
Abstract
Pluripotent stem cells (PSCs) have the ability of self-renewal that can retain the characteristics of the mother cell, and of pluripotency that can differentiate into several body types. PSCs typically include embryonic stem cells (ESCs) derived from the inner cell mass of the preimplantation embryo, and epiblast stem cells (EpiSCs) derived from the epiblast of postimplantation embryo. Although PSCs are able to be used by differentiation into endothelial cells as a potential treatment for vascular diseases, human ESCs and induced PSCs (iPSCs) are followed by ethical and safety issues.
Pigs are anatomically and physiologically similar to humans. Therefore, the goal of this study was to establish an efficient protocol that differentiates porcine EpiSCs (pEpiSCs) into the endothelial cells for applying the treatment of human vascular diseases. As a result, alkaline phosphatase (AP)-negative (−) pEpiSCs cultured in endothelial cell growth basal medium-2 (EBM-2) differentiation medium in association with 50 ng/mL of vascular endothelial growth factor (VEGF) for 8 days were changed morphologically like the feature of endothelial cells, and expression of pluripotency-associated markers (OCT-3/4, NANOG, SOX2, and C-MYC) in porcine differentiated cells was significantly decreased (p < 0.05). Additionally, when pEpiSCs were cultured in EBM-2 + 50 ng/mL of VEGF, porcine differentiated cells represented a common endothelial cell marker positive (CD31+) but monocytes and lymphocytes marker negative (CD45-). Therefore, these results indicated that pEpiSCs cultured in EBM-2 + 50 ng/mL of VEGF culture condition were efficiently differentiated into endothelial cells for the treatment of blood vessel diseases.
Introduction
Pluripotent stem cells (PSCs), including embryonic stem cells (ESCs), epiblast stem cells (EpiSCs), and induced PSCs (iPSCs) have the potential of self-renewal, and to differentiate into any cell type (endoderm, mesoderm, and ectoderm) in the adult body (Brons et al., 2007; Evans and Kaufman, 1981; Takahashi and Yamanaka, 2006). In the mouse, ESCs were derived from the inner cell mass of a preimplantation embryo at E3.5 (Evans and Kaufman, 1981; Martin, 1981). Since then, ESCs were able to induce successfully from primate species such as monkey (Thomson et al., 1995) and human (Thomson et al., 1998). EpiSCs were derived from the late epiblast presented in a postimplantation embryo at E5.5–E7.5 in mouse.
iPSCs were able to be generated by introduction of four reprogramming factors (OCT-3/4, SOX2, KLF4, and C-MYC) in mouse fibroblasts (Takahashi and Yamanaka, 2006) and terminally differentiated cells (Aoi, 2008; Hanna et al., 2008). These PSCs are potentially available for therapeutic purpose as well as disease model and tissue engineering for human.
In particular, human ESCs and EpiSCs might have high potentials for cell therapies to treat human chronic diseases. However, there are still many problems with actual uses of treatment for human diseases, such as ethical problems, efficient production of differentiated cells, and cell sorting (Klimanskaya et al., 2008; Lo and Parham, 2009). Although iPSCs are able to avoid ethical problems due to be derived from human differentiated somatic cells, there are potential dangers because they have been induced by such methods as gene modification techniques (Zacharias et al., 2011).
The inner surface layer (lining) of the entire circulatory system, including arteries, veins, and capillaries, is called endothelium, and the cells that make it up are called endothelial cells. They are critical to maintain permeability of vessel wall, and to regulate angiogenesis and blood flow (Michiels, 2003). Dysfunction of the endothelial cells provides serious pathogenic causes, such as heart attacks and strokes in the cardiovascular system (Gimbrone et al., 2000; Hansson et al., 2005).
Blood vessels are generally produced through two distinct processes, angiogenesis and vasculogenesis. Angiogenesis occurs by forming new blood vessels through the migration and proliferation of preexisting vascular cells (Carmeliet, 2000), as opposed to vasculogenesis, by which blood vessels are formed in the embryo and by the new generation of endothelial cells (Patan S., 2004). Therefore, embryonic cells and ESCs could be potential clinical sources specifically for the induction of vascularization in therapeutic application. It has been shown that early endothelial progenitor cells derived from mouse ESCs are able to produce blood vessel components, including endothelial, hematopoietic, and smooth muscle cells (Yamashita et al., 2000).
On the other hand, CD31, namely platelet/endothelial cell adhesion molecule-1 (PECAM-1), is an adhesion molecule that was identified mostly in endothelial cells, platelets, and blood leukocytes (Li and Guo-Ping, 2012; Liu and Shi, 2012). Vascular endothelial growth factor (VEGF) in developing adult tissues is a well-established mitogen, as differentiation and survival factors of endothelium (Olsson et al., 2006). Additionally, VEGF has the ability to promote the proliferation of endothelial cells (Ferrara, 1999). After all the above, CD31 and VEGF play important roles specifically in angiogenesis and vasculogenesis.
On the contrary, CD45 is commonly expressed on all differentiated hematopoietic cells, not including erythrocytes and platelets. So it is often considered as a marker for cells of hematopoietic origin and commonly used to isolate leukocytes. Interestingly, CD45 is not normally expressed on endothelium or endothelial cells (Bischoff et al., 2016). Therefore, PSCs could be of use as a source of endothelial cells for the treatment of severe vascular diseases of people with diabetes or Alzheimer's and life-threatening strokes (Weiss et al., 2009). It has been reported that the differentiation of human ESCs and iPSCs using growth factors gives rise to tissue-specific microvascular endothelial cells (Wilson et al., 2014; Yoder, 2015).
None the less, there are many problems such as ethical and stability issues to apply the endothelial cells differentiated by human ESCs and iPSCs for the treatment of human vascular diseases. To solve these problems, endothelial cells might be differentiated using porcine PSCs to develop appropriate alternative products for the treatment of vascular diseases in humans.
In the case of pigs in domestic animals, they have anatomical and physiological similarities with humans (Nowak-Imialek et al., 2011). Therefore, porcine PSCs are of considerable importance as biomedical models for the treatment of human diseases (Kobayashi et al., 2017). The establishment of PSCs from pigs is significantly important to use as cell therapy for human diseases in the future. Until now, however, the porcine ESCs have not been established and specific markers and incubation conditions for the porcine ESCs have not been clarified (Back et al., 2019; Chen et al., 1999; Li et al., 2003; Notarianni et al., 1990; Piedrahita et al., 1990).
In the present study, we purposed to differentiate EpiSCs as one of PSCs into vascular endothelial cells for the treatment of vascular diseases. So, we have examined that differentiation medium induces the differentiation of porcine EpiSCs into vascular endothelial cells. Also, whether the addition of VEGF to differentiation medium improves the efficiency of differentiation of the porcine EpiSCs into endothelial cells was examined. These studies have established an efficient protocol that differentiates endothelial cells from porcine EpiSCs with two-dimensional culture.
Materials and Methods
All chemicals and reagents were purchased from Sigma-Aldrich Co. (St. Louis, MO) unless otherwise stated.
Culture for porcine EpiSCs
Porcine epiblast production, isolation, and stem cell culture were performed as previously described by Baek et al. (2019). Porcine epiblast stem cells (pEpiSCs) were cultured on mitomycin C-inactivated mouse embryonic fibroblast (iMEF) cells in Dulbecco's modified Eagle's medium (DMEM/F12; GIBCO) supplemented with 1% MEM nonessential amino acids, 2 mM L-glutamine, 1% penicillin/streptomycin, 0.1 mM β-mercaptoethanol, 20% fetal bovine serum (GIBCO Lot No. 2039230), 5 ng/mL basic fibroblast growth factor (bFGF; R&D), and 10 μM Y-27632 (Enzo). MEF cells were inactivated by 10 μg/mL mitomycin C (DUCHEFA) at passage 3. pEpiSCs were passaged onto iMEFs by enzymatic passaging every 3–5 days. Enzymatic passaging was used as a mixture of 1 mg/mL Collagenase type IV and 1 mg/mL Dispase (GIBCO) and incubated for 10 minutes at 39°C.
Alkaline phosphatase activity
The alkaline phosphatase (AP) staining was performed using an AP Detection Kit (Chemicon/Millipore) according to the manufacturer's protocol. Briefly, pEpiSCs 2 line and 8 line were fixed with 4% paraformaldehyde (PFA) for 1–2 minutes and then washed three times with tris buffered saline with Tween-20 (TBS-T) (20 mM Trics-HCl, pH 7.4, 0.15 M NaCl, 0.05% Tween-20). These pEpiSCs 2 line and 8 line were stained with stain solution (the ratio of Fast Red Violet/Naphthol AS-BI phosphate solution/water = 1:2:1) for 15 minutes in dark at room temperature. After the staining, they were washed with TBS-T and then covered with PBS to prevent drying. Images were observed with LEICA microscope (LEICA, Germany, type 090-135 001) and captured by NIS Elements microscope imaging software (Nikon, Japan, version 3.0).
In vitro differentiation of pEpiSCs
For the differentiation of pEpiSCs, pEpiSCs [AP-negative (−) pEpiSCs 8 line] were passaged by manually picking the colonies and then seeded onto 0.5% gelatin-coated culture plates in four differentiation media [endothelial cell growth basal medium 2 containing the BulletKit supplement, excluding VEGF (EBM-2, Lonza), EBM-2 supplemented with 50 ng/mL (EBM-2+VEGF, R&D), StemDiff APEL™ 2 medium (APEL-2, STEMCELL Technologies), and StemDiff APEL 2 medium supplemented with 50 ng/mL VEGF (APEL-2+VEGF)] for 8 days at 39°C. These media were changed every 3 days for 8 days.
Quantitative real-time polymerase chain reaction
Total RNAs of cells differentiated from pEpiSCs [AP-negative (−) pEpiSCs 8 line] were extracted using the RNeasy Plus Mini Kit (Qiagen) following the manufacturer's instructions. cDNA was synthesized using the Revoscript™ RT Premix (iNtRON Biotechnology, Inc.).
Total RNAs and cDNA were measured by MaestroNano® Spectrophotometer (MAESTROGEN). Quantitative real-time polymerase chain reaction (q-PCR) was executed using the THUNDERBIRD® SYBR® qPCR Mix (TOYOBO) on the Rotor-Gene Q—Pure Detection system (QIAGEN). The conditions were followed: predenaturation for 60 seconds at 95°C, denaturation for 15 seconds at 95°C, and extinction for 60 seconds at 60°C for 40 cycles. Data analysis was done by the ΔΔCt method and gene expression was standardized relative to the reference gene (18S). The primer list used for q-PCR is represented in Table 1.
Quantitative Real-Time Polymerase Chain Reaction Primer Lists Used in This Study
Immunocytochemistry
pEpiSCs [AP-negative (−) pEpiSCs 8 line] cultured in differentiation medium were fixed with 4% PFA for 20 minutes at 4°C and then washed with PBS-T three times for 5 minutes. After being washed in PBS-T, the cells were treated with the blocking solution (5% bovine serum albumin [BSA] in PBS-T) for 1 hour at room temperature. In addition, the cover slips were incubated overnight with primary antibodies in blocking solution at 4°C using OCT-3/4 (1:100; Santacruz, cat. no. #sc-6828), NANOG (1:100; Abcam, cat. no. #500-p237), SOX2 (10 ng/mL, R&D System, cat. no. #MAB2018), and CD31 (1:100, Novusbio, cat. no. #NB100-2284).
After overnight, the cells were washed with PBS-T three times for 5 minutes and then incubated with secondary antibodies in blocking solution at room temperature for 1 hour; Alexa Fluor® 568 Donkey anti-Goat IgG (1:150; Invitrogen), Alexa Fluor 546 Goat anti-Rabbit IgG (1:150; Invitrogen), and Alexa Fluor 555 Donkey anti-Mouse IgG (1:150; Invitrogen). To indicate the nuclei in cells, the cells were treated with 5 μg/mL of Hoechst 33342 in PBS at room temperature for 15 minutes. All images were explored using the LEICA fluorescence microscope (LEICA, DM 2500) and performed with the Leica Application Suite (LAS) (LEICA, version 2.7).
Western blots
Proteins in whole cells were reacquired using Lysis buffer supplemented with protease/phosphatase inhibitor (Cell signaling technology). After the collected cells were incubated in ice for 30 minutes, they were centrifuged to 15,000 rpm for 15 minutes at 4°C. After the centrifuge, only supernatant of the tube was collected. The harvested proteins were quantified using Bio-Rad Protein Assay Dye Reagent Concentrate (Bio-Rad). Proteins harvested from cells were separated on sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene fluoride (PVDF) membrane (Bio-Rad). After being blocked with 5% skim milk (Bioshop) for 1 hour, the membrane was washed three times with PBS-T for 15 minutes.
The membranes were incubated with specific primary antibodies overnight at 4°C in the following condition; CD31 (1:1000; Novusbio), CD45 (1:1000; Novusbio), and β-actin (1:4000; Santacruz). The membrane was washed three times with PBS-T for 15 minutes and then incubated with the secondary antibody in 5% skim milk at 4°C for 3 to 5 hours in the following condition; Goat anti-Rabbit IgG (H&L) (1:1000; Tonbo Biosciences) and Goat anti-Mouse IgG (H&L) (1:2000; Tonbo Biosciences).
Horseradish peroxidase-conjugated secondary antibody was used. After the membrane was washed three times with PBS-T for 15 minutes, Immunocomplexes were detected by enhanced chemiluminescence (Bio-Rad). ChemiDoc™ XRS+ System (Bio-Rad) to detect protein by being exposed for 1 hour was used. Bio RAD The Discovery Series Quantity One 10D Analysis software (Bio-Rad) was used to analyze.
Fluorescence-activated cell sorting (FACS) analysis
Differentiated porcine cells derived from pEpiSCs [AP-negative (−) pEpiSCs 8 line] were cultured in tissue culture medium and then washed with PBS (w/o Ca2+ or Mg2+) at room temperature. These cells were treated with 0.05% Trypsin ethylenediaminetetraacetic acid (EDTA) at 39°C for 5 minutes. The trypsin-treated cells were collected on the tube using PBS and then centrifuged at 300 g for 5 minutes. The cells were resuspended in stain buffer (1 × PBS, 2% BSA, 0.1% NaN3, pH 7.1–7.4). Then the single cells were stained with fluorescently conjugated antibody [CD31-PE (1:100, BD Pharmigen™)] for flow cytometry and incubated for 20–45 minutes on ice in the dark. After the incubation, the cell pellet was washed twice with stain buffer and then resuspended in stain buffer. The stained cell samples were analyzed by fluorescence-activated cell sorting (FACS) verse™ (BD Biosciences) and FlowJo.
Karyotyping analysis
pEpiSCs [AP-negative (−) pEpiSCs 8 line] and differentiated porcine cells derived from AP-negative (−) pEpiSCs 8 line at passage 30 cultured in EBM-2+VEGF differentiated medium were treated with 10 μL/mL colcemide (Biological industries kibbutz Beit Haemek) at 37°C for 1 hour. After being incubated, all of the cells were arrested in the metaphase stage. Single cells harvested by 0.05% Trypsin/EDTA were incubated with hypotonic solution (0.4% KCl and 0.4% NaCl in H2O) at 39°C for 6 minutes and then fixed by fixation solution (ratio of methanol: glacial acetic acid = 3:1). The cell pellets were resuspended in a fixation solution, and then dropped on a cold slide and dried.
Statistical analysis
At least three replicates were measured for each group. Statistical significance was assessed by one-way analysis of variance (ANOVA) and t-test to examine differences between the two groups using GraphPad Prism software v7.00 (GraphPad) were used to test the significance of the data. p-Values of <0.05 were considered statistically significant.
Results
AP activity of pEpiSCs
While pEpiSCs (pEpiSCs 2 line and 8 line) were incubated on the iMEF, colonies derived from both pEpiSCs 2 line and 8 line were formed from day 2 (Fig. 1A). When pEpiSCs 2 line and 8 line with AP staining were compared, pEpiSCs 2 line represented AP-positive (+) staining but pEpiSCs 8 line showed AP-negative (−) staining at passage 2. Additionally, pEpiSCs 2 line showed AP-positive (+) staining, but pEpiSCs 8 line represented AP-negative (−) staining for passages 10, 20, and 30 (Fig. 1B). Among the two lines, AP-negative (−) pEpiSCs 8 line was selected for the differentiation of vascular endothelial cells because they showed higher efficiency of differentiation of pCardiac actin (mesoderm) as previously published by Baek et al. (2020).

Morphology and AP activity in pEpiSCs (2 line and 8 line).
Morphology of differentiated porcine cells derived from pEpiSCs
When AP-negative (−) pEpiSCs 8 line was incubated in four different differentiation media (EBM-2, EBM-2 + 50 ng/mL VEGF, APEL-2, and APEL-2 + 50 ng/mL VEGF) on 0.5% gelatin-coated plate for 8 days, morphological changes in differentiated porcine cells were examined (Fig. 2B). While AP-negative (−) pEpiSCs 8 line was cultured in stem cell culture medium, the morphological change in these pEpiSCs was not observed but these cells have shown continuously self-renewal.
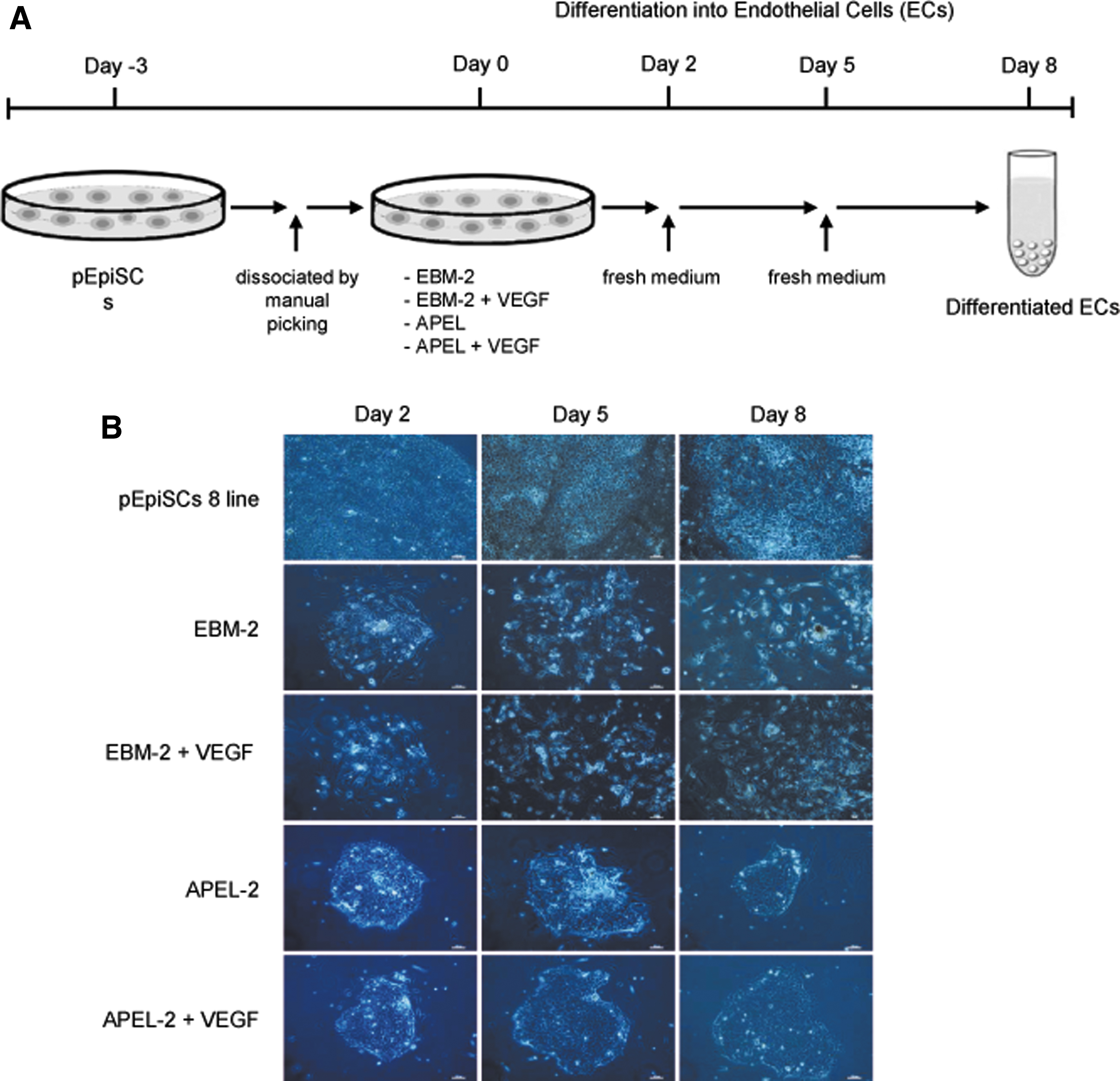
Morphologies of pEpiSCs cultured in differentiation media.
However, as these pEpiSCs were cultured in EBM-2 and EBM-2 + 50 ng/mL VEGF, morphological changes in pEpiSCs were observed from day 2 and morphological similarity like endothelial cells was represented at day 8 (Fig. 2B). On the contrary, when these pEpiSCs were cultured in APEL-2 and APEL-2 + 50 ng/mL VEGF, the morphology of these cells has not changed.
Expression of pluripotency-associated genes in differentiated porcine cells derived from pEpiSCs
While AP-negative (−) pEpiSCs 8 line was cultured in four different differentiation media on 0.5% gelatin-coated plate for 8 days, expression changes of pluripotency-associated genes (OCT-3/4, NANOG, SOX2, and C-MYC) in these differentiated porcine cells were compared (Fig. 3). There was no difference in expression of pluripotency-associated genes between EBM-2 medium and APEL-2 medium. When 50 ng/mL of VEGF was added to EBM-2 medium, the expression of the pluripotency-associated genes (OCT-3/4, NANOG, and SOX2) in these differentiated cells were significantly reduced than these differentiated cells incubated in EBM-2 and APEL-2 (p < 0.05) (Fig. 3). However, differentiated porcine cells cultured in APEL-2 + 50 ng/mL VEGF for 8 days did not induce decreasing OCT-3/4, NANOG, and SOX2 expressions. On the other hand, the expression of C-MYC in differentiated porcine cells incubated with APEL-2 and APEL-2 + 50 ng/mL VEGF was significantly decreased as compared with AP-negative (−) pEpiSCs 8 line and differentiated cells cultured in EBM-2 and EBM-2 + 50 ng/mL VEGF medium (p < 0.05).

Expression of pluripotency-associated genes in porcine differentiated cells derived from pEpiSCs cultured in four differentiation media. Relative mRNA level of pluripotency-associated genes (OCT-3/4, NANOG, SOX2, and C-MYC) in pEpiSCs [AP-negative (−) pEpiSCs 8 line] at passage 30 cultured in different differentiation media [endothelial cell growth basal medium-2 (EBM-2), endothelial cell growth basal medium-2 supplemented with 50 ng/mL VEGF (EBM-2+VEGF), APEL-2, APEL-2 supplemented with 50 ng/mL VEGF (APEL-2+VEGF)]. Columns, mean of three independent experiments performed in triplicate; bar, SEM; *p < 0.05 and **p < 0.01 versus pEpiSCs 8 line. SEM, standard error of the mean.
Expression of pluripotency-associated proteins in differentiated porcine cells derived from pEpiSCs
As long as PSCs differentiate into any cell type in the body, the pluripotency in PSCs should be reduced or lost. When AP-negative (−) pEpiSCs 8 line was incubated with differentiation media (EBM-2, EBM-2 + 50 ng/mL VEGF, APEL-2, and APEL-2 + 50 ng/mL VEGF) on 0.5% gelatin-coated plate for 8 days, changes in expression of the pluripotency-associated proteins (OCT-3/4, NANOG, and SOX2) of differentiated porcine cells were examined (Fig. 4).

Immunocytochemistry of pluripotency-associated markers in porcine differentiated cells derived from pEpiSCs cultured in differentiation media. Immunostaining images of pluripotency-associated markers (OCT-3/4, NANOG, and SOX2) in porcine differentiated cells derived from pEpiSCs [AP-negative (−) pEpiSCs 8 line] cultured in four differentiation media [endothelial cell growth basal medium-2 (EBM-2), endothelial cell growth basal medium-2 supplemented with 50 ng/mL VEGF (EBM-2+VEGF), APEL-2, APEL-2 supplemented with 50 ng/mL VEGF (APEL-2+VEGF)] on 0.5% gelatin-coated plate for 8 days. Blue: Hoechst 33342 signal for nuclei. Red: Staining of pluripotency-associated marker (OCT-3/4, NANOG, and SOX2) proteins. Merge: Hoechst 33342 signal and OCT-3/4, NANOG, and SOX2 proteins. Scale bar = 50 μm. Color images are available online.
As a result, the pEpiSCs were positively immunostained with pluripotency-associated proteins (OCT-3/4, NANOG, and SOX2). However, the expression of OCT-3/4, NANOG, and SOX2 proteins of differentiated porcine cells derived from AP-negative (−) pEpiSCs 8 line cultured with EBM-2 and EBM-2 supplemented with 50 ng/mL of VEGF were decreased considerably. However, when these pEpiSCs were cultivated in APEL-2 and APEL-2 supplemented with 50 ng/mL of VEGF, the fluorescence of pluripotency-associated proteins did not represent difference from those of AP-negative (−) pEpiSCs 8 line (Fig. 4).
Expression of endothelial cell marker (CD31) in differentiated porcine cells derived from pEpiSCs
The expression of a specific marker (CD31) of vascular endothelial cells was tested in differentiated porcine cells derived from AP-negative (−) pEpiSCs 8 line cultured in four different differentiation media (EBM-2, EBM-2 + 50 ng/mL of VEGF, APEL-2, and APEL-2 + 50 ng/mL of VEGF) (Fig. 5).
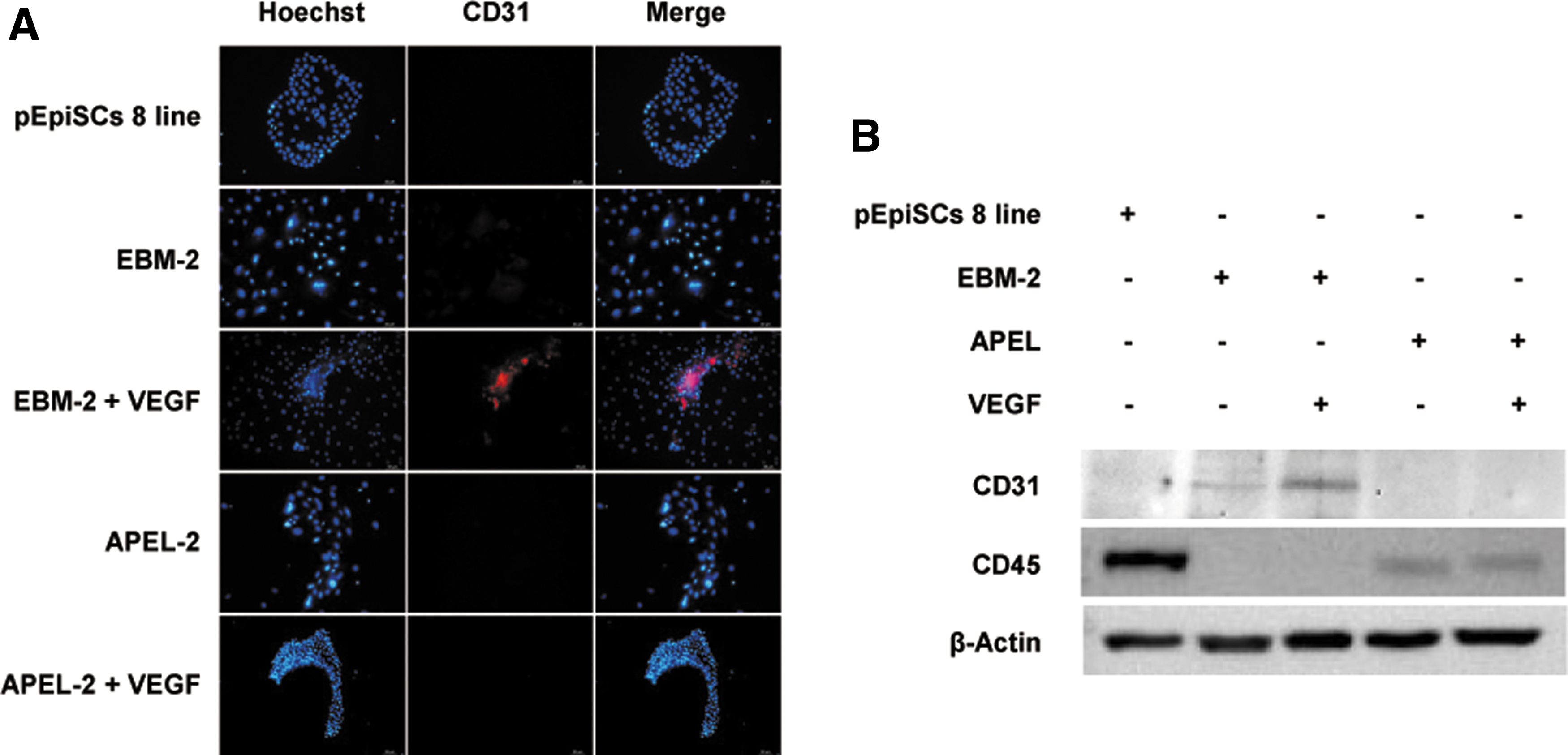
Expression of endothelial cell marker (CD31) protein of porcine differentiated cells derived from pEpiSCs cultured in differentiation media. Immunocytochemistry
AP-negative (−) pEpiSCs 8 line and differentiated porcine cells derived from pEpiSCs cultured in APEL-2 and APEL-2 + 50 ng/mL of VEGF did not express CD31 protein. However, differentiated porcine cells derived from AP-negative (−) pEpiSCs 8 line incubated with EBM-2 and EBM-2 + 50 ng/mL of VEGF expressed the protein of CD31 (Fig. 5A). Especially, when 50 ng/mL of VEGF was added to the differentiation medium of EBM-2, the expression of CD31 increased significantly to compare with the differentiated porcine cells cultured in EBM-2 medium. Like the result of immunofluorescence, the expression of CD31 as a marker of vascular endothelial cells was observed higher in the differentiated porcine cells derived from pEpiSCs cultured in EBM-2 + 50 ng/mL of VEGF.
However, CD45 as a monocyte and lymphocyte marker was not detected in the differentiated porcine cells derived from pEpiSCs cultured in EBM-2 + 50 ng/mL of VEGF (Fig. 5B). Therefore, it was verified that AP-negative (−) pEpiSCs 8 line were specifically induced to differentiate into vascular endothelial cells by introducing VEGF to the EBM-2 media.
Flow cytometry analysis of porcine endothelial cells differentiated from pEpiSCs
To examine the rate of differentiation of AP-negative (−) pEpiSCs 8 line into vascular endothelial cells using flow cytometry analysis, these stem cells were cultured in four different differentiation media (EBM-2, EBM-2 + 50 ng/mL of VEGF, APEL-2, and APEL-2 + 50 ng/mL of VEGF) (Fig. 6). When these pEpiSCs were incubated with differentiation media of EBM-2 and APEL-2 for 8 days, the efficiency of differentiation into endothelial cells was ∼7.32% and 3.96%, respectively (Fig. 6A). Noteworthy, when 50 ng/mL of VEGF was added to EBM-2 and APEL-2 differentiation medium, the rate of differentiation into endothelial cells was found to be about 27.4% and 3.83%, respectively.

Flow cytometry analysis of endothelial cell (CD31) protein in porcine differentiated cells derived from pEpiSCs cultured in differentiation media. Flow cytometry analysis
These results indicate that the efficiency of differentiation of AP-negative (−) pEpiSCs 8 line into vascular endothelial cells increases only with the addition of 50 ng/mL VEGF to the EBM-2 differentiation medium.
Karyotype analysis of differentiated porcine cells derived from pEpiSCs
Karyotype analysis of differentiated porcine cells at passage 30 derived from the AP-negative (−) pEpiSCs 8 line incubated with the EBM-2 differentiation medium was carried out to prove the normal presence of the chromosomes. As a result, we have identified normal chromosomes (2N = 38) in the differentiated porcine cells derived from pEpiSCs cultured in EBM-2 + 50 ng/mL of VEGF (Fig. 6B).
Discussion
PSCs are the appropriate source of human therapies because they have the ability to self-renewal and pluripotency. Especially, pigs have physical and anatomical similarities with humans, so they are able to be properly applied as human therapies using porcine PSCs (Kobayashi et al., 2017; Nowak-lmialek et al., 2011). However, until now, the porcine PSCs have been not fully established, and no differentiation experiment was carried out to develop for human therapies (Chen et al., 1999; Li et al., 2003; Notarianni et al., 1990; Piedrahita et al., 1990). Therefore, we tried differentiation of pEpiSCs into endothelial cells to develop cellular therapies for human vascular diseases.
First of all, we have compared two pEpiSCs lines (pEpiSCs 2 and pEpiSCs 8) on the basis of AP activity. As a result, pEpiSCs 2 line represented almost positive AP activity [AP-positive (+) pEpiSCs 2 line] but pEpiSCs 8 line did not show at all [AP-negative (−) pEpiSCs 8 line]. Generally, high AP activity was reported in mouse ESCs (mESCs) and human ESCs (hESCs), whereas EpiSCs, which were developed later than ESCs, represented the activity of low AP. In our previous experiment, AP-negative (−) pEpiSCs 8 line induced strong multiple lineages (Cardiac actin, Nestin, and GATA 6) and primordial germ cells (Dazl and Vasa) in differentiated porcine cells, whereas AP-positive (+) pEpiSCs 2 line did not express (Baek et al., 2020).
Therefore, we chose the AP-negative (−) pEpiSCs 8 line to differentiate favorably into vascular endothelial cells for the development of cellular therapy in vascular diseases.
AP-negative (−) pEpiSCs 8 line was cultured in APEL-2 and EBM-2 differentiation medium to differentiate into endothelial cells for 8 days. APEL-2 was generally used with a variety of different induction factors or cytokines for inducing the differentiation of human induced pluripotent hESCs and human iPSCs (hiPSCs) into endothelial cells (Harding et al., 2017; Ng et al., 2008). It was also reported that EBM-2, an endothelial cell growth medium, has been used to differentiate hESCs into endothelial cells (Joddar et al., 2018; Nourse et al., 2010).
In this study, EBM-2 differentiation medium induced morphological change in differentiated porcine cells generated from AP-negative (−) pEpiSCs 8 line, whereas APEL-2 differentiation medium did not. These porcine differentiated cells were morphologically changed into the typical feature of vascular endothelial cells from the day 2 of incubation. In addition, VEGF induced the differentiation of functional endothelium from hESCs and hiPSCs for tissue engineering (Lin et al., 2012; Nourse et al., 2010).
In the present study, when 50 ng/mL of VEGF was added to EBM-2 and APEL-2, the morphological change in porcine differentiated cells derived from pEpiSCs was significantly observed in only EBM-2 differentiation medium. However, interestingly, VEGF did not affect the morphology in APEL-2 differentiation medium.
To differentiate PSCs into any cell type (endoderm, mesoderm, and ectoderm) in vitro, expression of pluripotency-associated genes (OCT-3/4, NANOG, and SOX2) in PSCs should be restricted and removed for avoiding heterogenetic induction of PSCs.
When examining the expression of pluripotency-associated genes of the AP-negative (−) pEpiSCs 8 line cultured in EBM-2 or APEL-2 differentiation medium for 8 days, two differentiation culture systems did not induce decreasing OCT-3/4, NANOG, and SOX2 expressions in differentiated porcine cells. However, when VEGF was added to EBM-2 culture condition, decreasing PSC markers (OCT-3/4, NANOG, SOX2, and C-MYC) were detected in these porcine differentiated cells. Additionally, the expression of OCT-3/4, NANOG, and SOX2 proteins was declined in EBM-2 and EBM-2 + 50 ng/mL VEGF culture conditions.
In particular, EBM-2 + 50 ng/mL VEGF significantly decreased the expression of PSC marker proteins. Like the above results, our results indicate that a successful culture system of porcine PSCs for differentiation into endothelial cells requires differentiation medium and growth factor.
To induce the differentiation of endothelial cells, we tested two differentiation culture conditions for porcine PSCs. Of the two culture conditions tested, EBM-2 differentiation medium resulted in the expression of the endothelial cell-surface marker CD31 (PECAM1), as determined by immunocytochemistry and western blotting. In particular, VEGF treatment for 8 days at 50 ng/mL resulted in the highest expression in differentiating endothelial cells. However, there is no expression of CD31 protein in APEL-2 and EBM-2 + 50 ng/mL VEGF culture conditions, suggesting that VEGF plays an important role in inducing endothelial cell differentiation when incubated with EBM-2 culture condition.
Moreover, pEpiSCs incubated with EBM-2 + 50 ng/mL VEGF resulted in ∼27% CD31 positive (+) of total cell population using flow cytometry analysis. It has been reported that when differentiation medium was cultured in association with VEGF, the efficiency of human endothelial cell derivation resulted in ∼20% of total cell population in hESCs and hiPSCs (Lin et al., 2012; Orlova et al., 2014). However, other differentiation culture conditions, EBM-2, APEL-2, and APEL-2 + 50 ng/mL VEGF, induced approximately 4%–7% CD31 positive (+) of total cell population in porcine PSCs. Finally, it was confirmed that all chromosomes in pEpiSCs 8 and differentiated porcine cells generated from AP-negative (−) pEpiSCs 8 line were normal (2N = 38) at EBM-2 + 50 ng/mL VEGF treatment.
In conclusion, we found that the differentiation of the AP-negative (−) pEpiSCs 8 line into vascular endothelial cells was induced in EBM-2 differentiation medium. Especially, when 50 ng/mL of VEGF was added to EBM-2 differentiation medium, expression of pluripotency-associated markers (OCT-3/4, NANOG, and SOX2) in porcine differentiated cells derived from AP-negative (−) pEpiSCs 8 line was dramatically decreased but the efficiency of differentiation into endothelial cells was significantly increased. The endothelial cells generated from the porcine PSCs would potentially be used for the treatment of human vascular diseases.
Footnotes
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
Funding Information
This work was supported by the National Research Foundation of Korea funded by the Korean Government (NRF-2017R1A2B1011438 and NRF-2018R1C1B6007728) Republic of Korea. S.-K.B. was supported by the scholarship from the BK21Plus Program, Ministry of Education, Republic of Korea.