
Abstract
Mouse embryonic stem cells (mESCs) can maintain self-renewal and differentiate into any cell type of the three primary germ layers. The vascular endothelial growth factor (VEGF) is involved in the regulation of mESC differentiation and induces the activation of a series of kinase responses and several cell signaling pathways by binding to its respective transmembrane receptors, vascular endothelial growth factor receptor VEGFR1, and VEGFR2. Fruquintinib is a selective inhibitor of VEGFRs, and we used it to investigate the effects on the maintenance of pluripotency and differentiation potential of mESCs in this study. Our results showed that fruquintinib-treated cells expressed higher levels of pluripotent markers, including Oct4, Nanog, Sox2, and Esrrb under serum and leukemia inhibitory factor (LIF) condition, whereas the expression of phosphorylated Erk1/2 was restricted. Mitogen-activated protein kinase (MAPK)/extracellular signal-regulated kinase (MEK) signaling inhibitor (PD0325901) and glycogen synthase kinase 3 (GSK3) signaling inhibitor (CHIR99021) (also known as 2i) enable cells to maintain naive pluripotency with LIF, and fruquintinib can also promote cells to maintain naive pluripotent state even under serum/LIF condition, whereas VEGF addition limits the pluripotency characteristics in serum/LIF mESCs. Furthermore, fruquintinib could inhibit the three-germ layer establishment in embryoid body formation and maintain the undifferentiated characteristics of mESCs, indicating that fruquintinib could promote the maintenance of naive pluripotency and inhibit early differentiation programs.
Introduction
Mouse embryonic stem cells (mESCs) are pluripotent cells derived from the inner cell mass (ICM) of mouse blastocysts and have the capacity to differentiate into cell types from the three primary germ layers: ectoderm, endoderm, and mesoderm (Pauklin et al., 2011; Takahashi and Yamanaka, 2006). Naive ESCs exhibit a developmental ground state under the treatment of the leukemia inhibitory factor (LIF) and the kinase inhibitors of glycogen synthase kinase 3 (GSK3) and mitogen-activated protein kinase (MAPK)/extracellular signal-regulated kinase (ERK) (also known as 2i) (Hamilton and Brickman, 2014). mESCs can also be maintained in the presence of serum and LIF (Alexandrova et al., 2016), which generates heterogeneous ESC populations through the self-induced stimulation of the MAPK/ERK pathway (Kunath et al., 2007; Stavridis et al., 2007).
For the early differentiation of mESCs, embryoid body (EB) formation is a process in which cells gradually withdraw from naive pluripotency, and early differentiation marker genes begin to express, which is accompanied by complex signal transduction pathways (Zhou et al., 2010). Hence, signaling pathway transmission is essential for pluripotent maintenance as well as differentiation potential of mESCs.
The small-molecule inhibitors for receptor tyrosine kinase can effectively promote the maintenance of naive pluripotent state (Hanna et al., 2010; Van der Jeught et al., 2013). MAPK/ERK signaling cascades transmit environmental signals of growth factors such as fibroblast growth factors (FGFs), and optimal levels of ERK signaling are essential for self-renewal and pluripotency (Lee et al., 2019; Ma et al., 2016).
Consistent with fibroblast growth factor receptors (FGFRs) and platelet-derived growth factor receptors, vascular endothelial growth factor receptors (VEGFRs) are members of the receptor tyrosine kinase family (Taeger et al., 2011). Similar to FGF signaling transmission, vascular endothelial growth factor (VEGF) signaling activates the MAPK/ERK cascade leading to the induction of a series of kinase responses in the RAS-RAF-MEK-ERK signal axis that complete intracellular signal transmission, which is essential during early embryonic development (Chang et al., 2003; Xu et al., 2008). The FGFR inhibitors or the MEK1/2 inhibitors promote a naive status of mESCs (Williams et al., 2016), however, the effects of VEGFR inhibitors on the maintenance of pluripotency and early differentiation remain to be characterized.
Previous studies have shown that the FMS-like tyrosine kinase (also known as Flt1 or VEGFR1) and kinase domain receptor (also known as Kdr or VEGFR2) are the two critical receptors of VEGF, which are also crucial for the early development of mouse embryos, with a study showing that Flt1−/− mice died on E8.5, E9.0, while Kdr−/− mice died on E8.5, due to a lack of blood vessels (Ji et al., 2018; Wang et al., 2011). Notably, inhibition of VEGF through treatment with a small interfering RNA was shown to lead to a decrease in phosphorylated ERK (pERK) levels (Narasimhan et al., 2009), thereby limiting cell differentiation, indicating that the VEGF signaling transmission is essential for early embryonic development as well as differentiation of ESCs. Taken together, the previous findings suggest that VEGF signaling is essential for cell differentiation and cell growth regulation during embryonic development.
In this study, we provide evidence that fruquintinib, a highly selective and long-term small-molecule inhibitor of VEGFRs (Sun et al., 2014), plays a regulatory role in mESCs' early differentiation. After fruquintinib treatment, early cell differentiation was remarkably suppressed, whereas naive pluripotency-related genes remained highly expressed. Our results indicate that fruquintinib treatment promotes the maintenance of naive pluripotency of mESCs, as well as inhibition of cell differentiation.
Materials and Methods
mESC culture
The mESC line was a gift from Jiahao Sha's laboratory (Nanjing Medical University, China). The cells were cultured at 37°C in a humidified incubator with 5% CO2. mESCs were maintained in feeder-free medium in a four-well plate (JETBIOFIL,TCP-011-004) coated with poly-L-ornithine (P4957; Sigma-Aldrich, Germany) followed by 10 ng/mL laminin (23017015; Invitrogen, USA), and then cultured in serum/LIF or 2i/LIF media. The N2B27 medium was used for the cell cultures, which contains 50% Dulbecco's modified Eagle's medium (10565-018; GIBCO, USA), 50% neurobasal medium (21103-049; GIBCO), 1 × N2 (17502048; GIBCO), 1 × B27 (17504044; GIBCO), and 1 × GlutaMAX (35050061; GIBCO), and is supplemented with 1% penicillin/streptomycin (SV30010; HyClone).
For the 2i/LIF medium, 1 μM of PD0325901 (S1036; Selleck), 3 μM of CHIR99021 (S2924; Selleck), 1000 U/mL LIF (ESG1107; Millipore), 0.1 mM β-mercaptoethanol (M3148; Sigma-Aldrich), and insulin (91077C-250MG; Sigma) were added. For the serum/LIF medium, 15% serum (10100139C; GIBCO), 1000 U/mL LIF, 0.1 mM β-mercaptoethanol, and insulin were added. The concentration of VEGF (96-100-20-10; PeproTech) is 10 ng/mL. The medium was changed every day.
Mouse EB generation
Mouse EBs were derived from mESCs cultured in 2i/LIF medium in vitro. The differentiation medium for EB consisted of Glasgow Minimum Essential Medium (11710035; GIBCO), 10% KSR, 1 × GlutaMAX, 1 × nonessential amino acids (NEAA, 11140050), 1 mM sodium pyruvate (11360070; GIBCO), 1% penicillin/streptomycin, and supplemented with 0.1 mM β-mercaptoethanol. mESCs were disaggregated into a single cell suspension using TrypLE, after which cells were seeded at 3000 cells per well in an ultra-low-attachment 96-well plate (TCP-011-096; JETBIOFIL). The cells were collected on a specified date.
Fruquintinib treatment
Fruquintinib (MCE) was used to evaluate its effects on mESC pluripotency maintenance and early differentiation capacity. The concentration used for experiments was set at 1 μM. mESCs in suspension were treated with or without fruquintinib for 72 hours and then the cells were harvested for analysis. mESCs under 2i/LIF condition were used for the EB induction process, and fruquintinib was added into the EB medium. The solvent dimethyl sulfoxide (DMSO) was used as the control.
Quantitative reverse transcription–polymerase chain reaction
Total RNA was extracted from cells using the TRIzol reagent (15596018; Invitrogen). For each sample, the cDNA was synthesized from 500 ng RNA using the RT Reagent Kit (RR047B; TaKaRa), after which the synthesized cDNA was diluted five times and 1 μL was used per reaction. The ChamQ SYBR QPCR Master Mix (Q341-03; Vazyme) was used to quantify gene expression levels. Relative expression values were normalized to β-actin expression. Each experiment was performed in triplicate. The primers used are provided in Table 1.
Primers Used for Reverse Transcription–Polymerase Chain Reaction in this Study
RT-qPCR, reverse transcription–polymerase chain reaction.
Western blotting
Cells were collected and lysed on ice for 1 hour in a lysis buffer containing 0.5 M EDTA, 1 M Tris-HCl (pH 8.0), 20% sodium dodecyl sulfate (SDS), 5 M NaCl, 10% NP-40, and 12.5% deoxycholate, along with a protease inhibitor cocktail (P5726; Sigma). Cell lysates were centrifuged at 13,000 g at 4°C for 20 minutes. Protein concentrations were quantified using the BCA Protein Quantification Kit (E112-02; Vazyme) according to the manufacturer's instructions. The samples were separated through sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) (PG112; Epizyme) transferred onto 0.45 mm nitrocellulose filter membranes (HATF00010; Millipore) using a wet transfer system. The membrane was then blocked in 5% skim milk diluted in Tris-buffered saline supplemented with 0.1% Tween-20 (TBST) for 1 hour at room temperature.
Afterward, the membrane was incubated with primary antibodies at 4°C overnight. The primary antibodies used were as follows: anti-GAPDH (AP0063, 1:5000; Bioworld, USA), anti-optimal cutting temperature 4 (OCT4) (sc-5279, 1:1000; Santa Cruz, USA), anti-ESRRB (PP-H6705-00, 1:1000; R&D, USA), anti-NANOG (AF1997, 1:2000; R&D), anti-ERK1/2 (4695, 1:1000; Cell Signaling Technology, USA), anti-pERK1/2 (4379, 1:1000; Cell Signaling Technology). The primary antibodies were washed three times with 0.1% TBST buffer for 10 minutes each time, before incubation with secondary antibodies. Immunoreactive protein bands were detected using an ECL reagent (WBKLS0100; Millipore).
Immunofluorescence imaging
For mESC cell slides, cells were cultured on treated clean glass slides. Cells were then washed with phosphate-buffered saline (PBS) and fixed with 4% paraformaldehyde (PFA) at room temperature for 20 minutes. The cells were washed with PBS three times and permeabilized for 20 minutes using PBS supplemented with 0.1% Triton X-100 (PBST). Then, the slides were blocked in 1% BSA at room temperature for 1 hour, after which the cells were incubated with primary antibodies at 4°C for at least 6 hours. The primary antibodies used were as follows: anti-OCT4 (sc-5279, 1:100; Santa Cruz), anti-NANOG (AF1997, 1:100; R&D), FOXA2 (sc-374376, 1:100; Santa Cruz), OTX2 (514195, 1:100; Santa Cruz), EOMES (293481, 1:100; Santa Cruz). The cells were then washed with PBS three times.
Subsequently, the cells were incubated with diluted fluorescence-conjugated secondary antibodies and 4′,6-diamidino-2-phenylindole (DAPI), after which 50% glycerol was used to cover the slides. For EB samples, cells were collected and fixed with 4% PFA. Afterward, the cells were dehydrated using 10% sucrose, embedded in an OCT (4583; SAKURA) compound and cut on a microtome at a 5-μm thickness (CM1950; Leica). Afterward, the same immunofluorescence protocol described above was used to analyze the sections. The image was obtained using a confocal microscope (LSM800; Carl Zeiss), and processed with Zeiss Zen Desk.
Alkaline phosphatase staining assay
The Alkaline Phosphatase Detection Kit (00-0055; Stemgent) was used to determine the differentiation status of ESCs, according to the manufacturer's instructions. The cells were seeded in a four-well plate and alkaline phosphatase (AP) staining was performed when the cells reached a confluency of ∼70%–80%. Briefly, cells were washed with 1 × PBST and then incubated with Fix Solution at room temperature for 2 to 5 minutes. After washing with 1 × PBST, the freshly prepared AP substrate solution was added. Then, the change in color was observed and when the color became bright, the AP substrate solution was immediately removed and the samples were washed with PBS to stop the reaction. Cells were covered with 1 × PBS to prevent drying.
Alkaline phosphatase activity
Colorimetric Assay Kit (K412–500; BioVision) was used to detect the AP activity according to the manufacturer's instructions. ESCs were digested and counted, 1 × 105 cells were washed with PBS, the cells were resuspended in 50 μL assay buffer, and centrifuged at 12,000 g for 2 minutes. The supernatants were mixed with 50 μL of 5 mM pNPP solution and incubated at room temperature for 1 hour in the 96-well plate, and optical density at 405 nm was measured in Synergy H1 (BioTek) after adding 20 μL of stop solution.
Cell cycle analysis using propidium iodide
The cells were fixed with 70%–80% cold ethanol or methanol overnight before staining. Spin-down cell samples at 1000 rpm for 5 minutes, ethanol or methanol solution were removed, then cells were washed with PBS for one time, and resuspended in 150 μL of Propidium Iodide staining solution. All of the cells were then stained. Cells preparing for division would contain increasing amounts of DNA and display proportionally increased fluorescence. Differences in fluorescence intensity are used to determine the percentage of cells in each phase of the cell cycle through FACS (Fluorescence-activated Cell Sorting).
Statistical analyses
GraphPad Prism 7.0 (GraphPad Software, San Diego, CA) was used for all statistical analyses. The expression levels of the genes in each group were compared with the day 3 control group using Student's t-test, two-tailed. The relative quantification of gene expression was performed using the 2−ΔΔCt method, data were presented as the mean ± standard error of mean. A probability (p) value of p < 0.05 was set as statistically significant. ns, no significant difference; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
Results
The addition of VEGF in serum/LIF medium limits the pluripotency characteristics in mESCs
VEGF signaling involves the cascade of signals triggered when the ligand binds the VEGFRs. The downstream signaling includes activation of MAPK/ERK pathway through Ras/Raf activation (Fig. 1A). Compared with the culture condition of 2i/LIF, the expression of Vegfrs was upregulated under the serum/LIF culture medium (Fig. 1B). Therefore, we further explored the effects of VEGF signaling on the pluripotency of mESCs under the condition of serum/LIF. Then, we tested the phosphorylation level of ERK downstream of the VEGF signaling pathway after VEGF treatment. Surprisingly, we found that the level of pERK1/2 was upregulated in response to stimulation with VEGF, which suggested that VEGF/VEGFR and ERK signaling activity could be activated by VEGF (Fig. 1C, D).

Stimulation with VEGF attenuates the pluripotency characteristics of the cells in serum/LIF medium.
In addition, VEGF treatment also reduced the activity of AP in mESCs (Fig. 1E, F). Immunostaining also showed that the expression levels of NANOG and OCT4 would be reduced in VEGF-treated samples, thereby supporting the notion that VEGF could limit mESCs' pluripotent state under serum/LIF condition (Fig. 1G, H). Moreover, the expression of naive pluripotent state-related genes Oct4, Nanog, and Esrrb were downregulated in VEGF-treated cells, except for Sox2 (Fig. 1I). Taken together, the results indicated that the stimulation with VEGF could further reduce the pluripotency of cells in serum/LIF medium with regard to restriction of AP reactivities and stable expressions of pluripotent markers. Therefore, VEGF impairs the maintenance of pluripotency in serum/LIF-cultured mESCs.
The naive pluripotency stabilities are comparable in 2i/LIF ESCs and serum/LIF ESCs with fruquintinib stimulation
To further explore the influences of VEGF signal on mESCs' pluripotency maintenance under the serum/LIF conditions, the effects of VEGFR restriction on mESCs were detected using fruquintinib, a highly selective small-molecule inhibitor of VEGFRs and we found that fruquintinib can significantly reduce the phosphorylation level of ERK (Fig. 2A, B).
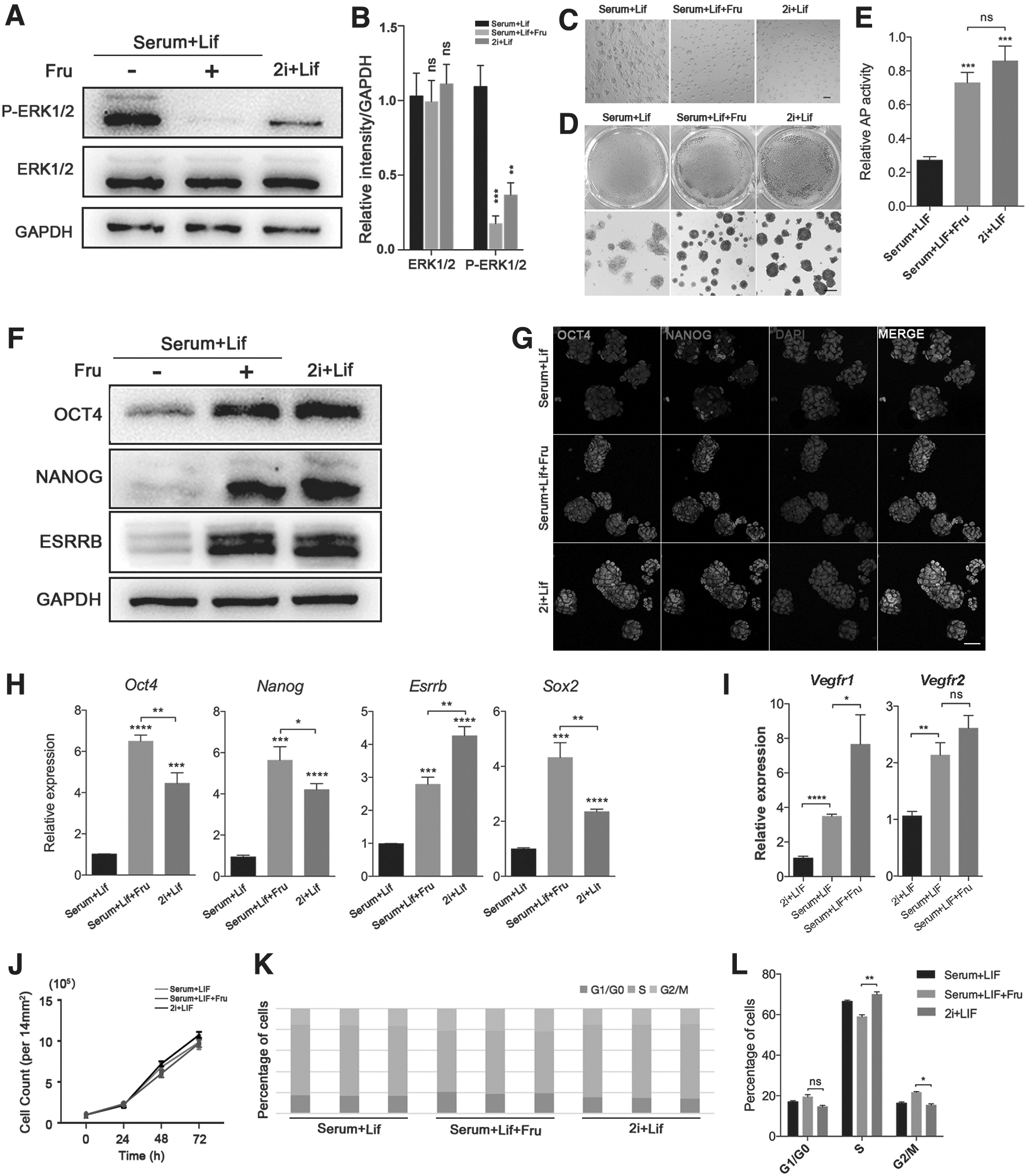
Fruquintinib treatment can maintain the naive pluripotency stability in serum/LIF-cultured mESCs similar with 2i/LIF-cultured cells.
To further study the effects of fruquintinib on the maintenance of the naive pluripotent state of mESCs, we compared mESCs cultured in the presence of 2i/LIF with mESCs cultured in the presence of serum/LIF with fruquintinib. Additionally, the fruquintinib treatment could induce serum/LIF mESCs to maintain round colony morphology like mESCs cultured in 2i/LIF medium, whereas the mESCs cultured in serum/LIF medium grew into flat and compact colonies (Fig. 2C). AP staining and analysis showed that mESCs treated with fruquintinib under the serum/LIF condition and mESCs cultured in 2i/LIF medium were comparable and both of them had the positive AP reactivities, indicating that fruquintinib helps maintain the naive pluripotency of mESCs (Fig. 2D, E).
Then, the changes on protein and mRNA levels of pluripotency markers were observed. Our results showed that the expressions of pluripotency-related factors were also comparable in 2i/LIF mESCs and serum/LIF mESCs with fruquintinib stimulation (Fig. 2F). Oct4 and Nanog are essential regulators for naive pluripotency maintenance and also regulate the maintenance of ICM during preimplantation development (Loh et al., 2006). Notably, we showed that 2i/LIF mESCs and fruquintinib-treated cells maintained similar high expression levels of OCT4 and NANOG through immunofluorescence staining, indicating that the cells remained in a naive pluripotent state under fruquintinib stimulation (Fig. 2G).
Additionally, the mRNA expression levels of the naive pluripotent genes Oct4, Nanog, and Sox2 in serum/LIF/fruquintinib mESCs were even higher than their expressions in cells cultured under 2i/LIF conditions (Fig. 2H). Although fruquintinib can reduce the phosphorylation level of ERK downstream of VEGF/VEGFR and promote the maintenance of the naive pluripotency of mESCs, the expression level of Vegfr2 did not change significantly and the expression of Vegfr1 compensatory increased slightly under the treatment of fruquintinib in serum/LIF medium, fruquintinib cannot inhibit the mRNA expression levels of Vegfr1/Vegfr2 (Fig. 2I).
The assays of cell proliferation and cell cycle showed that the 2i/LIF mESCs proliferation rate was the fastest, and the serum/LIF and serum/LIF/Fru mESCs also proliferate at a comparable rate (Fig. 2J). Compared with the 2i/LIF ESCs, the percentage of cells in S phase is reduced in serum/LIF/Fru cells, but there is no cell cycle arrest in G1 phase (Fig. 2K, 2L). These results suggesting that similar to mESCs cultured under 2i/LIF conditions, fruquintinib can also promote the maintenance of naive pluripotency in mESCs cultured in serum/LIF medium.
Fruquintinib treatment promotes the maintenance of pluripotent characteristics in mESCs during the EB formation process
mESCs are pluripotent and can therefore differentiate into various types of cells under different induction conditions. EBs are three-dimensional aggregates, which can be formed in suspension by ESCs and induced pluripotent stem cells (Sakai et al., 2011). EBs consist of representatives of all three germ layers formed from mESCs with spontaneous differentiation capacity. EB differentiation is a common platform for the generation of specific cell lineages from PSCs (Dang et al., 2002). During the process of EB differentiation, the expression levels of pluripotent genes decreased, whereas the expression of genes associated with germ layer formation gradually increased (Mansergh et al., 2009).
First, an in vitro differentiation system to form EBs was used, and the naive mESCs cultured under 2i/LIF conditions were induced to differentiate into three germ layers. We found that during the differentiation of naive mESCs into EBs, the expression of Vegfrs were increased gradually under normal culture conditions, suggesting that VEGF signaling may play an important role in the differentiation process of EBs (Fig. 3A). Next, naive mESCs were digested and counted, then resuspended in EB differentiation medium and seeded in 96-well plates. Fruquintinib or 2i was used in the EB medium and the effects on the exit of the naive state had been detected, the solvent DMSO was added as a control (Fig. 3B).
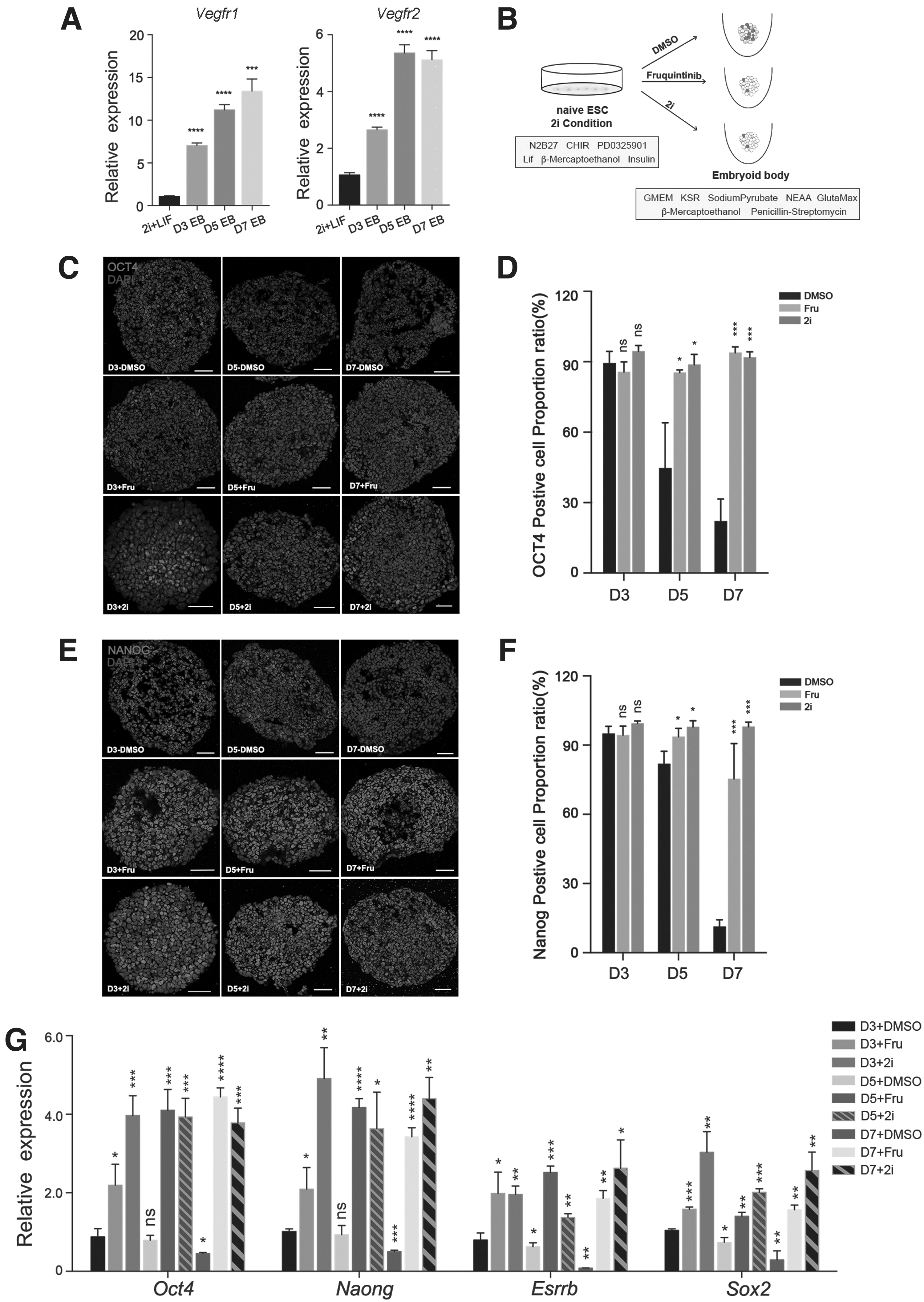
Fruquintinib promotes the maintenance of pluripotent characteristics during the EB differentiation process.
We quantified the expressions of pluripotency markers by immunostaining analysis at the indicated time points of differentiation. It is indicated that stimulation with fruquintinib or 2i promotes the maintenance of naive pluripotency due to the homogeneous and high expression of OCT4 and NANOG (Fig. 3C–F). As expected, the expression of naive pluripotent genes was shown to be gradually downregulated in DMSO-treated cell aggregates, whereas fruquintinib or 2i-treated cells were shown to maintain high gene expression levels (Fig. 3G). Overall, our results showed that fruquintinib limits the exit of naive pluripotent state under EB differentiation conditions.
Fruquintinib impairs mESCs' early differentiation
In addition to maintaining self-renewal, mESCs can differentiate into the three germ layer cells (Sladitschek and Neveu, 2019). To study the potential influence of fruquintinib on the fate determination of mESCs during early differentiation, mEBs were formed in vitro, after which the expression levels of lineage-specific markers were quantified through quantitative reverse transcription–polymerase chain reaction (RT-qPCR) and immunostaining analysis. mEBs were harvested for analysis on day 3, 5, and 7. Our results showed that expression of endoderm markers, such as FOXA2; mesoderm markers, such as EOMES; and neuroectoderm markers, such as OTX2; could be detected in day 5 and 7 EBs, whereas these lineage-specific markers failed to be detected in EBs treated with fruquintinib or 2i (Fig. 4A–F).
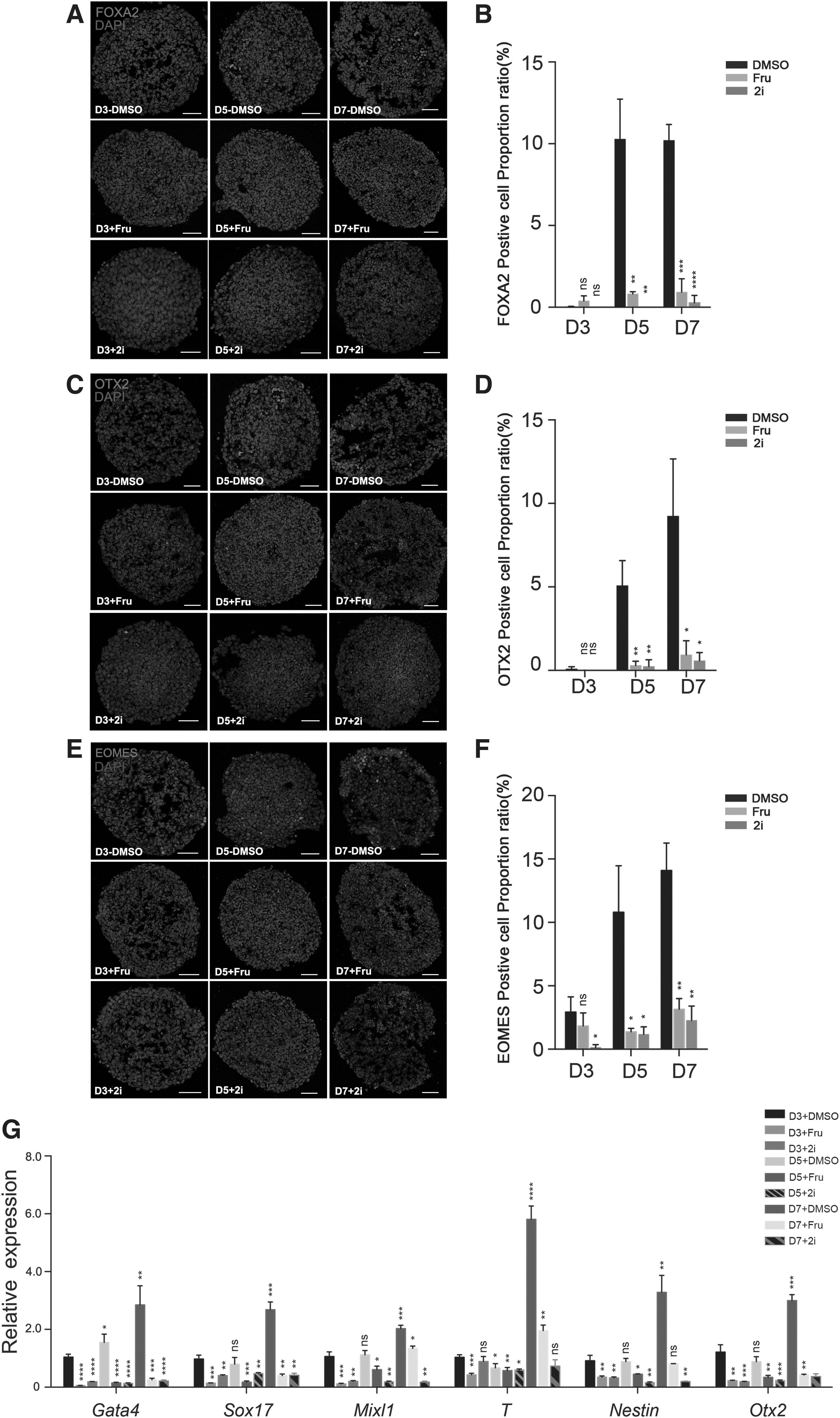
Cells cannot successfully enter the early differentiation process with the fruquintinib treatment.
Additionally, the RT-qPCR analysis confirmed that the expression of endoderm markers (Gata4 and Sox17), mesoderm markers (Mixl1 and T), and ectoderm markers (Nestin and Otx2) was significantly reduced in fruquintinib- or 2i-treated EBs (Fig. 4G). On the other hand, the naive pluripotency markers Oct4, Nanog, Esrrb, and Sox2 maintained a high level of expression under fruquintinib or 2i stimulation (Fig. 3G). Consistent with previous studies, which reported that inhibiting VEGFR activity results in abnormal cell differentiation and fetal death during embryonic development, our results also suggested that fruquintinib impairs early differentiation of mESCs during EB formation and keeps cell aggregates in a naive pluripotent state.
Discussion
Pluripotency, self-renewal, and continuous proliferation are the most salient features of mESCs. A transcription factor network and a series of specific signaling pathways work together to maintain this state (Ng and Surani, 2011). Initial findings suggest that mESCs can be cultured in the presence of serum on feeder cells and maintain the cells in a pluripotent state. However, recent studies have found that the combination of the key signaling factors, LIF and bone morphogenetic protein 4, can also help promote the maintenance of naive pluripotency and self-renewal capacity in the absence of feeder cells (Chambers, 2004; Evans and Kaufman, 1981; Niwa et al., 2009).
Furthermore, the naive pluripotency of ESCs can be maintained in minimal medium with MEK and GSK3 inhibitors (2i) (Ye et al., 2014). LIF is often used as a supplement in this kind of culture system, which not only supports the self-renewal capacity of ESCs but also contributes to extraembryonic development (Davey et al., 2007; Yi et al., 2020). There are also some small-molecule inhibitors of receptor tyrosine kinases that are also effective in promoting the maintenance of a naive pluripotent state. These inhibitors can bind to the active site of a RTK, thus preventing phosphorylation and by doing so regulate or modulate signaling.
VEGFRs are members of the receptor tyrosine kinase family and VEGF family members bind to their receptors (VEGFRs) to regulate angiogenesis, which is essential for early embryonic growth and development (Boer et al., 2008; Ferrara and Gerber, 2001). Fruquintinib is an orally effective, efficient, and highly selective inhibitor of VEGFR-1, -2, and -3. It also acts as a novel anti-VEGFR tyrosine kinase inhibition. Administration of VEGF leads to the activation of various cell signaling pathways, including the MAPK/ERK signaling pathway, which results in increased levels of phospho-Erk1/2 and regulates cell proliferation, differentiation, and adaptation (Takahashi et al., 1999; Tan et al., 2013). Therefore, the role of fruquintinib in the early differentiation of mESCs is of great interest to us.
Recent studies have shown that signaling pathways are crucial to ESC differentiation both in vitro and in vivo (Sun et al., 2015). When the MAPK/ERK signaling pathway in ESCs is blocked, the cell fate will no longer move toward the mesoderm or neuroectoderm (Hayashi and Ogura, 2020). Our study confirms that adding 10 ng/mL VEGF to activate the VEGF/VEGFR signaling pathway will increase the phosphorylation level of the downstream ERK, and at the same time weaken the pluripotency in the serum/LIF-cultured mESCs. On the contrary, blocking the VEGF signaling pathway through fruquintinib administration leads to reduced levels of phospho-Erk1/2, thereby limiting cell differentiation and further affecting cell fate decisions.
Feed-forward loops are formed by Oct4, Nanog, Esrrb, and Sox2 to maintain the pluripotency of mESCs (Heng and Ng, 2010; Xue et al., 2011; Yeo and Ng, 2013). The four master modulators of pluripotency control the destiny of ESCs and regulate the induction of developmental programs. In this study, we found that the expression of Oct4, Nanog, Esrrb, and Sox2 were comparable in serum/LIF-cultured mESCs treated with fruquintinib and 2i/LIF-cultured mESCs.
The effects of fruquintinib on mEBs differentiation were also investigated and the results indicated that when the naive mESCs were switched to the EB medium that induces differentiation, fruquintinib limits the exit of naive pluripotency and the expression of different germ layer markers in mEBs were significantly decreased compared with the cells under normal culture conditions.
Altogether, these results showed that fruquintinib plays a regulatory role in maintaining mESC pluripotency and restricting early differentiation. mEBs containing three germ layers can be formed when 2i is withdrawn. The existence of 2i will help to establish a naive ground state, and fruquintinib can also maintain the pluripotency state and hinder cell differentiation, which is similar to 2i. The results in our study improve our understanding of the effects of fruquintinib on the cell pluripotent state of mESCs.
Cellular morphology and homogeneity of pluripotency-associated gene expression differed between the 2i/LIF and serum/LIF conditions. 2i/LIF ESCs were morphologically uniform and homogeneously expressed pluripotency-associated genes, whereas serum/LIF ESCs were heterogeneous for both. Interestingly, when the growth medium conditions were switched, ESCs changed to take on the morphological characteristics of the new condition and previously observed expression differences also switched suggesting that the cell states exhibited in 2i/LIF and serum/LIF conditions are interconvertible and are dependent on their growth conditions.
mESCs grown in serum exhibit greater heterogeneity in morphology and expression of pluripotency factors than ESCs cultured in defined medium with 2i postulated to establish a naive ground state. We have proved that fruquintinib can promote the maintenance of the naive pluripotency of mESCs under serum/LIF conditions, and its effects on the differentiation potential of mESCs is also worthy of further study. Both mESCs cultured under serum/LIF and 2i/LIF conditions have pluripotency, but their differentiation potential is different. The pluripotency characteristics of mESCs under the conditions of serum/LIF/fruquintinib and 2i/LIF were found to be comparable. While the ability to differentiation into three germ layers of the cells in the similar pluripotency state obtained by these two culture methods is also worthy of further investigation.
Pluripotency is maintained by a synergy between external stimulation and internal circuits that allow the maintenance of undifferentiated and self-renewal states. In this study, we confirmed that fruquintinib treatment restricts the early differentiation of mESCs, specifically, on the reduction of the expression levels of pERK1/2 and the three germ layer markers under the differentiation conditions. Studies on small-molecule inhibitors of receptor tyrosine kinases help us to understand the molecular mechanisms of pluripotency maintenance and differentiation potential of ESCs, which are also important for developmental biology and regenerative medicine applications.
Conclusion
In this study, we demonstrated that fruquintinib helps to maintain the naive pluripotency stabilities in serum/LIF medium. In addition, treatment with fruquintinib specifically restricted the expression of endodermal, mesodermal, and ectodermal markers and promoted the expression of pluripotent marker genes in mEBs. These results help us better understand the effects of VEGF signal transmission on mESC pluripotency maintenance and differentiation potential. Taken together, these results indicate that fruquintinib treatment could be easily and widely applied to maintain pluripotency and regulate the pluripotent state of mESCs similar to 2i.
Footnotes
Author Disclosure Statement
The authors declare there are no conflicting financial interests.
Funding Information
This research is funded by the National Key R&D Program of China [No. 2016YFA0503300, 2017YFC1001302], and the National Natural Science Foundation of China [No.31871445, 31971330].