Wang et al., 2007), Compared to mouse, H3K9m2 and H3K9m3 exhibited slight differences in bovine IVF zygotes; H3K9m2 and H3K9m3 were incomplete demethylated and slightly present in the pPN. Our results indicated that the asymmetry of H3K9me2 and H3K9m3 in zygotes was evolutionary conserved, and aberrant reprogramming of H3K9me2 and H3K9m3 modifications existed at the first cell cycle of bovine SCNT embryos.
In particular, it was demonstrated that PGC7 protects maternally methylated DNA from oxidation or demethylation by recognizing H3K9m2 (Nakamura et al., 2012); therefore, H3K9 methylation has been suggested to be mechanistically linked to DNA methylation. In mouse zygotes, H3K9m2 exhibits asymmetric distribution between the mPN and pPN (Santos et al., 2005), which is coincident with the distribution pattern of DNA methylation (Mayer et al., 2000). In this study, H3K9m2 and H3K9m3 also exhibit asymmetric distribution between the mPN and pPN. H3K9m2 and 5mc underwent partial demethylation and then de novo methylation, while the H3K9m3 stayed stable.
Our results indicated that association between the pattern of H3K9me2 and 5mC was much more stronger than that between H3K9m3 and 5mc. The incomplete demethylation of 5mc might be attributed to the partial demethylation of H3K9m2. Although the demethylation patterns of 5mc and H3K9m2 were slightly different on genome, a close correlation between DNA methylation and H3K9m2 appears to be conserved between different mammalian species.
The results of this study were useful in understanding the patterns of DNA methylation and H3K9 methylation of bovine IVF and SCNT one-cell stage embryos. It showed that bovine pPN underwent partial demethylation, while SCNT embryos underwent much more dramatic demethylation and much earlier de novo methylation, which made significantly higher DNA methylation level at the end of the first cell cycle in SCNT embryos. H3K9m2 and H3K9m3 were asymmetrically distributed in IVF zygotes, and aberrant reprogramming of H3K9me2 and H3K9m3 modifications existed at the first cell cycle of bovine SCNT embryos. The aberrant 5mc and 5hmc levels in the first cell stage of SCNT embryos might be attributed to its abnormal TET3 expression and H3K9m2 levels to some extent.
Footnotes
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This work was supported by the foundation for fostering the natural science foundation of Shaanxi University of Chinese Medicine (No. 2017PY02) and the foundation for discipline innovative team of Shaanxi University of Chinese Medicine (No. 2019-QN03).