
Abstract
Past researches have shown that pluripotency maintenance of naive and primed-state pluripotent stem cells (PSCs) depends on different signaling pathways, and naive-state PSCs possess the ability to produce chimeras when they are introduced into a blastocyst. Considering porcine is an attractive model for preclinical studies, many researches about pig induced pluripotent stem cells (piPSCs) have been reported. Some cytokines and small molecule compounds could transform primed piPSCs into naive state. However, there are no suitable culture conditions for generation of naive-state piPSCs with high efficiency; other small molecule compounds need further exploration. In this study, we investigated whether p38 MAPK and JNK signal pathway inhibitor SB203580 and SP600125 could be of benefit for acquiring naive-state piPSCs. By comparing reprogramming efficiencies under conditions of different donor cells and culture environment, we found that porcine bone marrow mesenchymal stem cells (PBMSCs) have higher efficiency on piPSC induction, and the culture condition of CHIR99021+PD0325901(2i)+Lif+bFGF is more suitable for subculturing of piPSCs. Our results also indicate that SB203580 and SP600125 could promote reprogramming of PBMSCs into naive-like state piPSCs. These results provide guidance for choosing donor cells, culture conditions, and research of different state iPSCs during the process of reprogramming pig somatic cells.
Introduction
Guangxi Bama mini-pig is an important livestock of China, mainly distributed in Bama Yao Autonomous County in Guangxi Zhuang Autonomous Region. It is also a valuable animal model in preclinical research, due to its anatomical and physiological similarity to human beings. Research of pluripotent stem cells (PSCs) from bama mini-pigs will provide important information in developmental biology, medical and transgenic animal production. So far, little is known about porcine embryonic stem cells (ESCs), which are derived from the inner cell mass of blastocysts and possess the capacity to self-renew and differentiate into all cell types of an organism. Many attempts have been made to establish authentic porcine ESC lines, but without successful results.
In 2006, Yamanaka and Takahashi first generated mouse induced pluripotent stem cells (iPSCs), similar to ESCs in many biological characteristics, by ectopic expression of four pluripotent genes Oct4, Sox2, Klf4, and c-Myc in somatic cells (Takahashi and Yamanaka, 2006). This method provided a new way to obtain livestock PSCs. Pig induced pluripotent stem cells (piPSCs) were first established by Wu (Wu et al., 2009) and two other groups with the same methods (Esteban et al., 2009; Ezashi et al., 2009), which laid the foundation for establishment of pig ESCs. Currently there exist many deficiencies in piPSCs (Kim et al., 2019; Setthawong et al., 2019; Su et al., 2020), such as donor cells, culture systems for piPSC induction, and passage, the pluripotency maintenance of piPSCs, and the difficulty of establishing stable piPSCs that could undergo unlimited passages, and there were few reports about the research on using piPSCs to give rise to chimeras. These problems suggest that the most existing piPSCs may be in a state of easier differentiation and have limited pluripotency, also known as “primed” state.
In 2009, Nichols and Smith found that mouse ESCs can be defined as naive and primed-state PSCs depending on the distinct phases of embryonic development from which they are derived (Nichols and Smith, 2009). It is now generally believed that differences between human and mouse ESCs may be reflected by the different properties of primed and naive state. Such naive-state PSCs are characterized by their morphology, the expression of marker genes like Klf2, Klf4, Klf5, Rex1, Stella, and Eras, tolerance to passage as single cells, dependence on LIF/STAT3 signaling for maintenance of pluripotency, no X chromosome inactivation in female cells, and ability to differentiate into cell types representing the three germ layers both in vitro and in vivo.
And most importantly, naive-state PSCs have the ability to give rise to germ-line chimeras and live offspring, which primed-state PSCs cannot (Honda et al., 2013; Park et al., 2018a; 2018b; Takahashi et al., 2018; Xu et al., 2019).
Naive-state piPSCs were previously generated in some laboratories with low efficiency (Fukuda et al., 2017; Li et al., 2018; Yuan et al., 2019). In 2014, Fang showed that a simple combination of cytokines and small molecule inhibitors could generate naive-state iPSCs from rhesus monkey fibroblasts with high efficiency (Fang et al., 2014). Among them, inhibitors SB203580 and SP600125 specifically inhibited p38 MAPK and JNK signaling pathways, respectively, which participate in varieties of cell biological activities, including cell apoptosis, differentiation, and adaptation of stress environment through competing with ATP to obtain the related binding sites.
However, whether they are beneficial for generation of naive-state piPSCs remain to be studied. Therefore, we used Dox-inducible lentiviral vectors encoded Oct4, Sox2, c-Myc, and Klf4 introduced into the porcine fetal fibroblasts (PFFs), porcine bone marrow mesenchymal stem cells (PBMSCs), and porcine adipose-derived stem cells (PASCs), which derived from Guangxi Bama mini-pig, to compare the efficiencies and identify the biological characteristics of piPSCs in different cells and different culture conditions, and to determine whether the two inhibitors SB203580 and SP600125 play similar roles in producing naive-like state iPSCs between rhesus monkeys and pigs.
Materials and Methods
Reagents and medium
The cell culture medium reagents were purchased from Gibco (Carlsbad, CA). Molecular detection reagents were purchased from Takara (Osaka, Japan). And the antibodies were obtained from Cell Signaling Technology (Danvers, MA). Other chemicals were purchased from Sigma-Aldrich (St. Louis, MO) unless otherwise stated. PFFs, PBMSCs, and PASCs were derived from Bama mini-pigs, which were provided by the Guangxi University Bama mini-pig breeding center. This study was conducted in accordance with the State Key Laboratory for Conservation and Utilization of Subtropical Agro-bioresources Guide for the Care and Use of Laboratory Animals.
Cell culture
The normal medium for cell culture consists of high-glucose Dulbecco's modified Eagle's medium (DMEM), 10% fetal bovine serum (FBS), and 1% penicillin-streptomycin; mTeSR induction medium consisted of mTeSR™ basic medium (Vancouver, BC, Canada), 1% penicillin-streptomycin, 2 μg/mL doxycycline (Dox), 2 mM VPA, and 10 ng/mL Y27632. Four mediums with different components (bFGF, 2i+Lif, 2i+Lif+bFGF, and 2i+Lif+bFGF+2s) were used for piPSC induction, consisted of high-glucose DMEM, 20% knockout serum replacement, 1% Penicillin-streptomycin, L-glutamine (100 × ), sodium pyruvate (100 × ), β-mercaptoethanol (100 × ), nonessential amino acids (100 × ), 2 μg/mL doxycycline, 2 mM VPA, 10 ng/mL Y27632, and 50 μg/mL Vc. Ten nanograms per milliliter bFGF, 10 ng/mL Lif, 1 μM PD0325901, 3 μM CHIR99021, 5 μM SB203580, and 5 μM SP600125 were added to the four different medium groups, respectively (2i: CHIR99021, PD0325901; 2s: SB203580, SP600125; VPA only used in 7 days after virus infection); freezing medium consisted of 90% FBS and 10% dimethyl sulfoxide.
The isolation, culture, and identification of PFFs, PBMSCs, and PASCs have been established and preserved in our laboratory (data not shown). Briefly, PFFs were obtained by adherent tissue method; PBMSCs were obtained from culture of bone marrow extract; and PASCs were obtained by collagenase digestion method. All cells were cultured under a humidified atmosphere of 5% CO2 in air at 37°C.
Inducible lentiviral transduction
PFFs, PBMSCs, and PASCs (passage 4) were transduced with a Dox Tet-on inducible lentiviral vector. Virus supernatants that contain the human transcription factors Oct4, Sox2, Klf4, and c-Myc (OSKM) were collected at 60 hours after transfection, and filtered through a 0.45 μm filter. Cells were infected by virus supernatant mixture, with 6 μg/mL polybrene to promote infection. Transduced cells were passaged onto a Matrigel-treated dish at 36 hours after virus infection and cultured in different mediums as mentioned above.
Alkaline phosphatase and immunocytochemistry
Alkaline phosphatase (AP) staining was carried out with the NBT/BCIP AP color development kit, following manufacturer's instructions.
Immunocytochemistry protocol was performed as previously reported (Park et al., 2018a). Primary antibodies were Oct4 (volume ratio = 1:200), Sox2 (volume ratio = 1:200), SSEA1 (volume ratio = 1:200), E-Cadherin (volume ratio = 1:200), TRA-1-60 (volume ratio = 1:200), Rex1 (volume ratio = 1:200), FGF5 (volume ratio = 1:200), and H3K27me3 (volume ratio = 1:250). Primary antibodies were detected using Alexa Fluor 555 Goat Anti-Mouse IgG or Goat Anti-Rabbit IgG (volume ratio = 1:250). Ten nanograms/mL Hoechst 333342 was used for nuclear staining.
RNA isolation and reverse transcription polymerase chain reaction (RT-PCR)
For assessment of endogenous and pluripotent gene expression, piPSCs were mechanically harvested from different donor cells and different culture conditions. For assessment of exogenous gene expression after removal of Dox, piPSCs were collected at P2, P4, P6, P8, and P10 after removal of Dox. For assessment, the expression levels of marker genes that represent the three germ layers, embryoid bodies (EBs) derived from piPSCs were harvested.
Total RNA was extracted using TRIzol reagent. Two micrograms of RNA was then amplified and converted into cDNA using the TranScript® One-step gDNA Removal and cDNA Synthesis SuperMix Kit following manufacturer's instructions. Electrophoresis was performed on a 2% agarose gel. Quantitative real-time polymerase chain reaction (QRT-PCR) was performed using SYBR Premix Ex Taq (Takara) and the 7500 Real-Time PCR System. The primers are listed in Supplementary Table S1.
Cell proliferation test
The cell growth curve assay was performed by manual counting (n = 3) every day for 8 days. Population doubling time was determined by the formula PDT = ▵tlg2/(lgNt−lgN0). To find out the suitable dosage of SB203580 and SP600125 for the following culture medium, cck-8 cell counting kit was used to test the effects of different concentrations of SB203580 and SP600125 on the proliferation of PBMSCs, according to the manufacturer's instructions. Statistical analysis was done using Duncan's test, with a p-value <0.05 being considered significant.
Karyotype analysis
When piPSCs reached 70% confluence, cells were treated with 0.2 μg/mL colchicine for 3–4 hours, followed by hypotonic treatment in 0.075M KCl solution at 37°C for 30 minutes, and then fixed in cold fixative (3:1 methanol and glacial acetic acid) for 2 minutes. We next centrifuged at 1000 rpm for 5 minutes, and added 5 mL cold fixative to fix 30 minutes, and the process was repeated once again. Finally, 300 μL of cold fixative was added to the cells, cell suspension was dropped onto cold slides at a vertical height of 60–80 cm, and then the slides were put to the outer flame of the alcohol lamp 3–5 times. After drying at room temperature, Giemsa dyeing solution was applied for 30 minutes and finally observed under an inverted microscope (Nikon, Tokyo, Japan). One hundred samples were selected for chromosome number analysis.
Sodium bisulfite genomic sequencing
Genomic DNA of PBMSCs and piPSCs was purified. The promoter region methylation level of Oct4 was examined using the Methylamp™ DNA Modification Kit according to the manufacturer's recommendations. After PCR and sequencing, the methylated CpG sites were analyzed. Details refer to our previous report (Deng et al., 2012). The PCR primer is listed in Supplementary Table S2.
EB formation
piPSCs were digested into single cells by trypsin, transferred into low adhesion dish, and cultured in induction medium without cytokines and small molecule compound. The medium was changed every other day and EBs were formed after 7 days of culture. EBs were harvested and analyzed by RT-PCR for expression of marker genes of three germ layers: endoderm (FOXA2), mesoderm (HAND1 and GATA2), and ectoderm (NESTIN and NCAM).
Results
PBMSCs are effective donor cells for piPSC induction
First, we explored the donor cells that can be efficiently used in piPSC induction (Fig. 1). After lentivirus infection, the iPSC colonies derived from PBMSCs, PASCs, and PFFs were generated from day 10 to 12 induction under mTeSR conditions (Fig. 1A). Results from colony morphology and AP staining assay showed that the reprogramming efficiencies of iPSCs derived from PBMSCs, PASCs, and PFFs were 0.28%, 0.25%, and 0.23%, respectively. The iPSC formation efficiencies derived from PBMSC group was significantly higher than PFF group (p < 0.05), but there was no significant difference between PASCs and PBMSCs, and PASCs and PFFs (Fig. 1B). When we picked colonies for subculturing in mTeSR medium without Dox, the colonies showed differentiation or apoptosis, or reverted back to its original cells (Fig. 1C). These results suggested that PBMSCs are effective donor cells for piPSC induction and mTeSR medium is not suitable for subculturing of piPSCs.
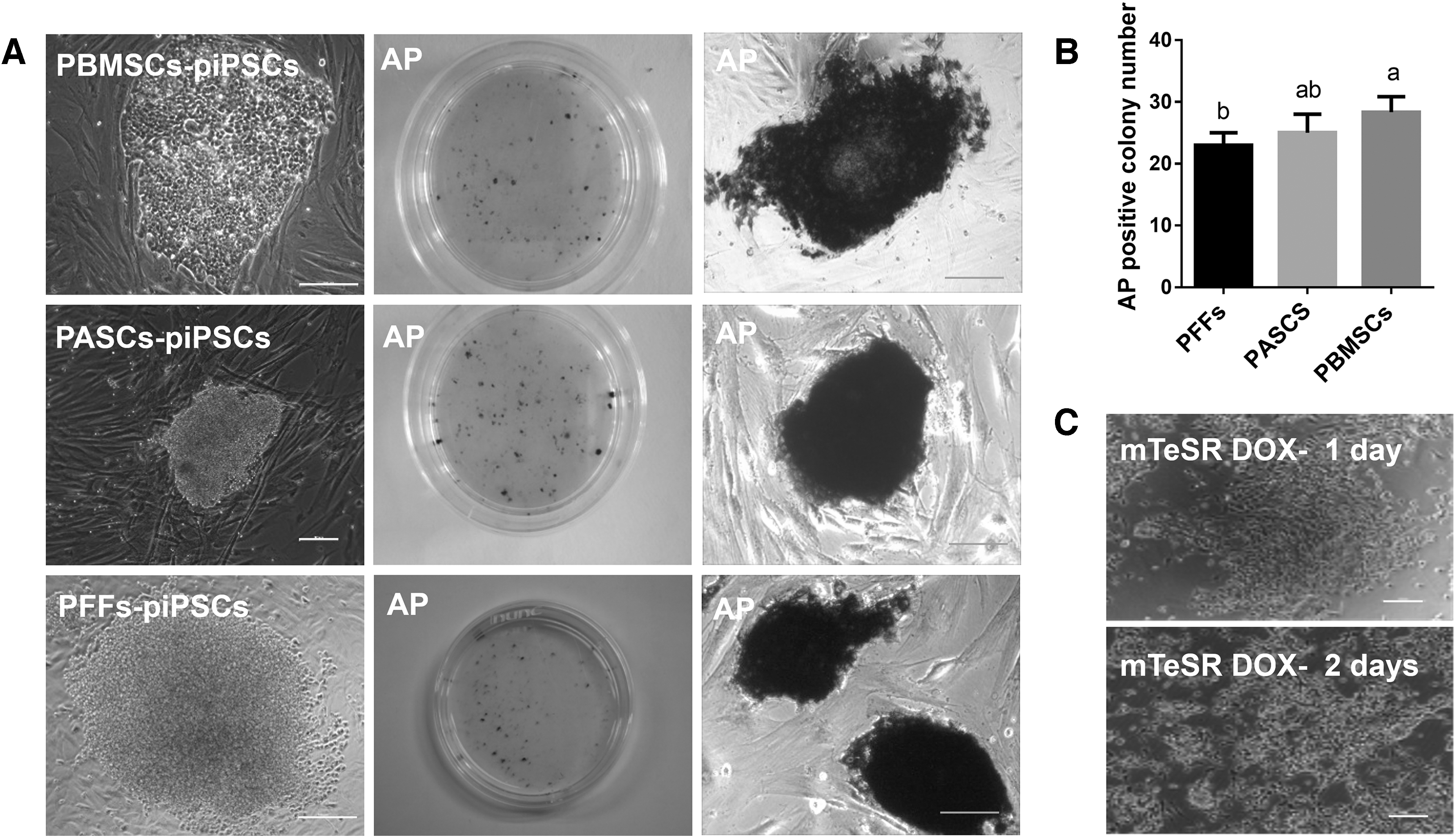
Comparing piPSC reprogramming efficiency of different donor cell induction in mTeSR culture medium (Scale bar = 100 μm).
2i+Lif+bFGF culture medium is more suitable for piPSC subculturing
To optimize the culture conditions, we chose PBMSCs as the donor cells, to compare the reprogramming efficiencies of iPSCs derived from PBMSC induction in three different culture medium (bFGF, 2i+Lif, and 2i+Lif+bFGF), respectively (Fig. 2). We found that the efficiencies of iPSC induction derived from PBMSCs in bFGF, 2i+Lif, and 2i+Lif+bFGF were 1.12%, 0.84%, and 1.07%, respectively. The efficiencies of iPSC colony formation in bFGF and 2i+Lif+bFGF group were significantly higher than 2i+Lif (p < 0.05), but there was no significant difference between bFGF and 2i+Lif+bFGF group (Fig. 2A, B).

Comparing piPSC reprogramming efficiency of PBMSC induction in different culture medium: bFGF, 2i+Lif, bFGF +2i+Lif (Scale bar = 100 μm).
To confirm the best conditions for iPSC subculturing, we picked piPSC colonies from different conditions and assayed the expression of endogenous pluripotent related genes. The results showed that the expression level of endogenous Oct4 and Klf4 from bFGF group was the lowest, and the colonies could not proliferate after passage 10. Colony morphology derived from 2i+Lif group was more round, but the cells proliferated slowly and stopped expanding after passage 6. The expression levels of endogenous Oct4 and Klf4 were highest in colonies derived from the 2i+Lif+bFGF group. The colonies had round morphology and could be expanded more than 15 passages (Fig. 2C, D). According to the results of reprogramming efficiency, subculturing time, and expression levels of endogenous pluripotent related genes suggest that the 2i+Lif+bFGF culture medium is more suitable for induction and subculturing of piPSCs.
Biological characteristic identification of piPSCs derived from different donor cells
Under 2i+Lif+bFGF medium condition, all PBMSCs, PASCs, and PFFs could be successfully induced into iPSC colonies with high efficiency. The biological characteristic identification results show that piPSCs were positive for AP activity (Fig. 3A); RT-PCR and immunohistochemistry results showed that these colonies expressed the endogenous pluripotent related genes, such as Oct4, Sox2, E-Cadherin (CDH1), Dnmt3b, and GDF3, as well as marker genes FGF5 and Klf4 and Klf5 that contribute to primed and naive-state PSCs, respectively (Fig. 3B, C). However, these piPSCs did not express naive-state marker gene Rex1, and some female piPSCs expressed H3K27me3 with some bar bodies in the nucleus, indicating inactivation of X chromosomes (Fig. 3D).
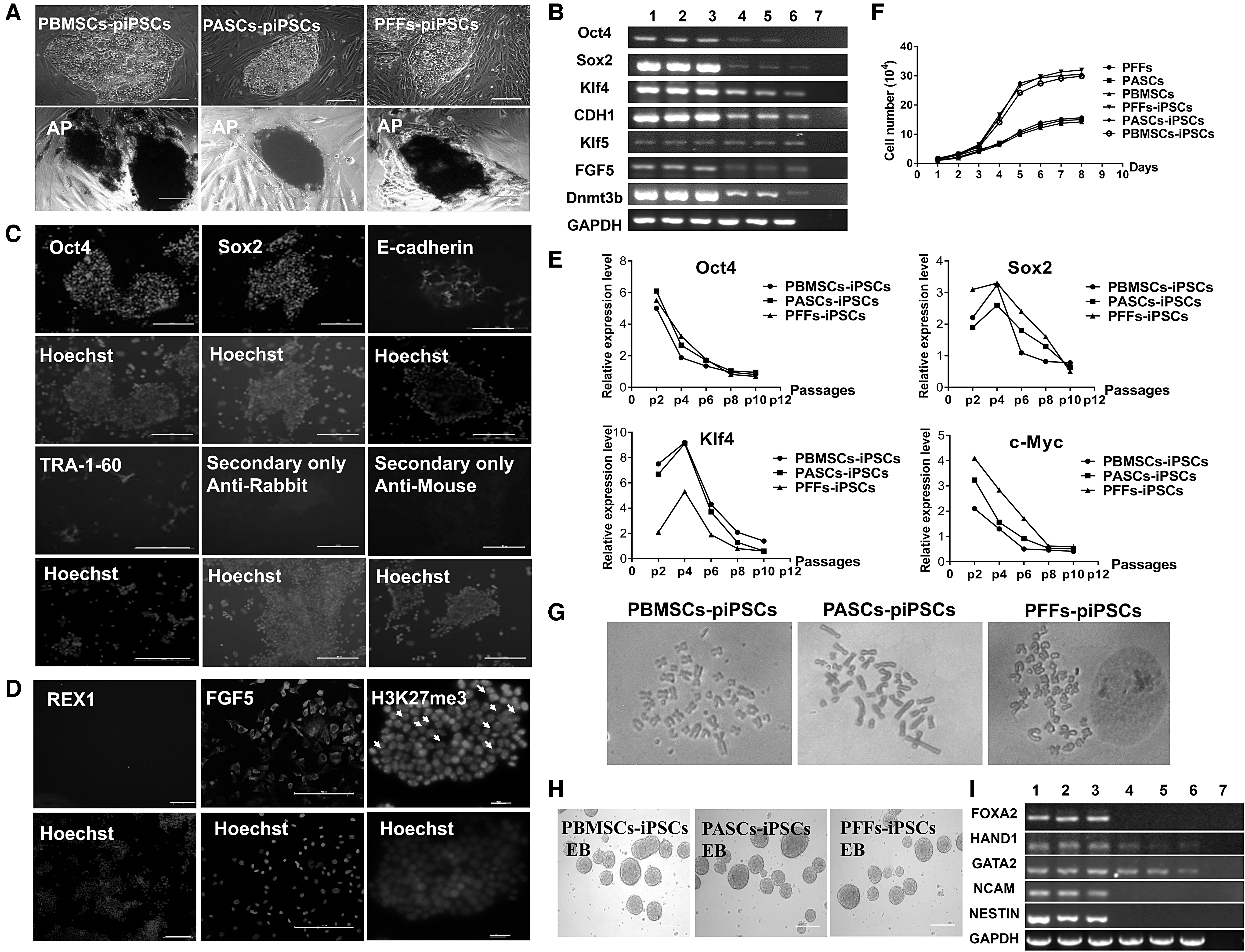
Biological characteristic identification of piPSCs derived from different donor cell induction in bFGF +2i+Lif culture medium (Scale bar = 100 μm).
By withdrawing Dox, piPSCs could still be subcultured and exogenous genes gradually became silent along with passaging (Fig. 3E); piPSCs could proliferate more quickly than their original cells, the population doubling time of piPSCs derived from PBMSCs, PASCs, and PFFs was 0.95, 0.93, and 0.97 days, respectively (Fig. 3F); and piPSCs had 38 chromosomes, the normal karyotype rate was over 70% (Fig. 3G). Furthermore, in vitro differentiation potential test showed that by culturing piPSCs in a low adhesion dish, they could form EBs after 7 days of suspension culture. RT-PCR showed these EBs expressed three germ layer marker genes FOXA2 (endoderm), HAND1 and GATA2 (mesoderm), and NESTIN and NCAM (ectoderm) (Fig. 3H, I).
These results suggest that these piPSCs have the characteristics of PSCs, exhibiting some characteristics of primed and naive-like pluripotent state at the same time.
SB203580 and SP600125 promoted reprogramming of PBMSCs into naive-like state piPSCs
To investigate whether JNK inhibitor SB203580 and P38 inhibitor SP600125 (2s) could promote inducing PBMSCs to naive-like state iPSCs, first, we used cck-8 method to assay different concentrations of SB203580 and SP600125 on the effect of PBMSC proliferation. The results showed that when the concentration of SB203580 and SP600125 exceeded 5 μM, the proliferation of PBMSCs was suppressed significantly (Supplementary Table S3). So 5 μM SB203580 and SP600125 combined with 2i+Lif+bFGF medium were used for subsequent induction and subculture. PBMSCs were chosen as donor cells for induction, and successfully formed piPSC colonies (2s-piPSCs) as we expected (Fig. 4A). 2s-piPSCs could also be expanded more than 15 passages. Biological characteristic identification indicated that the 2s-piPSC colonies exhibited “dome-like” morphology, and were AP staining positive (Fig. 4A). 2s-piPSCs also expressed the naive-state candidate marker genes, such as Rex1, SSEA1, Oct4, Sox2, Klf4, and Klf5, but did not express the primed-state marker gene FGF5 (Fig. 4B, C). QRT-PCR results showed that the expression level of Klf4 derived from 2s treatment group was significantly higher than 2i+Lif+bFGF and bFGF group (Fig. 4D).

Naive-state PSC-related biological characteristics identification of 2s-piPSCs. Scale bar = 100 μm.
In addition, some female piPSCs expressed H3K27me3 with no bar bodies in the nucleus, indicating activation of X chromosomes (Fig. 4C). The methylation status of the Oct4 promoter region was examined in PBMSCs, piPSCs derived from 2i+Lif+bFGF medium, and 2s-piPSCs. The results showed methylation status of the Oct4 promoter region was hypomethylated in piPSCs and 2s-piPSCs, and was lower than in PBMSCs (Fig. 4E). Finally, piPSC colonies could tolerate trypsin digestion, similar to the mouse ESCs, and could form colonies from single cell (Fig. 4F). 2s-iPSCs could also form EBs and three germ layer marker genes FOXA2 (endoderm), HAND1 and GATA2 (mesoderm), and NESTIN and NCAM (ectoderm) expressed in these EBs (Fig. 4G, H).
By comparison with 2i+Lif+bFGF culture medium, some properties of primed-state PSCs disappeared in 2s-piPSCs. These data suggested that 2s-piPSCs have partial properties of naive-state PSCs and that SB203580 and SP600125 can promote piPSC shift to a naive-like pluripotent state.
Discussion
Currently, low reprogramming efficiency and short-term in vitro culture impede the study and application of piPSCs, through selection of different donor cells, or optimizing culture conditions may provide effective mean to solve these problems (Choi et al., 2016; Gao et al., 2014). In this study, we have demonstrated that mesenchymal stem cells PBMSCs and PASCs have higher induction efficiency of piPSCs compared with somatic cells: PFFs. These results were consistent with the previous reports (Tang et al., 2012). Different donor cells have different efficiency of reprogramming into iPSCs. It is generally believed that the efficiency of reprogramming adult stem cells into iPSCs is higher compared with fibroblasts (Tat et al., 2010). Adult stem cells are easy to reprogram mainly due to their specific features.
Adult stem cells express some endogenous pluripotent genes at low level, such as Oct4, Sox2, Nanog, c-Myc, and Klf4, which are also key reprogramming factors (Deng et al., 2019). Single Oct4 could be sufficient for reprogramming of human amniotic fluid stem cells (Qin et al., 2016). Adult stem cells also have the ability of secretion of growth factors, including bFGF, Lif, TGF-β, and activin A, which are always used in promoting proliferation and self-renewal of PSCs (Ahmadzadeh et al., 2020). Recent reports indicated that the pattern of demethylation in pig ADSCs may be another key reason for their easy reprogramming compared with fibroblasts (Zhang et al., 2014).
Dox Tet-on inducible expression lentiviral vector was used in this study. Withdrawing Dox, the interference of the continuous expression of exogenous transcription factors could be eliminated, and is suitable for screening the culture medium of piPSCs. Contrary to previous reports (Liu et al., 2013), we did not establish stable iPSCs in mTeSR culture condition, suggesting that the mTeSR condition may not be suitable for subculturing of piPSCs in our induction conditions. These may be due to the culture mTeSR medium, which is widely used in subculturing of human ESCs and iPSCs, but may not be suitable for the piPSCs. In addition, the long-term cultured piPSCs were induced from lentiviral vector that could continuously express exogenous transcription factors, which maintained the undifferentiated state of piPSCs (Liu et al., 2013; Zhang et al., 2015b).
Although some laboratories have already established piPSCs, culture conditions that support the pluripotency and undifferentiated state of piPSCs in the absence of expression of transgenes are not well known. The results here indicate that the 2i+bFGF+Lif medium is important for piPSC subculturing. Even though both bFGF and 2i+Lif mediums can obtain piPSC colonies, these colonies with low expression level of pluripotent genes in bFGF medium, and proliferation slowly in 2i+Lif medium. bFGF or 2i+Lif medium alone was not suitable for long-term subculture of piPSCs in vitro. However, synergistic bFGF and 2i+Lif could promote proliferation and the expression level of pluripotent genes in piPSCs, and sustain more than 20 passages in vitro.
Many reports stated synergistic action of bFGF and 2i+Lif could support pluripotency and self-renewal of iPSCs (Bourillot et al., 2020). In fact, bFGF and 2i+Lif culture medium were the classical culture medium, which were widely used in primed and naive-state pluripotent stem cells, respectively (Dong et al., 2019). During reprogramming, a different medium may lead the reprogrammed cells to different pluripotent state iPSCs. An intermediate state of piPSCs could be induced by using the combination of Lif, FGF2, BMP4, CHIR99021, and SB431542 (Zhang et al., 2015a). The recent study also generated two types of piPSCs using bFGF and 2i+Lif culture conditions, respectively (Choi et al., 2016). This may be the reason that our piPSCs exhibited some characteristics of primed and naive-like pluripotent state at the same time, when cultured in 2i+Lif+bFGF medium.
Considering naive-state PSCs have the capacity to contribute to chimeric offspring, studies have revealed that naive-state PSCs possess beneficial effect on production of defined filial generation in livestock through chimerism. Exploring the ways to produce naive-state piPSCs is urgently needed. Reports have shown that exogenous cytokines or small molecule inhibitors, which regulate pluripotency or other physiological activities of cell, are an effective way to transform primed-state PSCs into naive state (Altshuler et al., 2018; Honda et al., 2013; Zhang et al., 2014).
However, little is known about naive-state PSCs of livestock, and more small molecule inhibitors need to be screened for naive-state transition. In our study, we added SB203580 and SP600125 into 2i+Lif+bFGF induction medium, and successfully generated naive-like state piPSCs (2s-piPSCs) from PBMSCs. 2s-piPSCs shared characteristics with mouse iPSCs, but not human iPSCs. The results here are similar to previous work on naive-state iPSCs from rhesus monkeys (Fang et al., 2014).
MAPK signaling pathway is downstream of bFGF, including three MAPK families: classical MAPK/ERK, p38 MAPK, and JNK/MAPK, which closely relate to cell proliferation and pluripotency (Liu and Yamashita, 2019; Haghighi et al., 2018). MAPK/ERK inhibitor PD0325901 is an indispensable component of 2i+Lif, a classical mouse ESC culture condition. A combination of p38 MAPK inhibitor SB203580 and JNK/MAPK inhibitor SP600125 enhanced the transition efficiency of naive state (Fang et al., 2014). bFGF was also added in our induction medium. Because many reports indicated that bFGF was indispensable for human ESCs to maintain the naive pluripotent state (Gafni et al., 2013; Ware et al., 2014), bFGF was also important for pig PSCs (Cha et al., 2018; Hou et al., 2016; Telugu et al., 2010).
For naive iPSC identification, our 2s-piPSCs expressed the naive marker genes Rex1, SSEA1, Klf4, and Klf5, but did not express the primed marker gene FGF5, which originally proposed as an epiblast marker. This is possibly due to the use of JNK inhibitor SP600125 and p38 pathway inhibitor SB203580, which stabilized the obtained piPSCs in the naive state. Rex1 and FGF5 are the specific naive and primed markers, respectively, and are widely used as molecular markers to distinguish naive and primed PSCs (Fujishiro et al., 2013; Liu and Yamashita, 2019).
It is generally known that SSEA1 is specifically expressed in naive mouse ESCs, but not primed human ESCs. 2s-piPSCs also exhibited some other characteristics of naive PSCs: “dome-like” morphology, activation of X chromosomes, hypomethylation, colony formation ability by trypsin digestion, and differentiation capability. However, 2s-piPSCs only exhibited partial characteristics of naive PSCs, because without chimerism testing, we called 2s-piPSCs as naive-like piPSCs. Whether 2s-piPSCs belong to naive state remains to be a topic for further study.
Conclusions
In conclusion, we found that PBMSCs as donor cells could improve the efficiency of reprogramming, and the piPSCs could maintain the typical self-renewal morphology on 2i+Lif+bFGF culture condition, even after removal of Dox. SB203580 and SP600125 combination of 2i+Lif+bFGF could reprogram PBMSCs into naive-like state PSCs.
Footnotes
Authors' Contributions
F.C., D.S., and Y.D. conceived the project, analyzed data, and wrote the article; F.C., L.Z., and X.Y. performed most of the experiments; S.Q. and R.Z. performed gene expression assays; Y.D. and S.Y. designed the experiments and supervised the project. All authors have read and agreed to the published version of the article.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This study was funded by the National Natural Science Foundation of China (31760334, 31860644, and 81800525), Guangxi Natural Science Foundation (2018GXNSFAA281007, AA17204051, and 2019GXNSFDA185002), and Project of Guangxi Department of Education 2019KY0048.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.