
Abstract
Adenosine plays a significant role in neurotransmission process by controlling the blood pressure, while adenosine triphosphate (ATP) acts as a neuromodulator and neurotransmitter and by activation of P2 receptors, regulates the contractility of the heart. Adenosine signaling is essential in the process of regeneration by regulating proliferation, differentiation, and apoptosis of stem cells. In this review, we have selected neurological disorders (Alzheimer's disease, Parkinson's disease, amyotrophic lateral sclerosis, multiple sclerosis, and epilepsy) with clinical trials using antagonists and epigenetic tools targeting adenosine receptor as a therapeutic approach in the treatment of these disorders. Promising results have been reported from many clinical trials. It has been found that higher expression levels of A2A and P2X7 receptors in neurological disorders further complicate the disease condition. Therefore, modulations of these receptors by using antagonists of these receptors or SAM (S-adenosylmethionine) therapy as an epigenetic tool could be useful in reversing the complications of these disorders. Finally, we suggest that modulation of adenosine receptors in neurological disorders can increase the regenerative phase by increasing the rate of proliferation and differentiation in the damaged tissues.
Introduction
The term “purinergic neurotransmission” was first coined by Burnstock after the identification of the role of ATP in the transmission of nonadrenergic and noncholinergic inhibitory nerves in guinea-pig taenia coli (Burnstock, 1972). Initially, it was supposed that one neuron can only release a single transmitter, referred as Dale's principle. Later in 1976, this misconception was cleared and the concept of as cotransmission became evident for the involvement of both noradrenaline and ATP in sympathetic transmission (Burnstock, 1976, 2014).
In 1978, two different types of purinergic receptors were proposed based upon the pharmacological properties (molecular structure and second messenger system): (1) one type of these receptors was selective for adenosine (it was named as P1 receptor, having four subtypes, designated as A1, A2A, A2B, and A3 receptors) and (2) the second type was selective for ATP/ADP (adenosine diphosphate) (it was named as P2 receptor) (Burnstock, 2018). P1 subtypes of purinergic receptors are metabotropic G protein-coupled receptors, requiring extracellular adenosine for its activation. Among these P1 subtype receptors, A1 and A2A receptors show high affinity for adenosine, whereas A2B and A3 receptors show low affinity for adenosine.
Mode of action of A1 and A3 subtypes of adenosine receptors is by Gαi/o protein coupling, causing inhibition of adenylate cyclase, while A2A and A2B receptors perform their activity through Gαs protein coupling, resulting in the activation of adenylate cyclase (Carpenter and Lebon, 2017; Glukhova et al., 2017). P2 subtype of purinergic receptors is further classified into two different families, (1) P2X receptors, are ionotropic ligand-gated ion channel receptors (having seven subtypes known as, P2X1-7) and they are activated by ATP, and (2) P2Y receptors, are metabotropic G protein-coupled receptors (having eight subtypes known as P2Y1, 2, 4, 6, 11, 12, 13, and 14) and they are activated by ATP, ADP, UDP-glucose, uridine diphosphate (UDP), or uridine triphosphate (UTP) (Valera et al., 1994).
Short-term activation of purinergic receptors is implicated in events like secretion, chemoattraction, acute inflammation, neuromodulation, and neurotransmission (Burnstock, 2016), whereas long-term activation of purinergic receptors is implicated in many key processes like development, regeneration, aging, and cancer by regulation of cell differentiation, cell migration, cell proliferation, and cell death (Burnstock, 2013).
Adenosine is a vital intermediate metabolic nucleoside, which is distributed throughout the human body. Adenosine mediates the glutamate release, which is the main exciting neurotransmitter of the brain (Fredholm et al., 2011; Sebastiao and Ribeiro, 1996). The adenosine levels increase in many immunological conditions of the body like inflammation hypoxia,, ischemia, cerebral lesions, and excitotoxicity (Ribeiro et al., 2002). Adenosine A2A receptors stimulate vasodilatation, tissue protection, immunosuppression, and sleep promotion (Cerqueira, 2004; El Yacoubi et al., 2003). Adenosine A2A receptors in immune cells perform a key role in protecting normal tissues from injury by reducing the inflammation (Ohta and Sitkovsky, 2001).
Ninety-five percent of neurons present in the striatum are GABAergic Medium spiny neurons (MSNs) having two subtypes (1) striatonigral MSNs and (2) striatopallidal MSNs. Highest expression of adenosine A2A receptors is present on the dendritic spines of striatopallidal MSNs where they regulate dopamine secretion (Mori, 2014). Low levels of A2A receptors are also present in the cerebral cortex and the hippocampus where they regulate glutamate secretion (Peterfreund et al., 1996). The striatonigral MSNs express mGlu and D1R and regulate the direct signaling pathway affecting motor neurons, whereas striatopallidal MSNs express D2R, A2AR, and metabotropic glutamate receptor (mGluR) and are associated with indirect pathways of motor activation and inhibition (Schiffmann et al., 2007).
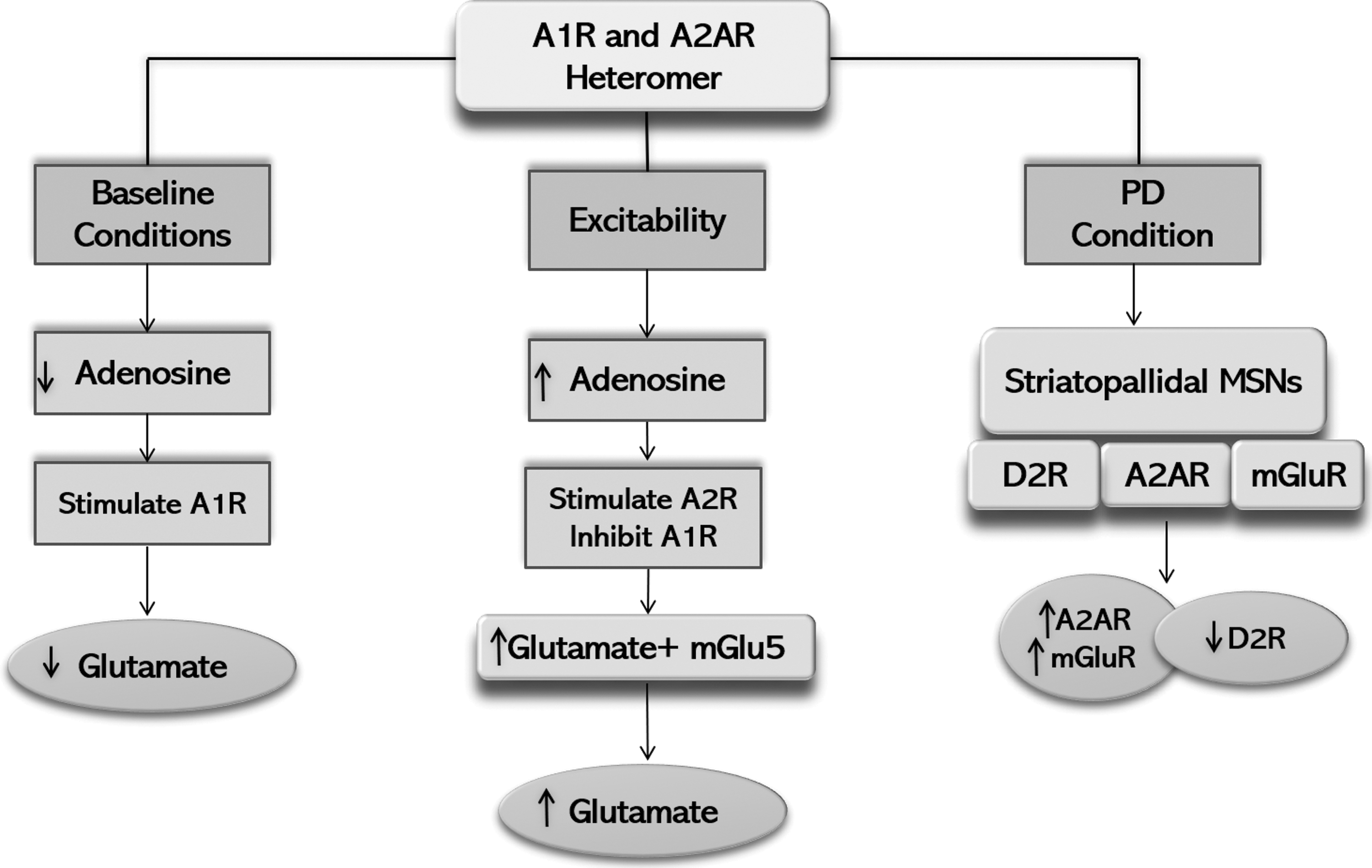
In Parkinson's disease (PD), when dopamine reserves are depleted, A2A receptors are expressed at the nerve ends of striatopallidal MSNs on Globus pallidus (GPe), which is the target site of A2A receptor antagonists (Mori, 2014) and transplantation of dopaminergic progenitor cells is derived from induced pluripotent stem cells (Doi et al., 2020). A2A receptors show their activity through physical interaction (oligomerization) with D2 receptors, and this type of receptor-receptor interaction produces a small adenosine/dopamine functional contact, which is regulated by A2A/D2 receptor oligomer. Two reciprocal interactions of antagonistic nature have been found between A2A/D2 receptors as shown in Figure 1.
Schematic diagram showing interaction of A2A receptor with other receptors.
The first interaction is an intermembrane interaction, where A2A receptor causes inhibition of D2 receptor, thus controlling the excitability of neurons and release of neurotransmitters. The second interaction takes place at the adenylate cyclase level in which D2 receptors inhibit A2A receptor-controlled gene expression and protein phosphorylation (Ciruela et al., 2011). Consequently, researchers are interested in developing A2A receptors antagonists as a principal therapeutic agent of nondopaminergic antiparkinsonian nature, due to the unique location of A2A receptors and A2AR/D2R oligomers in the brain (Fuxe et al., 2003). In addition, mGluRs are also co-localized with A2AR, D2R, and alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors at the dendritic spine of striatopallidal MSNs forming heteromeric multiplexes with A2A receptors (Doi et al., 2020; Rodrigues et al., 2005).
This type of co-localization offers a morphological platform for the presence of numerous interactions among mGlu/A2A/D2 receptors (Cabello et al., 2009). It is suggested that higher extracellular levels of adenosine and glutamate might stimulate A2A receptors and mGluR, which in turn may synergistically promote D2 receptors inhibition (Ferre et al., 2007). Extracellular levels of adenosine are elevated under situations of neuronal excitability, increasing A2A receptor activity and inhibiting A1 receptor activity. This provides a stimulus for glutamate release, which can further stimulate mGluR, and together with A2A receptors, mGluR causes further release of glutamate (Rodrigues et al., 2005).
Interestingly, PD patients presenting with dyskinesias show an upregulated expression of A2A receptors in the striatum. This increase in A2A receptor expression is because of Levodopa (L-dopa) treatment in these patients (Calon et al., 2004). A correlation has been found between the upregulation of A2A receptors in the striatum and the lymphocytes with the motor symptoms in patients with PD, who remained untreated or treated with a variety of medicines, but not limited to L-dopa alone (Varani et al., 2010). Upregulation of A2A receptors in patients with PD inhibits D2 receptors, and with low dopamine levels in striatum, it results in the dopaminergic neuron's death in substantia nigra, further leading to the synergistic loss of the D2R.
Modulation of Adenosine Receptors in Neurological Disorders
Alzheimer's disease
Alzheimer's disease (AD) is an advanced degenerative disorder of the brain affecting about 15 million people worldwide (Ferri et al., 2005). Clinical signs and symptoms of AD are memory loss, dementia, psychiatric disorders, progressive deterioration of cognitive functions, and inability to communicate, learn, and make decisions (Ferri et al., 2005; Waldau and Shetty, 2008). Major pathological features of AD are the intracellular neurofibrillary tangles and the development of extracellular senile plaques of amyloid-beta (Aβ) protein (Alzheimer et al., 1991). These plaques generally appeared years ago before the appearance of AD symptoms (Selkoe and Hardy, 2016).
Enough evidence is available about the crucial role of A1 and A2A receptors in the pathophysiology of AD as shown in Figure 2. Extremely low expression levels of A1 receptors have been found in the postmortem analysis of the hippocampus of AD patients (Ikeda et al., 1993), whereas upregulated expression levels of A1 receptors were found in the frontal cortex in both the initial and the advanced stages of AD patients (Albasanz et al., 2008). It has been shown that there is loss of A1 receptors in the hippocampus (CA1 region), which leads to cell death in this area (Rudolphi et al., 1989). Thus, the activation of A1 receptor induced tau phosphorylation and translocation of this receptor to the cytoskeleton of human neuroblastoma cells (SH-SY5Y cells), where this receptor is naturally expressed (Angulo et al., 2003) and might be associated with the progression of AD.

Treatment of AD can be enhanced beneficially with the help of adenosine receptor-based therapy causing regeneration of stem cells. Upregulated expression of adenosine A2A receptor is found in the cortical areas of patients with AD pathology, whereas blockade of A2A receptor can avoid cognitive defects and tau hyperphosphorylation. Postmortem analysis in and around Aβ plaques has shown an elevated expression of P2X7 receptors. Stimulation of P2Y2 receptor in AD may perform a neuroprotective role. Adenosine A1 receptors are differentially expressed in AD across various regions of the brain. AD, Alzheimer's disease.
The cell culture studies of A1 receptors from AD patients are demonstrated to form impaired Aβ fragments of neurofibrillary tangles. (Angulo et al., 2003), suggesting that activation of A1 receptors in AD could slow down the neurodegeneration process. Hence, further studies are required to precisely understand the role of A1 receptors in AD. Elevated expression levels of A2A receptors are found in the cortical regions of AD patients (Angulo et al., 2003) and in cortical tissues of animal models (Arendash et al., 2006). It has been shown that the blockade of A2A receptor activation in AD can prevent the cognitive impairment and memory loss. In a study of mice models, an oral dose of a selective antagonist of A2A receptor decreased tau hyperphosphorylation and improved spatial memory in AD (Arosio et al., 2016).
In another study of mice model, it has been reported that the memory loss induced by administration of Aβ by intracerebroventricular injection was prevented by the administration of either antagonist of A2A receptor or caffeine (Dall'Igna et al., 2007). Also, in A2A receptor (knockout) KO mice, the use of Aβ did not induce synaptotoxicity or produce any learning defect (Arendash et al., 2006). Therefore, it is concluded that the reduced activity of A2A receptor is linked with improved cognitive function in AD in vivo.
Parkinson's disease
PD is defined as a neurodegenerative disease of the substantia nigra pars compacta region of the brain, which is characterized by the extensive loss of dopaminergic neurons present in this area (Braak et al., 1995). Other parts of the brain that are also affected by this disease are nucleus basalis of Meynert, dorsal nuclei of the vagus, locus coeruleus, raphe nuclei, and catecholamine-secreting neurons in the ventrotegmental area of the brain stem (Damier et al., 1999). Microglial cells play a vital role in supporting homeostasis in the Central Nervous System (CNS), especially in the synapse. Noxious stimuli triggered with chemokine such as α-synuclein, β-amyloids, purines (ATP/ADP), and nitric oxide (NO) lead to microglial activation, which further leads to phagocytosis to keep synaptic homeostasis.
These neuroinflammations are well-known pathogenic processes involved in the early stages of PD, which is the target site for neuroprotective strategies with A2A receptor antagonists (Illes et al., 2020; Le et al., 2016). As a result of these neuroinflammations, elevated levels of interleukin (IL)-2, tumor necrosis factor alpha (TNF-α), IL-1β, and IL-9 are noted in the striatum, and elevated levels of TNF-α are also noted in the substantia nigra, which were confirmed in postmortem samples of PD patients (Mogi et al., 1996). Furthermore, elevated plasma levels of IL-6 were found to be correlated with the severity of PD (Pereira et al., 2016).
Among the ATP signaling molecules, the human adenosine A2A receptor comes across as a receptor with ample evidence of being involved in PD as shown in Figure 3. It performs neuroexcitatory role (Latini and Pedata, 2001) in the striatopallidal MSNs, a prominent site of its expression, by encouraging the release of glutamate and by regulating dopaminergic neurotransmission (Ferre et al., 1991). A2A receptor forms a functional complex with D2 receptor in the striatopallidal MSNs and antagonizes its functions reciprocally and independently at the second messenger level.

Death of dopaminergic neurons in PD results in the release of ATP in extensive amounts. ATP can induce continued stimulation of the P2X7 receptor that can create a large pore in the plasmatic membrane, which results in disturbance of ion balance leading to cell death, further deteriorating the pathology of PD. Expression of A2A receptor is elevated in initial stages of PD, which enhances interests in the use of A2A receptor antagonist as a valued treatment strategy. iPSC. iPSCs, induced pluripotent stem cells; PD, Parkinson's disease.
Evidence of this interaction has been observed in rat striatal membrane and human neuroblastoma cell lines. A2A receptor activation in the rat striatal membrane decreased D2 receptor affinity for its agonists with subsequent reduction of G protein coupling with D2 receptors. Similarly, in human neuroblastoma cell line, the activation of D2 receptors suppressed cyclic Adenosine Monophosphate (cAMP) production by the stimulation of A2A receptors (Mori, 2014). High expression of A2A receptor is found in the early phases of PD, which increases the severity of the disease (Villar-Menéndez et al., 2014).
The involvement of A2A receptors in PD has been proved in A2A receptor KO PD mice model, which provides protection against dopaminergic neuron degeneration induced by a mutant transgene of human α-synuclein (hm2-α syn) possessing both A53T and A30P (Kachroo and Schwarzschild, 2012). It is further proved in MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) mice model with the use of A2A receptor antagonist, which reestablishes the capability of microglia to respond to damage and provided neuroprotection most likely by modulating the microglial extension process in response to cell death (Gyoneva et al., 2014).
Amyotrophic lateral sclerosis
Amyotrophic lateral sclerosis (ALS) is a disease of the motor neuron, which leads to the death of the upper and lower motor neurons. Upper motor neurons extend from the brainstem and spinal cord to the cortex and lower motor neurons extend from the spinal cord or brainstem to the muscles (Kirk et al., 2019; Zucchi et al., 2019). ALS patients present with spasticity and atrophy of voluntary muscles, which are due to the loss of motor neurons (Zucchi et al., 2019). At final stages of the ALS, patients present with respiratory insufficiency, which may lead to the death of the patient due to respiratory failure (Hardiman et al., 2017). In addition to the motor symptoms, ALS patients also experience comorbidities of psychiatric and cognitive disorders such as bipolar disorders and schizophrenia (SZ) (Zucchi et al., 2019).
ALS is an aggressive disease, and the patients having ALS usually expires within 2–5 years after the diagnosis (Mitchell and Borasio, 2007). Currently, the only drug of choice against ALS, which has been approved by FDA (Food and Drug Administration), is Riluzole. This drug has been shown to increase the survival of patients by 2–3 months. Riluzole is much more effective if the treatment is started at the initial phases of the disease (Lee et al., 2013; Zoccolella et al., 2007a). Adenosine receptors play a significant role in the pathophysiology of ALS as shown in Figure 4. High adenosine levels are found in the cerebrospinal fluid (CSF) sample of ALS patients (Yoshida et al., 1999).
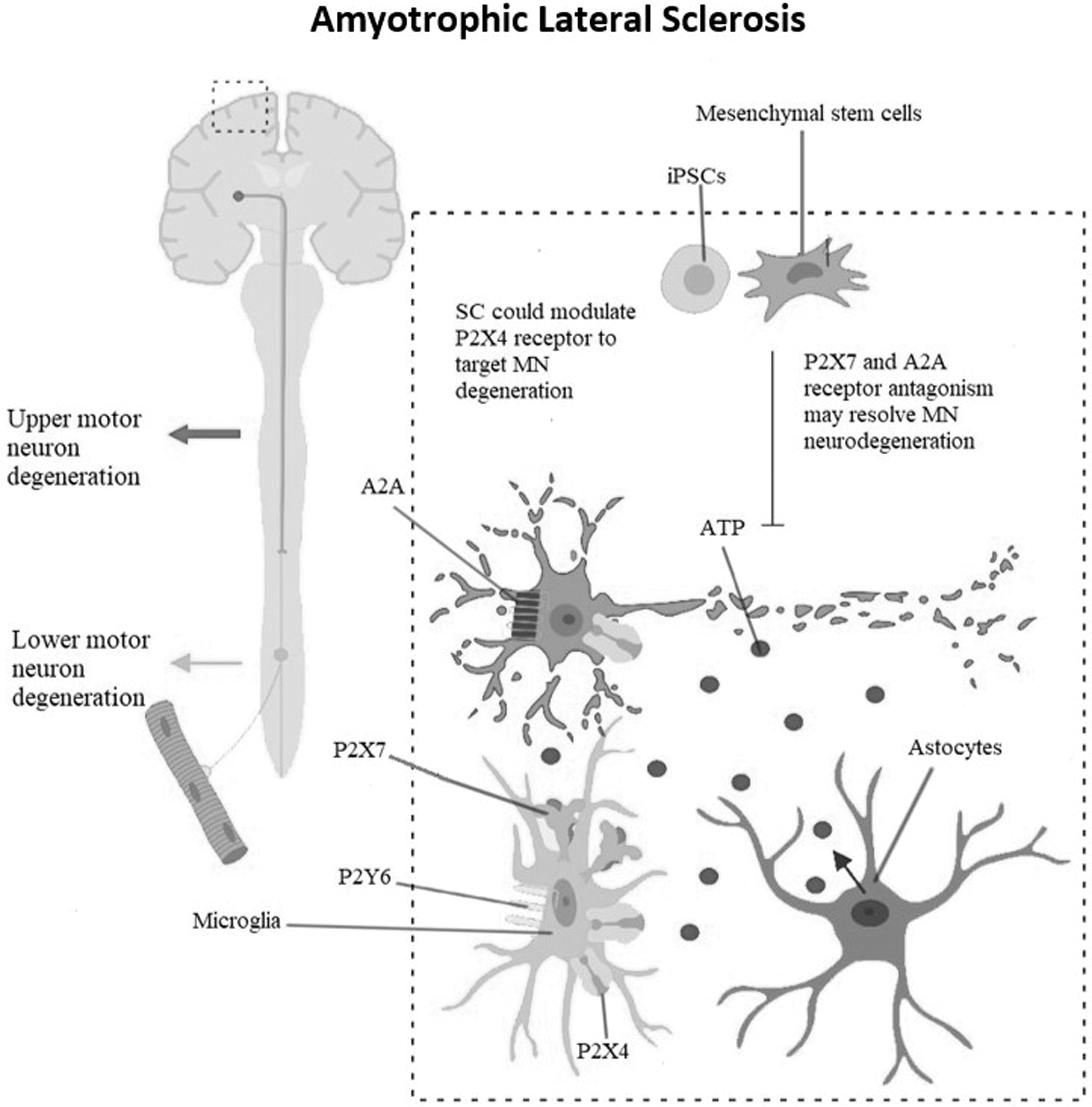
Upregulated expression of P2X4, P2X7, and P2Y6 receptor is reported in the pathophysiology of ALS. In the rat model of degenerating motor neurons (SOD1G93A), P2X4 receptor antibody precisely identifies the degenerating motor neurons. Elevated expression levels of adenosine are found in CSF of patients with ALS. The spinal cord of SOD1G93A mice and postmortem analysis of human samples also show an elevated expression level of adenosine A2A receptor, which attracts antagonists of A2A receptor for mediating a neuroprotective role in ALS. iPSC. ALS, amyotrophic lateral sclerosis; CSF, cerebrospinal fluid.
The previous researches have shown upregulated expression of adenosine A2A receptor in lymphocytes of patients with ALS (Vincenzi et al., 2013). In a preclinical trial performed on spinal cord cell culture, the administration of A2A receptor antagonists provided neuroprotection in motor neurons probably by reducing the activation of TrkB receptor (Mojsilovic-Petrovic et al., 2006). This neuroprotection provided by adenosine A2A receptor antagonists can be elucidated by the use of brain-derived neurotrophic factors in the pathology of ALS (Küst et al., 2002; Mojsilovic-Petrovic et al., 2006). Ng et al. (2015) in their study have shown an increased expression of A2A receptor in the spinal cord of SOD1G93A mice and the higher expression of A2A receptor was also found in the postmortem sample of ALS patients in the last stage of the disease.
The in vitro studies have also demonstrated the death of Embryonic stem cells (ESC)-derived motor neurons by adenosine therapy. To further validate these findings, partial deletion of (A2AR+/−) gene and the use of antagonists of A2A receptor in the SOD1G93A mice have prevented the death of ESC-derived motor neuron and decelerated the progression of the disease (Ng et al., 2015). In addition, A2A receptor agonist treatment in SOD1G93A mice at the presymptomatic stage resulted in the enhancement of neuromuscular transmission and had no effect at the symptomatic phase (Nascimento et al., 2014).
In disagreement with the previous statement, chronic intake of caffeine has resulted in reduced survival of SOD1G93A mice, impaired function of motor neurons, and has encouraged downregulation of A2A receptor in the spinal cord of wild-type mice (Potenza et al., 2013). These results may have been owing to the nonselective nature of caffeine as an A2A receptor antagonist. Importantly, these conflicting findings should be further tested.
Multiple sclerosis
Multiple sclerosis (MS) is a devastating disorder of the brain and spinal cord affecting around 2.5 million individuals globally, with more prevalence in women compared to men. The precise etiology of MS is still to be discovered; however, it is linked with genetic (Sawcer et al., 2011) and environmental factors, seropositivity of immunoglobulin G against infectious mononucleosis, and nuclear antigen of Epstein-Barr virus (Belbasis et al., 2015).
The signs and symptoms of MS typically appear at the most productive age group (between 20 and 40 years of age) (Simone et al., 2000), which highlights the high level of social and economic burden caused by this disease (Kobelt et al., 2017). Currently, there is no treatment available to cure the MS; the only option available is the symptomatic relief of the signs and symptoms (Costa et al., 2018). Therefore, a deep understanding of the role of adenosine receptor modulation in MS can provide a way toward successful therapeutic approach.
The past studies have indicated that the stimulation of A1 receptors plays a beneficial role in MS by reducing the levels of proinflammatory cytokine such as TNF-α. TNF-α induces demyelination and is believed to be an initiating factor in the development of MS pathology (Probert et al., 1995). Postmortem analysis of glial cells in MS patients demonstrated lower expression of A1 receptors in these cells (Johnston et al., 2001). Further confirmation has been achieved in A1 receptor KO mice by EAE (experimental autoimmune encephalomyelitis) induction, which leads to an aggressive relapsing type of MS with axonal loss, microglial activation, and demyelination (Tsutsui et al., 2004).
Chronic administration of A1 receptor antagonist (caffeine) results in the upregulated expression of A1 receptor in microglia, together with a reduction in the severity of the EAE, which may be further reduced by adjunct therapy of A1A receptor agonist (adenosine amine congener) (Tsutsui et al., 2004). Hence, caffeine treatment in the EAE animal model and MS patients has been established to play a neuroprotective and anti-inflammatory role (Hedström et al., 2016; Olsson et al., 2017). So stimulation of A1 receptors in MS patients is suggested to induce neuroprotection against neurological disorders as shown in Figure 5.

MS pathology shows an elevated expression of P2X7 receptor in oligodendrocytes, microglia, and astrocytes. Increased cell death in MS causes a massive release of ATP, which causes activation of the P2X7 receptor in microglia. Postmortem analysis of the highly demyelinated regions of the brain has revealed that expression of the P2Y12 receptor is reduced in these regions, and stimulation of the P2Y12 receptor in microglia is proposed to provide neuroprotection in this area. Furthermore, postmortem studies of the glial cells of MS patients have also found reduced expression levels of adenosine A1 receptors in these areas. MS, multiple sclerosis.
Epilepsy
Epilepsy is a neurological disorder of the central nervous system, which presents with at least two seizures of incontinence, falling, tongue biting, or concomitant epileptic irregularities in the electroencephalogram (de Timary et al., 2002). Previous studies have highlighted the role of adenosine receptors in modulation of epileptogenesis and presynaptic excitability as shown in Figure 6 (Malva et al., 2003). Variable expressions of A1 receptors have been noted in different parts of the brain in epilepsy patients. Upregulated expression of A1 receptors has been found in the neocortex region of the postmortem brains of epilepsy patients (Angelatou et al., 1993), and downregulated expression has been found in the hippocampus and temporal cortex regions (Glass et al., 1996).

Decreased ATPase activity is observed in hippocampal slices of epilepsy patients. Elevated expression of P2X7 receptor is found in the hippocampi of pilocarpine-induced chronic epileptic mice. Stimulation of adenosine A1 receptor has been found to encourage antiepileptic and anticonvulsant effects, while stimulation of A2A receptor seems to possess proconvulsant effects.
Similar downregulation has also been found in the nucleus reticularis thalami region of rat brain (Ekonomou et al., 1998). It has been observed in chronic epileptic rats that adenosine delivery to the hippocampus can decrease the frequency of spontaneous seizures (Gouder et al., 2003), and activation of A1 receptor in pharmacoresistant epileptic mice model can prevent spontaneous seizures (Boison, 2012; Sørensen and Kokaia, 2013). Lately, it has been found that acute administration of caffeine (nonselective antagonist of P1 receptor), at a dosage of 5 mg/kg, reduces seizure threshold significantly (Esmaili and Heydari, 2018).
Previous researches have shown that different dosages of caffeine produced different results through different pathways (Esmaili and Heydari, 2018) and high caffeine doses most likely mediated their effects through the NO pathway (Esmaili and Heydari, 2018). The ability of adenosine in suppressing the seizure is due to the inhibition of A1 receptors, which further results in the inhibition of glutamatergic transmission (Miranda et al., 2014).
Promising results have been achieved by using the antiepileptic properties of adenosine, primarily through its interaction with A1 receptor, and thus provided an alternative treatment approach against epilepsy by designing cells with knockdown expression of adenosine kinase (ADK). These implants can provide large quantities of adenosine, restricting its action to the foci of the seizure (Cavaliere et al., 2015). In this respect, the adenosine releasing brain implants have shown suppression in epileptogenesis (Li et al., 2007) and seizures (Huber et al., 2001).
Similarly, adenosine-releasing implants in a rat model with temporal lobe epilepsy were effective in seizure suppression, indicating that central activation of adenosine A1 receptors results in the reduction of seizure activity in a drug-resistant epileptic mouse model (Gouder et al., 2003). It can be concluded from the available information that the local delivery of adenosine into the brain is an effective way of treating intractable seizures (Gouder et al., 2003). An interesting study used different approaches for the modulation of ADK expression, and it was found that genetic downregulation of ADK enzyme resulted in the suppression of spontaneous recurrence of seizures, while overexpression of ADK enzyme resulted in the recurrence of seizures in mice (Theofilas et al., 2011).
Clinical Trials with Modulation of A2A Receptor Expression by Its Antagonists
The most efficient treatment of choice for the symptomatic relief of PD is L-dopa; however, its prolonged use can lead to motor dysfunctions (dyskinesias) (Obeso et al., 2000). To overcome this problem, the best alternative is nondopaminergic therapy by regulating the dopaminergic neurotransmission. A2A receptors have evolved as a prospective pharmacological target in PD due to its evident association with the dopaminergic neurotransmission system (Ferre et al., 2002). Since A2A receptors are exclusively present in the striatopallidal MSNs, their antagonists can specifically target a definite brain area (Brooks et al., 2008).
After the promising results of some preclinical studies showing the mechanism of action of A2A receptor in causing the inhibition in dopamine release in the badly affected regions of brain in PD, numerous clinical trials have been done in PD patients to evaluate the therapeutic potential of A2A antagonists. Preladenant, a selective and potential antagonist of A2A receptor, was testified for its efficacy in phase IIb and III of clinical trials. Results of the phase IIb clinical trial showed promising results with a reduction in the appearance of symptoms and improvement in the motor functions (Hauser et al., 2011); however, no efficacy of this drug was seen in the phase III results of clinical trial both as a monotherapy (Stocchi et al., 2017) and as an adjunctive therapy along with levodopa (Hauser et al., 2015).
Another antagonist of A2A receptor showing promising results is istradefylline (KW-6002) as shown in Figure 7. Istradefylline has proven its efficacy and safety as an adjunct therapy with levodopa or other anti PD therapies (Vorovenci and Antonini, 2015). A striatopallidal MSN in PD exerts selective loss of dendritic spine with A2A receptor localizing at spines, so istradefylline (KW-6002) targets the axon terminal of the MSN at GPe inducing motor activation through D2 receptors (Mori, 2014; Orru et al., 2011). Istradefylline use as a mono therapy in PD patients has not been proven valuable (Fernandez et al., 2010), but its use as a co-adjuvant may permit a lower dose of L-dopa. Striatopallidal MSNs or striatal GABAergic neurons contain heteromers that comprise mGluR, D2 receptor, and A2A receptors (Cabello et al., 2009).
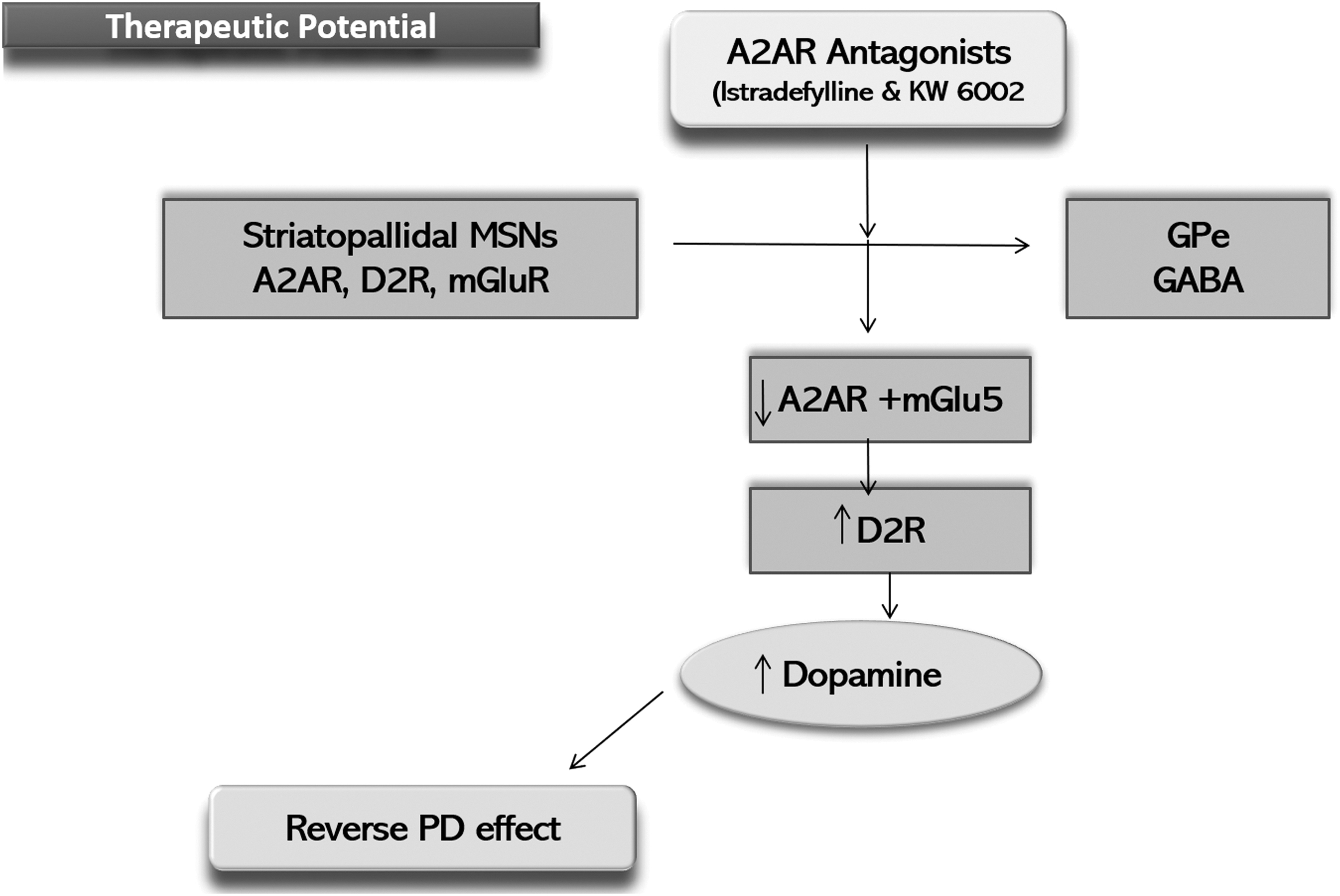
Schematic diagram showing therapeutic potential of A2A receptor antagonists.
Elevations in the extracellular levels of glutamate and adenosine can activate mGluR and A2A receptors, which are synergistically involved in the blockage of D2 receptors (Ferre et al., 2007). In a PD model rat, simultaneous inhibition of mGluR and A2A receptors has been proven to reverse the PD defects (Coccurello et al., 2004), thus making a strong base to propose antagonists of A2AR and mGluR are important therapeutic candidates for PD (Bonsi et al., 2007). Many clinical trials are in phase II and III to access the efficacy of these antagonists as adjunctive therapy with carbidopa and levodopa. A 12-week phase II clinical trial has proven the effectiveness of istradefylline in reducing the “off” state, without increasing the “on” state, and it was well tolerated as an adjunct therapy along with levodopa (Stacy et al., 2008).
Other clinical trials are in phase II to analyze the suitable concentration of istradefylline at 20 or 40 mg/day, both as a monotherapy and as an adjunct therapy in both early and advanced PD. Furthermore, phase III clinical trials have been done to assess the effectiveness, tolerability, and safety of istradefylline as an adjunct therapy at concentration of 10, 20, and 40 mg/day in moderate to severe PD patients. In fact, Istradefylline is approved in Japan as adjunctive therapy with levodopa, showed its ability as an anti-parkinsonian therapy (Zhu et al., 2014). Many other studies are in progress as shown in Table 1 using A2A receptor antagonists like V81444, to evaluate the potential modulatory role of these antagonists and their safety concerns. Some of these clinical trials were discontinued, specifically those that were using preladenant and vipadenant as A2A antagonist (Antonini and Poewe, 2014; Pinna, 2014).
Summarizing the Results of Clinical Trials Using Adenosine Receptor Modulation as a Therapeutic Approach Against Parkinson's Disease
Epigenetic Modulation of A2A Receptor as a Therapy Against Brain Disorders
DNA methylation
DNA methylation is as a natural process that takes place during the development of mammalian embryos and is also associated with proviral genes, endogenous transposon repression, and the inactivation of X-chromosome. DNA methylation is a major tool of epigenetics in mammals and it plays an important role in gene silencing by methylation of the cytosine preceding guanines at the well-established CpG sites. These CpG-rich areas in the genome are known as CpG islands (CGIs) ranging in size from 200 bp to several kilobases. CGIs are generally not methylated in the normal tissues, but they are methylated in tumor cells, particularly in the tumor repressor genes (Illingworth and Bird, 2009).
Methylation of the CpG sites is carried out by the enzymes of the transferase family referred to as DNA methyltransferases (Dnmt). These enzymes transfer a methyl group from methyl donors such as SAM to CpG sites in the DNA. Methylome map of human DNA has annotated the methylated genes in normal tissue as well as in human disorders like SZ and AD (Ballestar and Esteller, 2008). DNA methylation at the CpG sites hinders with the expression of gene in two ways. First, it interferes with the binding of transcription factors with the DNA by their methyl group causing hindrance in transcription. Second, it causes interference in gene expression by binding of specific methyl CpG-binding proteins (MBDs), like MBD1, MBD2, and MeCP2, to the methyl CpG sites.
These proteins recruit chromatin-remodeling complexes and histone-modifying complexes toward the methylated sites (Portela and Esteller, 2010). The significance of MBD proteins in gene expression is evident from the Rett syndrome, which is a complex mental disease resulting from the mutations in the MeCP2 gene (Amir et al., 1999). Examination of 12 loci, which are linked to AD, shows an age-specific epigenetic drift on the extent of DNA methylation (Siegmund et al., 2007; Wang et al., 2008). Furthermore, the extent of DNA methylation and the expression of DNA methyltransferase 1 (Dnmt1) are variable in different parts of the brain (Ladd-Acosta et al., 2007). It is noteworthy that Dnmt1 levels are higher in cortical interneurons, whereas the GAD67 gene is inhibited in SZ patients (Veldic et al., 2005).
Demethylating agents are a well-known epigenetic tool and they are in use as a therapy of cancer (Herranz and Esteller, 2006), but their utility as epigenetic therapy for the treatment of neurodegenerative disorders is poorly understood so far. The current focus in on SAM, which is a methyl donor required for DNA methylation and is found to be deficient in PD patients (Linnebank et al., 2010). Therefore, SAM can be a potential epigenetic therapy for AD treatment as in cell culture models, SAM already proved to downregulate PSEN1 and decrease β-amyloid production (Scarpa et al., 2003).
Endogenous SAM biosynthesis cycle
SAM is as a major methyl donor of the methionine in the biological systems causing methylations of proteins, DNA, lipids, and polyamines. SAM also serves as a glutathione precursor in the liver and possibly in the brain as well (Vitvitsky et al., 2006). Demethylation of SAM produces S-adenosylhomocysteine (SAH), which is further converted to homocysteine (HCY) and adenosine. To avoid HCY accumulation, it is remethylated and is converted to methionine (Chiang et al., 1996; Lu, 2000). Ratio of SAM/SAH is known as methylation potential, and it is extremely important to maintain this ratio endogenously. Chronic intake of L-dopa by PD patients produces hyperhomocysteinemia in the brain, peripheral tissues, and plasma.
This is because the metabolism of L-dopa needs SAM as a methyl donor (Muller and Kuhn, 2009; Zoccolella et al., 2010) and it has been proven in mice that L-dopa treatment depletes SAM content of the brain (Liu et al., 2000). Elevated levels of plasma HCY have been linked with impairment of cognitive and motor neurons in PD patients and with the pathogenesis of other neurological disorders as shown in Figure 8 (Morris, 2003; Quadri et al., 2004). Methylenetetrahydrofolate reductase (MTHFR) is an enzyme that converts HCY to methionine.

Schematic diagram showing consequences of hyperhomocysteinemia.
Polymorphism has been reported in MTHFR gene, which is linked with its decreased enzymatic activity, causing a rise in the levels of HCY in PD patients taking L-dopa treatment as shown in Figure 8 (Yasui et al., 2000). Vitamin B6 increases the immediate conversion of HCY to a glutathione precursor, cysteine. The deficiency of vitamin B6 leads to oxidative stress and increases the levels of HCY (Obeid et al., 2009). Hence, a lot of reasons have been associated with hyperhomocysteinemia in PD patients taking L-dopa other than a decrease in the levels of SAM in blood and CSF (Cheng et al., 1997).
These reasons consist of the following: (1) there is an increased synthesis of SAH when metabolism of L-dopa produces catechol-O-methyltransferase (COMT), which decreases SAM levels and reduces SAM/SAH ratio; (2) the decreased enzymatic activity of MTHFR; and (3) the deficiency of folic acid or vitamin B12 (dos Santos et al., 2009; Muller and Muhlack, 2009). In this regard, several clinical trials have been done using decarboxylase (carbidopa) and COMT (entacaptone/tolcapone) inhibitors and using vitamin B12 and folate supplements to decrease the levels of HCY in PD patients (Zoccolella et al., 2007b).
Epigenetic study of ADORA2A gene
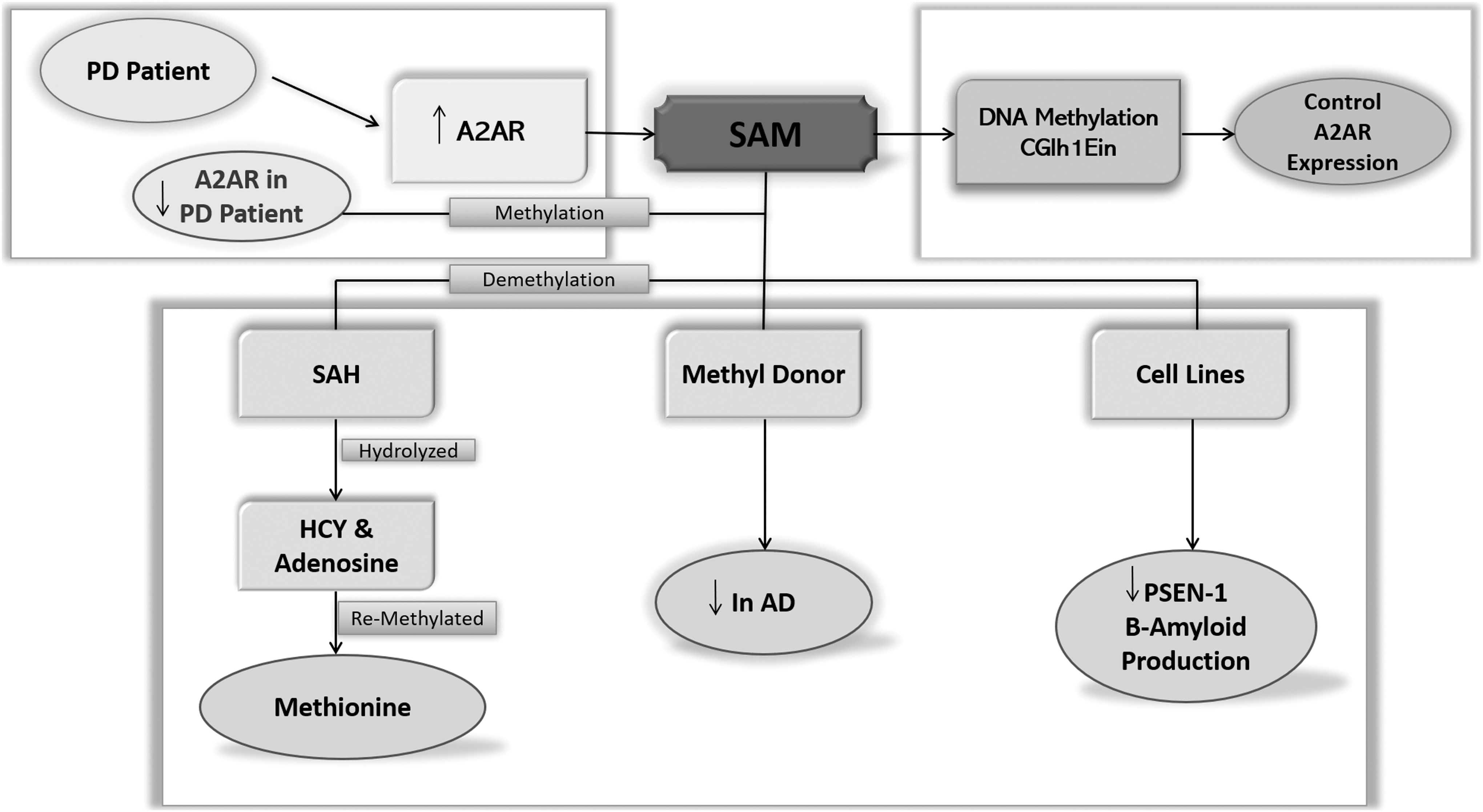
Variable expression of human adenosine A2A receptor gene (ADORA2A) that has been found in the granulocytes of sepsis patients with 5′ untranslated region (5′ UTR) of this gene is considered to play vital regulatory functions in its expression (Kreth et al., 2008). Later on, studies of different cell lines have shown a functional CGI adjacent to the h1E isoform, which reflects the role of DNA methylation in controlling the basal expression of the ADORA2A gene in a variety of cell lines. This molecular mechanism is responsible for differential expression of A2A receptor gene in different regions of the human brain like cerebellum and putamen (Buira et al., 2010a).
The expression level of A2A receptor gene can be modified in U87MG (human glioblastoma) and SH-SY5Y (human neuroblastoma) cell lines by SAM therapy. Because higher expression levels of A2A receptor are present in PD patients, (Varani et al., 2010) SAM therapy can be tried as a vital therapeutic tool in these patients to decrease the expression of A2A receptors by increasing the methylation of the ADORA2A gene (Buira et al., 2010b). Chronic therapy with A2A receptor antagonist istradefylline as an adjunct to L-DOPA (IST-LD) for PD patients revealed to restore higher demethylation of ADOA2AR, resulting in reduced expression of A2A receptor in the striatum (Kanzato et al., 2020).
Another recent study has used epigenesis-based tools to increase the metformin response in type 2 diabetes mellitus using DNA methylation of metformin transporter-associated gene (SLC 22A1 and SLC47A1), and it showed an improved drug response, but safety concerns of to this epigenetic tool should be further investigated (García-Calzón et al., 2020).
Clinical trial with SAM
DNA hypomethylation in hyperhomocysteinemic PD patients is considered to be accountable for a generalized dysregulation in the gene expression and adverse outcomes of the disease pathology. Previous studies have shown an improved cognitive function in PD patients by increasing the levels of vitamin B6 and by increasing the SAM/SAH ratio (Obeid et al., 2009). A clinical trial has shown that SAM therapy can reduce depression in PD patients (Di Rocco et al., 2000). In AD patients at the initial and moderate stages of the illness, SAM treatment, including dietary vitamin supplements, has been effective (Chan et al., 2008; Remington et al., 2009).
In addition, SAM supplementation has shown antioxidant properties in animal models of AD (Cavallaro et al., 2010). As oxidative stress is the hallmark of PD at the initial stage (Ferrer et al., 2011), SAM supplementation can be used as an effective therapy in PD and AD as shown in figure 9.
Schematic diagram showing epigenetic modulation of A2A receptors.
Conclusion and Future Perspectives
This review intends to describe the role of adenosine signaling and their clinical applications as a therapeutic tool for the treatment of neurological diseases (AD, PD, ALS, MS, and epilepsy). Inactivation of A2A receptor or intake of caffeine in AD patients has resulted in the prevention of memory deficits and also associated with improved cognitive functions. Higher expressions of A2A receptors are accountable for the damage to the striatal dopaminergic neurons in PD patients, and its antagonist, istradefylline, is already in the clinical trial to analyze its efficacy. In MS, activation of A1 and P2Y12 receptors resulted in the blockade of P2X7 receptor expression, which leads to remyelination. In ALS patients, P2X7 receptors perform an important role in the activation of microglia and inflammation initiation, which result in the killing of motor neurons.
Thus, modulation of A2A and P2X7 receptor functions in pathogenesis of PD and ALS provides an effective treatment option. Adenosine reduces seizures in epilepsy patients, mainly through the activity of A1 receptors, inhibiting glutamatergic transmission. It is noteworthy that A2A receptor inactivation increased the affinity of D2 receptors for dopamine. This could be the main reason behind the prodopaminergic effects, which were observed in most of the clinical trials of PD patients treated with antagonists of A2A receptors.
In this review, we have also discussed the literature based on different studies and clinical trials focusing on the transcriptional regulation of the ADORA2A gene. These studies indicate that SAM therapy is a highly successful epigenetic tool in modulating the enhanced expression of A2A receptor in PD patients. Several clinical trials have reported limited side effects of SAM therapy, which must be further evaluated.
However, knowing that the restricted expression of A2A receptors is in the brain (which is up to 95% in striatopallidal MSNs), it can be concluded that the SAM therapy will have a “specific” targeted effect on A2A receptors in the brain. This might be particularly true in the cerebral regions of the brain where A2A receptors are co-localized with D2 receptors, which in turn show a decreased activity due to reduced dopamine levels in PD patients. Although SAM therapy may decrease the hypomethylated gene expression, it will lead to substantial activation of D2 receptors through its effect on A2A receptors.
Footnotes
Acknowledgment
We thank the members of the Department of Clinical Laboratory Sciences for helpful discussions.
Authors' Contributions
Conceptualization, M.A. and A.A.; resources, M.A and F.N.; writing—original draft preparation, M.A., A.A. and F.N.; writing—review and editing, M.I., M.I., M.I.U., A.A.M.A., T.A.G., and Q.R.; supervision, M.A. and A.A.; and project administration, M.A. and A.A. All authors have read and agreed to the published version of the article.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
No funding was received for this article.