
Abstract
Transgenic goats are ideal bioreactors for the production of therapeutic proteins in their mammary glands. However, random integration of the transgene within-host genome often culminates in unstable expression and unpredictable phenotypes. Targeting desired genes to a safe locus in the goat genome using advanced targeted genome-editing tools, such as transcription activator-like effector nucleases (TALENs) might assist in overcoming these hurdles. We identified Rosa 26 locus, a safe harbor for transgene integration, on chromosome 22 in the goat genome for the first time. We further demonstrate that TALEN-mediated targeting of GFP gene cassette at Rosa 26 locus exhibited stable and ubiquitous expression of GFP gene in goat fetal fibroblasts (GFFs) and after that, transgenic cloned embryos generated by handmade cloning (HMC). The transfection of GFFs by the TALEN pair resulted in 13.30% indel frequency at the target site. Upon cotransfection with TALEN and donor vectors, four correctly targeted cell colonies were obtained and all of them showed monoallelic gene insertions. The blastocyst rate for transgenic cloned embryos (3.92% ± 1.12%) was significantly (p < 0.05) lower than cloned embryos (7.84% ± 0.68%) used as control. Concomitantly, 2 out of 15 embryos of morulae and blastocyst stage (13.30%) exhibited site-specific integration. In conclusion, the present study demonstrates TALEN-mediated transgene integration at Rosa 26 locus in caprine fetal fibroblasts and the generation of transgenic cloned embryos using HMC.
Introduction
The generation of transgenic farm animals is crucial due to their potential application as bioreactors for the cost-effective production of pharmaceutical/therapeutic proteins in their mammary gland (Mehta et al., 2017). Additionally, these animals have tremendous potential to serve as models for biomedical research and organ and tissue donors for xenotransplantation in humans. In this regard, goats are ideal candidates as bioreactors due to the high yield of purified product, short generation interval, and generation of multiple offsprings (Fernandez and Hoeffler, 1998). For instance, the recently commercialized recombinant human antithrombin III expressed in transgenic goats' milk (Shepelev et al., 2018) serves as the most convincing evidence regarding potential applications of transgenic livestock animals for human welfare.
For the past several years, researchers have made numerous efforts to generate transgenic farm animals relying upon the random insertion of the transgene within the host genome, although it resulted in its low, unstable, and unpredictable expression (Kalds et al., 2019; Lantinga-van Leeuwen et al., 2006; Papapetrou and Schambach, 2016; Smithies, 2001). However, in recent advancements, the development of gene-targeting technologies encompassing site-specific nucleases viz. zinc-finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and clustered regularly interspaced short palindromic repeats (CRISPR)/Cas has played a pivotal role to overcome these barriers. These advanced methods stimulate the homologous recombination by several folds and targeting a single copy of transgene at a specified locus resulting in its highly stable and constitutive expression.
Among these novel transgenic technologies, TALENs have proven to be highly efficient genome-editing tools due to their simple and easy designing, high specificity, low cytotoxicity, and rare off-target activities (Li et al., 2020). It has been rapidly and robustly used for precise genome editing in various cell types across the species (Carlson et al., 2016; Zhang et al., 2019). TALENs consist of TALE DNA-binding domain fused with a nonspecific FokI endonuclease, which introduces a double-strand break at the target site within the genome, which is then repaired by either nonhomologous end joining, resulting in gain or loss of few base pairs leading to the gene knockout or homology-directed repair if a transgene is provided, leading to its insertion/knockin at the specific locus (Boch and Bonas, 2010).
In addition to the genome-editing tool, selecting a safe genomic site with high homologous recombination frequency and ubiquitous transcriptional activity is crucial for a successful gene-targeting study. Rosa 26 locus has remained the most preferred docking site for transgene integration in the mouse model (Friedrich and Soriano, 1991; Imayoshi et al., 2020; Zambrowicz et al., 1997). The insertion of the transgene into Rosa 26 locus allows site-specific integration of a single copy of transgene resulting in its stable and optimized expression without causing any aberrant physiological consequences (Dulauroy et al., 2012; Sadelain et al., 2012). Rosa 26 locus has also been identified and successfully targeted in rat (Kobayashi et al., 2012), rabbit (Yang et al., 2016), pig (Li et al., 2014; Xie et al., 2017), sheep (Wu et al., 2016), cattle (Wang et al., 2018), and human (Irion et al., 2007).
However, to the best of our knowledge, there is no report available on identifying and targeting Rosa 26 loci in the goat genome. Identifying Rosa 26 locus in the caprine genome is a valuable asset to the toolbox of transgenic goat research and would improve the efficiency of targeted transgene integration to generate transgenic goats. Somatic cell nuclear transfer (SCNT) is an efficient technique for generating transgenic farm animals and has been preferred over pronuclear microinjection for this purpose (Srirattana et al., 2013; Zhou et al., 2015). The use of transgenic cells harboring the desired gene as nuclear donors for SCNT ensures the transgene's presence and expression in resultant offspring (Bressan et al., 2013). Moreover, one founder animal's production this way provides an opportunity to generate a herd of transgenic animals through germline transmissions (Whitelaw et al., 2016).
With this backdrop, the present study was undertaken to identify the Rosa 26 locus in the goat genome and optimize the site-directed introduction of GFP at Rosa 26 locus in fetal fibroblasts using TALENs, followed by their use as a nuclear donor for producing transgenic handmade cloned goat embryos.
Materials and Methods
The present study was carried out at the Embryo Biotechnology Laboratory, Animal Biotechnology Center, ICAR-National Dairy Research Institute, Karnal, India. All the experiments were duly approved and performed following relevant guidelines and regulations by the Institute Bio-Safety Committee (IBSC/NDRI/Dt.29.04.2014). Goat ovaries and uterus were collected from a local abattoir following the guidelines by the Institute Animal Ethics Committee. All the chemicals and media for cell and embryo culture were procured from Sigma Chemical (St Louis, MO) unless otherwise specified. The molecular biology reagents were obtained from Thermo Fisher Scientific (Waltham, MA, USA). The fetal bovine serum (FBS) was procured from HyClone (South Logan, UT) and plastic ware from Nunc (Roskilde, Denmark).
Identification, cloning, and sequencing of goat Rosa 26 locus
The porcine Rosa 26 sequences (>gi|347615106|ref|NW_003611693.1| Susscrofa breed mixed chromosome 13 genomic scaffold, Sscrofa10.2) were used as a template to search the goat genome (www.goatgenome.org/projects.html). Using bioinformatics tools, a highly conserved sequence located on caprine chromosome 22 was isolated as putative Rosa 26 locus. Putative goat Rosa 26 loci were also aligned with sheep Rosa 26 sequences for further confirmation (Wu et al., 2016). A 2.2 kb fragment of Rosa 26 locus was amplified from caprine genomic DNA isolated from uterine tissue using the following polymerase chain reaction (PCR) program: initial denaturation at 95°C for 3 minutes; followed by 35 cycles at 95°C for 30 seconds, 58°C for 30 seconds, and 72°C for 2.2 minutes; with a final extension at 72°C for 10 minutes. The primer sequences used for amplification of goat Rosa 26 locus are given in Table 1.
Polymerase Chain Reaction Primers for Goat Rosa 26 Amplification and Targeting Vector Construction
Bold and underscored denotes sequence of restriction enzyme sites (in parentheses) added in primers.
KIGS, Kan-IRES-GFP-SV40pA.
The PCR products were observed under ultraviolet transilluminator, excised, and purified from the gel using the GeneJET Gel Extraction Kit (Thermo Fisher Scientific). The purified PCR product was ligated into a TA cloning vector (InsTAclone PCR Cloning Kit; Thermo Fisher Scientific) and cloned into Escherichia coli XL1-Blue MRF' host and sequenced from Merck Specialities Pvt. Ltd. The sequence obtained was verified by BLAST and Pairwise alignment in Bio-Edit.
Generation of a TALEN pair and Rosa 26 targeting vector P5′KC3′
Left (L) and Right (R) TALEN vectors (Fig. 1a) targeting Rosa 26 intron 1 region were designed and constructed from Life Technologies Corporation (Carlsbad, CA). The TALEN vectors contained TALE proteins specific for binding TAL L and TAL R sequences of Rosa 26 intron 1 region, fused with nonspecific FokI endonuclease. RVD (NN-NN-NNHD-NN-NG-NG-NN-NG-HD-HD-NG-NN-HD-NI-NN-NG-NN) was designed for 18 bp left TAL-binding sequence NGGGCGTTGTCCTGCAGTG, whereas RVD NN-NN-NNNG-NG-HD-NI-NG-HD-HD-NG-HD-NG-NI-NI-NI-NG-HD for 18 bp right TAL-binding sequence NGGGTTCATCCTCTAAATC. These left and right TALEN-binding sites were separated by a 16 bp spacer sequence GATCGAGCAGGTATAA.

The Rosa 26 targeting vector/donor vector P5′KC3′ (pIRES 5′ KIGS CMV 3′) was constructed by using the pIRES2-AcGFP1 (Clontech) vector as a backbone (Fig. 1b). The fragments 5′ goat Rosa 26 homology arm, Kan-IRES-GFP-SV40pA fragment, CMV promoter, and 3′ goat Rosa 26 homology arm were sequentially added into backbone vector by restriction enzyme digestion and ligation using T4 DNA ligase. The Rosa 26 homology arms 5′ (864 bp) and 3′ (1283 bp) were amplified from the goat genome and cloned into a TA cloning vector. The Rosa 26 5′ homology arm was inserted into the pIRES2-AcGFP1 (5.3 kb) vector using NheI and AseI restriction enzymes replacing CMV promoter. Kanamycin resistance gene was inserted into multiple cloning sites (MCS) of the pIRES2-AcGFP1 vector using EcoRI and BamHI, and Kan-IRES-GFP-SV40pA fragment was generated from this vector using PCR.
Kan-IRES-GFP-SV40pA (KIGS) fragment (2.25 kb) was inserted into MCS of pIRES 5′ arm vector by EcoRI and SalI restriction digestion and ligation. CMV fragment (600 bp) was amplified from pIRES2-AcGFP1 mammalian expression vector using primers having NheI and EcoRI sites into 5′ region of forward and reverse primer, respectively, and modified AseI site in forward primer and inserted into MCS of pIRES 5′ KIGS by using NheI and EcoRI enzymes. Subsequently, a 3′ Rosa 26 homology arm was inserted into MCS of pIRES 5′ KIGS CMV vector with SalI and BspTI restriction enzyme digestion and ligation. Finally, the Rosa 26 targeting vector P5′KC3′ (8.0 kb) was verified by sequencing inserts from both the ends. The primer sequences used are given in Table 1.
Establishment and characterization of goat fetal fibroblast cells
Goat fetal fibroblasts (GFFs) were established from ear skin tissues collected from a 50-day-old fetus as described previously (Shah et al., 2009). The sex of the established GFFs was confirmed by PCR amplification of SRY and PLP genes using genomic DNA. The fetal fibroblast cultures were characterized by immunocytochemistry for the expression of fibroblast cell-specific (β-Tubulin, Vimentin) as well as nonspecific markers (Keratin 78, Cytokeratin 18) as described previously (Selokar et al., 2012). The cells were further characterized by reverse transcription/polymerase chain reaction (RT-PCR), for which RNA was isolated from GFFs using TRIzol (Invitrogen), and complementary DNA (cDNA) was synthesized using the Superscript III First-Strand Synthesis Kit (Invitrogen).
The expression of cell-specific markers, namely, VIMENTIN, TUBULIN, CYTOKERATIN-8, and CYTOKERATIN-18 was analyzed by RT-PCR under the following cycling conditions: initial denaturation at 94°C for 3 minutes; followed by 94°C for 30 seconds, X°C (annealing temperature as mentioned for each primer pair in Table 2) for 30 seconds, and 72°C for 30 seconds for 35 cycles; followed by a final extension at 72°C for 10 minutes.
Primers for Fibroblast Cell Characterization
Evaluation of TALEN efficiency
To determine TALEN vectors' efficiency to bind at Rosa 26 locus and to introduce a double-strand break, GFFs were transfected with left and right TALEN vectors by nucleofection as described previously (Sharma et al., 2020). Briefly, GFFs at passage 3 were grown to 70%–80% confluency, trypsinized, and 0.5 × 106 cells were resuspended in 82 μL P3 solution and 18 μL supplement 1 solution (Amaxa). Left and right TALEN vectors (1.5 μg each) were added to the resuspended cells and nucleoporated in 1 cm transfection cuvettes using EN-150 program in 4 D nucleofector (Amaxa). The transfected cells were seeded in a six-well plate containing Dulbecco's modified Eagle's medium (DMEM) +10% FBS and cultured at 37°C in a 5% CO2 incubator.
After 72 hours, the genomic DNA was isolated from the transfected cells, and the fragment spanning the target site was amplified using primers TAL F and TAL R following PCR cycling conditions: initial denaturation at 94°C for 3 minutes; followed by 94°C for 30 seconds, 52°C for 30 seconds, and 72°C for 30 seconds for 33 cycles; and a final extension at 72°C for 10 minutes. The amplified fragment was purified and cloned into E. coli XL1-Blue MRF' host. After confirmation of the recombinant clones for insert sequence based on PCR amplification, 30 clones were sent for custom sequencing from Merck Specialities Pvt. Ltd. The sequence obtained was verified by BLAST and Pairwise alignment in BioEdit to detect indels.
TALEN-mediated targeting of circular donor vector at Rosa 26 locus
GFFs were cotransfected with left and right TALEN vectors (1.5 μg each) and circular donor vector (2.5 μg) by nucleofection as mentioned earlier, and after 72 hours, G418 (700 μg/mL) was added to select the transfected cells. G418-resistant cell colonies expressing GFP were marked by visualizing under the fluorescence microscope, and cloning rings (Sigma, St. Louis, MO) were loaded over the marked colonies. The cells were then trypsinized and seeded at low density in 96-well cell culture plates and further propagated for cryopreservation and DNA isolation for detecting mutations.
TALEN-mediated targeting of linear donor vector
Linear donor vector fragment from 5′ to 3′ arm was amplified using circular donor vector (P5′KC3′) as the template by Phusion High-Fidelity DNA Polymerase (Thermo Fisher Scientific) to get a blunt-end vector using PCR cycling conditions: initial denaturation at 98°C for 1 minute, followed by 35 cycles of 98°C for 10 seconds, 60°C for 30 seconds, 72°C for 2.5 minutes, and a final extension at 72°C for 10 minutes. The PCR product was purified by the Gene JET PCR Purification Kit (Thermo Fisher Scientific). The GFFs were cotransfected by left and right TALEN vectors (1.5 μg each) and linear donor vector (2.5 μg) by nucleofection, selected using G418, and seeded in 96-well cell culture plates for cryopreservation or DNA isolation as mentioned above.
Screening of correctly targeted cell colonies
To detect targeted integration of donor vector at Rosa 26 locus, DNA was isolated from the cell colonies picked using cloning rings and used as a template to perform 5′ and 3′ junction PCR. PCR analysis was performed with primers GR5′END F/CMV-R3 (5′ junction, 1679 bp product) and primers SV40 F/GR3′END R (3′ junction, 1857 bp product). Primers for 5′ and 3′ junction PCR (Table 3) were designed such that only correctly targeted cell colony gave the desired band, as shown in Figure 2a. The PCR cycling conditions were: 94°C for 3 minutes, followed by a cycling program of 94°C for 30 seconds, 54°C for 30 seconds, and 72°C for 1.5 minutes for 35 cycles. The reactions were ended with a final extension at 72°C for 10 minutes.
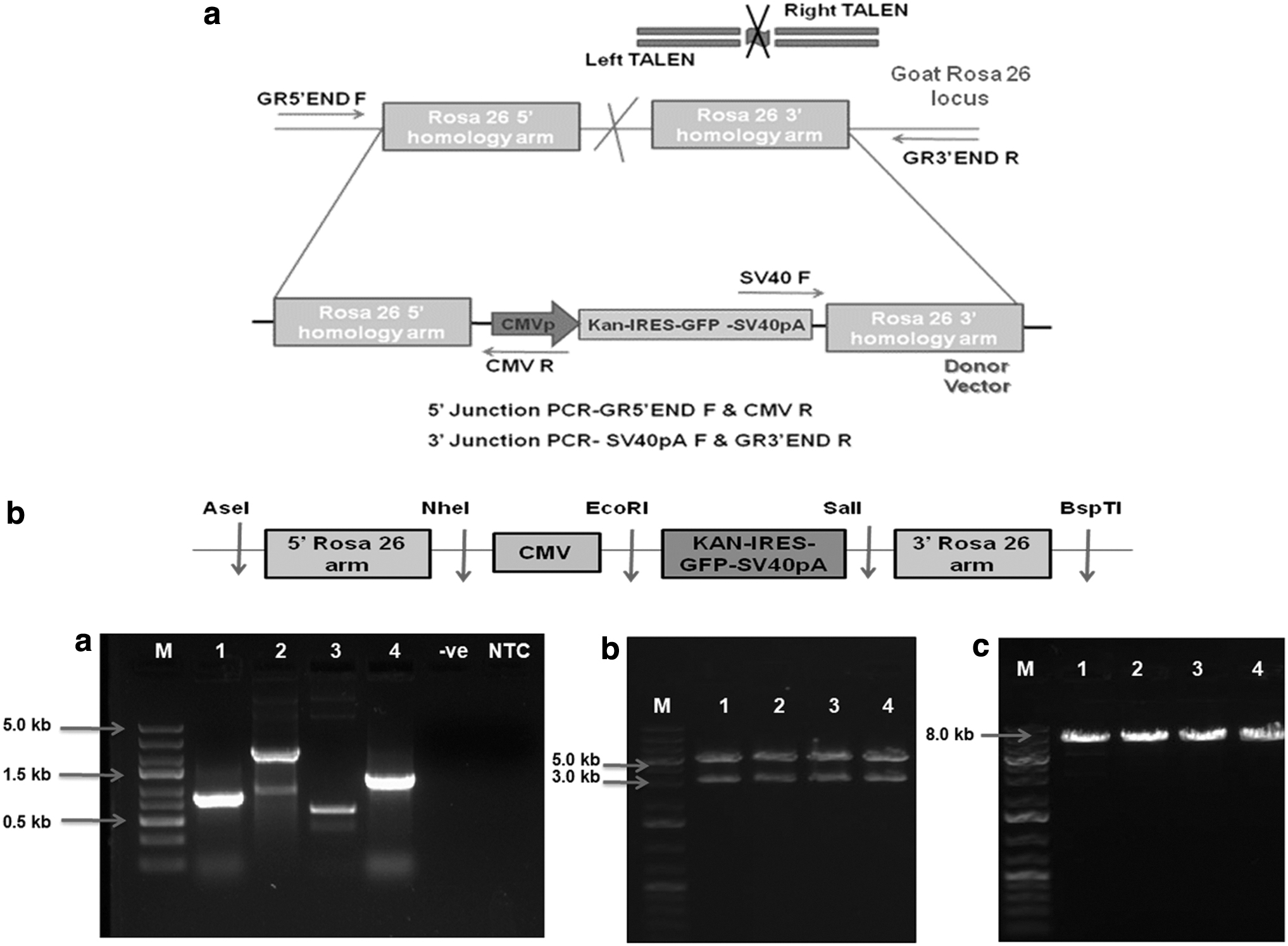
Primers for 5′ and 3′ Junction Polymerase Chain Reaction
PCR, polymerase chain reaction.
Screening targeted cell colonies for mono and biallelic gene insertions
DNA from the correctly targeted cell colonies was isolated and used as the template for the amplification of fragment spanning the target site using specific primers for screening biallelic variants. The PCR was performed as follows: initial denaturation at 94°C for 3 minutes followed by a cycling program of 94°C for 30 seconds, 52°C for 30 seconds, and 72°C for 5 minutes for 33 cycles. The reactions were ended with a final extension at 72°C for 10 minutes. The primers used are given in Table 4.
Primers for Amplification of Fragment Spanning Target Site
Production of transgenic and nontransgenic embryos by handmade cloning
For handmade cloning (HMC), transgenic and nontransgenic cells were grown to full confluency for maximal synchronization in the G1 stage of the cell cycle as described previously (Selokar et al., 2012). The recipient oocytes were prepared by in vitro maturation, cumulus, and zona removal, and finally, manual enucleation as described in an earlier study (Shah et al., 2009) with slight modifications as increased duration of in vitro maturation for 24 hours, hyaluronidase treatment for 5 minutes instead of 1 minute followed by gentle pipetting for 1 minute, and finally reducing the duration of pronase treatment from 8 minutes to 6 minutes. During electrofusion, a single DC pulse (2.3 kV/cm for 8 μseconds) was applied.
The reconstructed, activated embryos were cultured in 400 μL of G1/G2 medium (Vitrolife) in a 4-well dish (10–15 reconstructed embryos per well) overlaid with mineral oil and incubated undisturbed at 38.5°C in a 5% CO2 incubator for 7 days. The blastocyst rate recorded on day 8 of in vitro culture (IVC) was taken as a measure of the in vitro developmental competence of the embryos.
Single embryo PCR
GFP expressing embryos of morula and blastocyst stage (n = 10) were randomly picked, and each was added to a 0.2-mL tube in 2–3 μL of water and lysed by heating at 95°C followed by immediate chilling on ice to extract the DNA to be further used as the template. The PCR was carried out to detect GFP expression as follows: initial denaturation at 95°C for 3 minutes; 35 cycles of 95°C for 30 seconds, 60°C for 30 seconds, 72°C for 30 seconds; and a final extension at 72°C for 10 minutes. The primers used are given in Table 5. Single-embryo PCR for 15 embryos was also performed using primers GR5′END F/CMV-R3 (5′ junction, 1679 bp product) and primers SV40 F/GR3′END R (3′ junction, 1857 bp product) for screening the correctly targeted embryos.
Primers for Amplification of GFP
Statistical analysis
GraphPad Prism 7 software was used for statistical analysis. The differences among means were analyzed using one-way analysis of variance to compare developmental competence of cloned embryos and by Student's t-test to compare knockin efficiencies between circular and linear donor vector. The percentage values were subjected to arcsine transformation before analysis. The differences were considered to be statistically significant at p < 0.05. Data are presented as mean ± standard error of the mean.
Results
Identification, cloning, and sequencing of goat Rosa 26 locus
Goat Rosa 26 locus was identified by aligning porcine Rosa 26 sequences with goat genome where a sequence located on chromosome 22 exhibited 86.50% similarity (Supplementary Fig. S1). The sequence was further aligned with sheep Rosa 26 sequences, where it exhibited 96% sequence conservation, which confirmed the bonafide identity of this region as goat Rosa 26 locus. The sequence containing the potential intron 1 region of Rosa 26 locus (2.2 kb) was isolated and amplified from caprine genomic DNA (Supplementary Fig. S2). The purified PCR product was cloned and sequenced and shared 99.90% similarity to the putative Rosa 26 sequence on goat chromosome 22.
Generation of a TALEN pair and Rosa 26 targeting vector P5′KC3′
A TALEN pair, that is, left and right TALEN vectors, were designed and generated from Life Technologies Corporation against the intron 1 region of Rosa 26 locus (Supplementary Fig. S3). These TALEN vectors were separated by a 16 bp spacer, where the FokI endonuclease would create a double-strand break, which would later be repaired by homology-directed repair in the presence of donor vector. The Rosa 26 targeting vector P5′KC3′ (pIRES 5′ KIGS CMV 3′) was constructed and verified by PCR and restriction digestion (Fig. 2b). The vector was designed to introduce the GFP gene cassette (CMV-KAN-IRES-GFP-SV40pA) at intron 1 of Rosa 26 locus. 5′ and 3′ goat Rosa 26 homology arms flanking GFP gene cassette were designed for efficient homologous recombination.
Establishment and characterization of GFFs
GFFs were successfully established from fetal ear tissue explants. The cells started originating from explants after 3–4 days of culture and formed a confluent layer within 8–9 days. The cells appeared spindle shaped and expressed fibroblast-specific markers, tubulin and vimentin, as determined by immunocytochemistry (Fig. 3), whereas the expression of epithelial cell-specific markers keratin 78 and cytokeratin-18 was not observed in these cells. Furthermore, verification was done by RT-PCR, where the cells were positive for VIMENTIN and TUBULIN expression and negative for CYTOKERATIN-8 and CYTOKERATIN-18 (Fig. 4). PCR amplification from genomic DNA showed the presence of X chromosome-specific PLP gene and absence of Y chromosome-specific SRY gene, which confirmed that the cell used in the present study was from the female fetus in origin (Fig. 4).
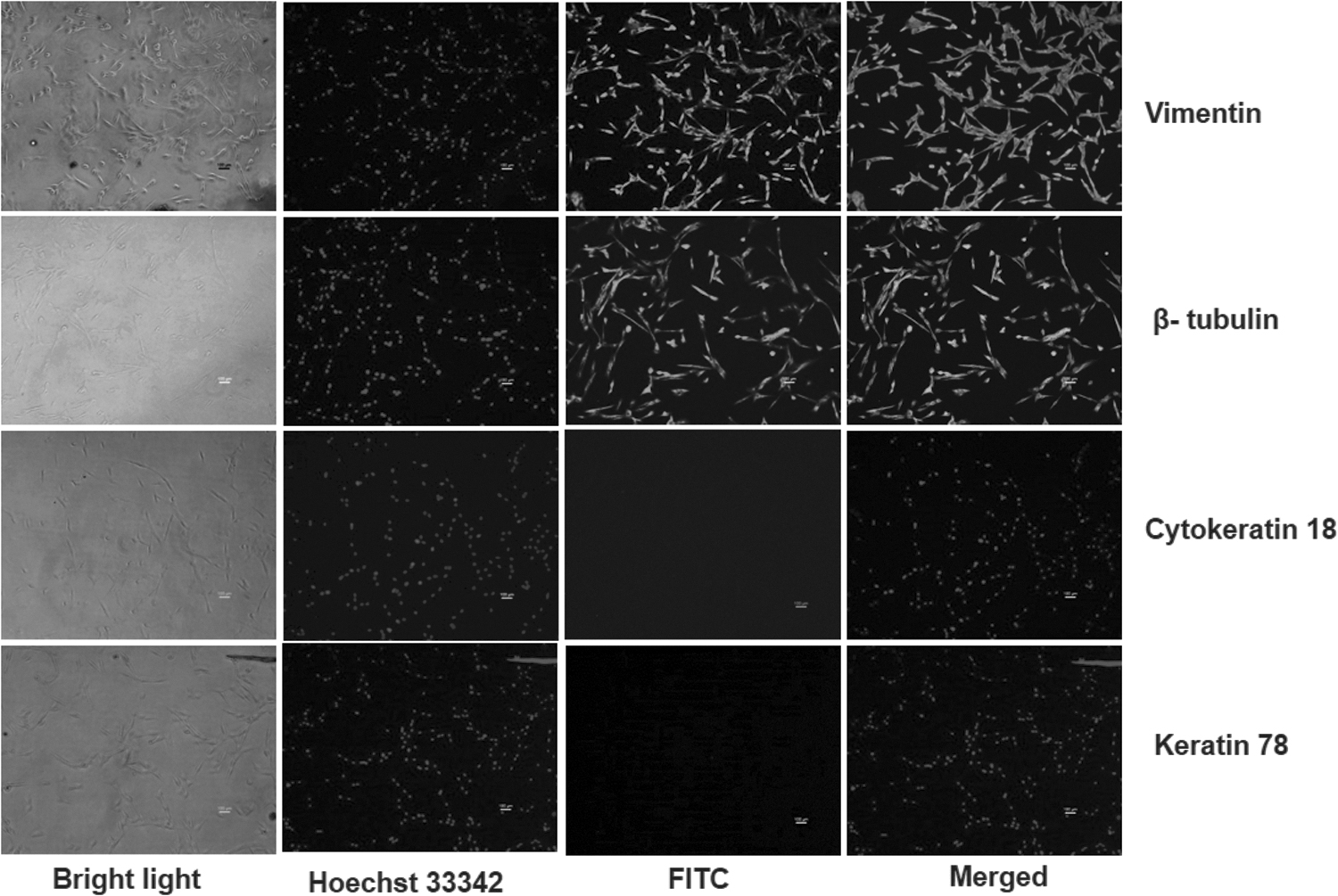
Characterization of GFF cells by immunocytochemistry: cells under bright light, UV light for visualization of nucleus stained using Hoechst 33342, fluorescent light for observing cells stained with FITC-conjugated 20 Ab and merged images of 20 Ab and Hoechst (magnification: 100 × ). GFF, goat fetal fibroblast; UV, ultraviolet.

Characterization of GFFs and sex confirmation:
Evaluation of the efficiency of TALENs
To evaluate the efficiency of TALENs, GFFs were transfected by TALEN pair, and genomic DNA was isolated from transfected cells after 72 hours of culture. Fragment spanning the target site was amplified using specific primers (TAL F and TAL R), and a 143 bp band was observed. The amplified fragment was cloned, and the sequence analysis of 30 recombinant clones showed 4 variants having indels in the spacer sequence (Fig. 5). A 13.30% indel frequency was observed, which confirmed the TALEN pair's efficient binding at the Rosa 26 locus.
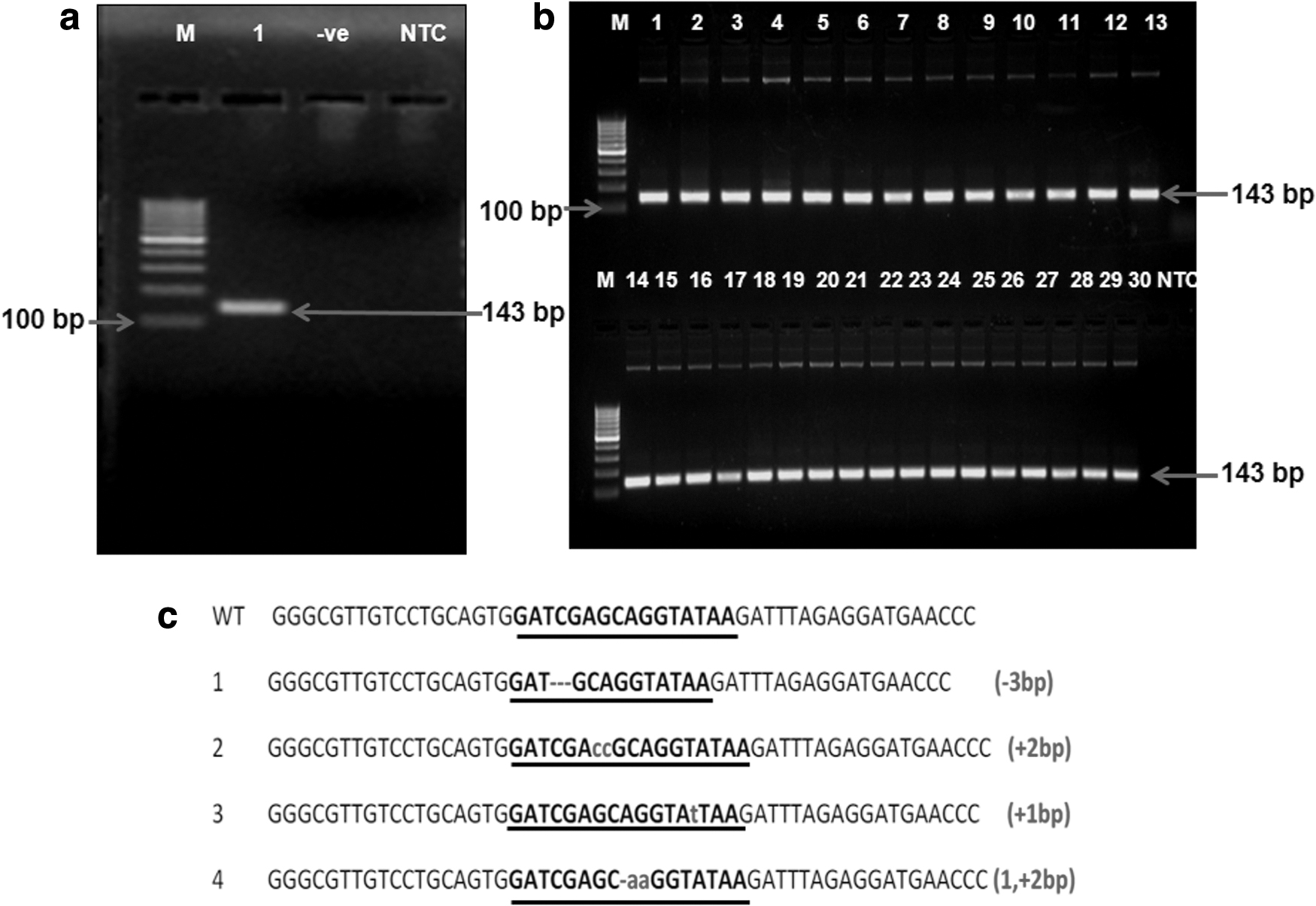
Evaluation of TALEN efficiency:
TALEN-mediated targeting of donor vector at Rosa 26 locus
Transfected cell colonies were screened for specific integration by 3′ and 5′ junction PCR. 3′ Junction PCR using primer SV40 F from endogenous vector and GR3′END R from exogenous DNA from right hand junction gave the desired band of 1857 bp in only 1 out of 24 colonies screened-in circular vector (Fig. 6), whereas 3/24 positive colonies were obtained in cells transfected with linear vector fragment (Fig. 7). The integration of the donor vector at Rosa 26 locus was further confirmed by obtaining the desired band of 1679 bp in 5′ junction PCR using CMV R3 and GR5′END F primers from the left junction region. Linear donor vector fragment (12.85% ± 1.43%) showed nonsignificantly (p = 0.08) higher frequency of homologous recombination when compared with the circular donor vector (3.70% ± 3.70%) (Supplementary Table S1).
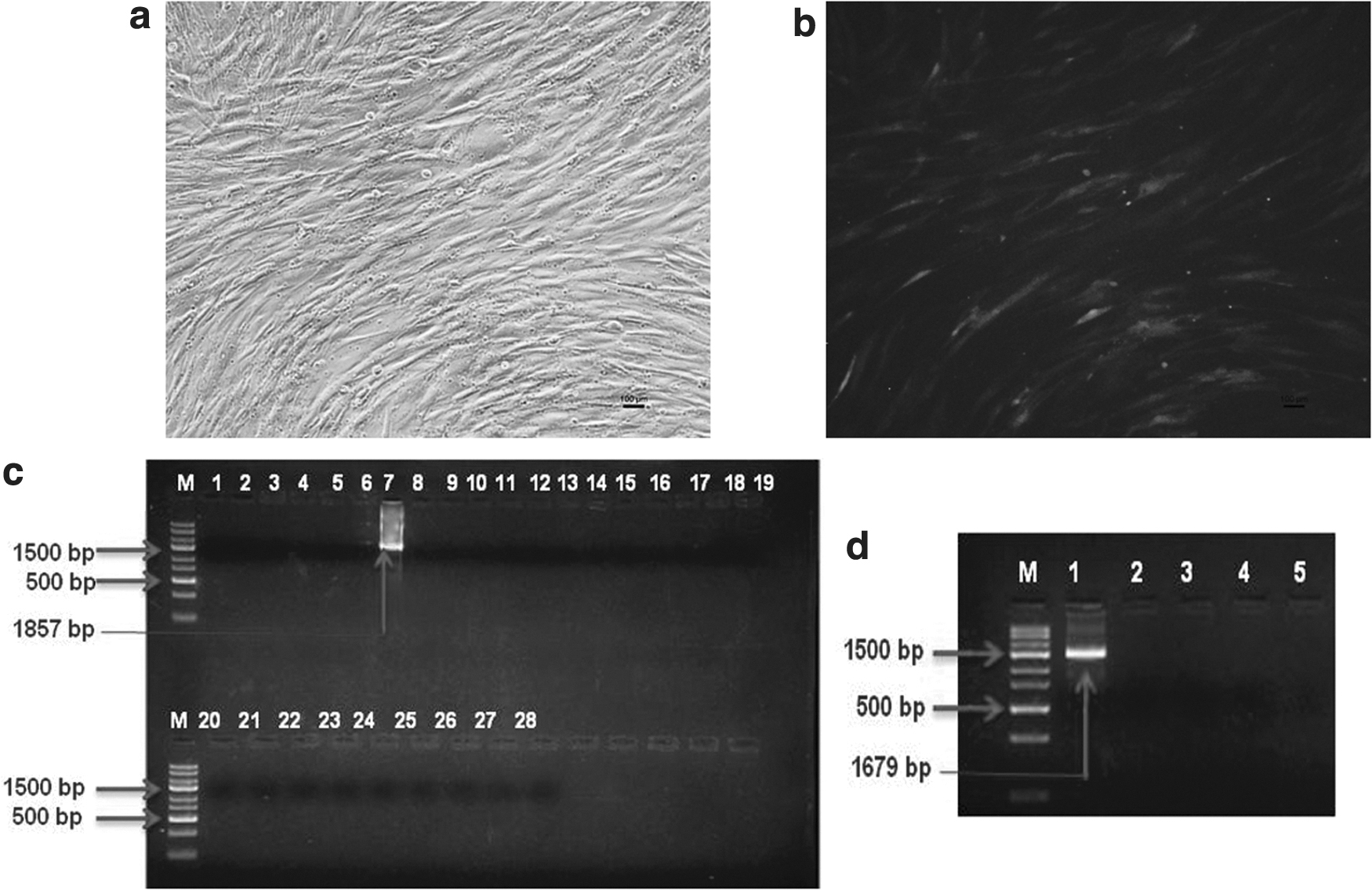
Cotransfection of TALEN and circular donor vector into GFF by nucleofection:

Cotransfection of TALEN and linear donor vector into GFF by nucleofection:
Screening targeted cell colonies for mono and biallelic gene insertions
When the four positive colonies were analyzed for the presence of unmodified allele using primers spanning the target site, the presence of 5 kb band and the endogenous 143 bp band in all demonstrated that they had monoallelic gene insertions (Fig. 8). Biallelic modifications were not observed in any of the correctly targeted cell colonies.

Screening monoallelic and biallelic gene insertions: Lane 1–4: positive cell colonies for 3′ and 5′ junction PCR, Lane 5: nontransfected cells, Lane 6: negative enzyme, Lane 7: NTC, M: DNA ladder 5 kb.
Production of transgenic goat embryos by HMC
Transgenic cells having specific integration were used as nuclear donors to generate cloned transgenic embryos expressing GFP. The blastocyst rate was significantly lower (p < 0.05) for transgenic cloned embryos than control (3.92% ± 1.12% vs.7.84% ± 0.68%; Table 6), although there was no significant difference between these two groups for the earlier stages of embryonic development.
In Vitro Developmental Competence of Cloned Embryos Produced from Transgenic and Nontransgenic Fetal Fibroblast Cells
Values are mean ± SEM. Values with different superscript within the same column differ significantly (p < 0.05).
SEM, standard error of the mean.
Screening of correctly targeted embryos at Rosa 26 locus
The fluorescence microscope-based evaluation confirmed that all the transgenic cloned embryos expressed GFP, which was further validated by the presence of a 181 bp band of GFP gene in single-embryo PCR (Fig. 9). Furthermore, the presence of consistent size bands in 2 out of 15 (13.30%) embryos (morulae and blastocysts) in single-embryo 5′ and 3′ junction PCR confirmed the successful integration and expression of donor vector at Rosa 26 locus (Fig. 10).

Production of goat transgenic embryos using targeted fibroblasts:
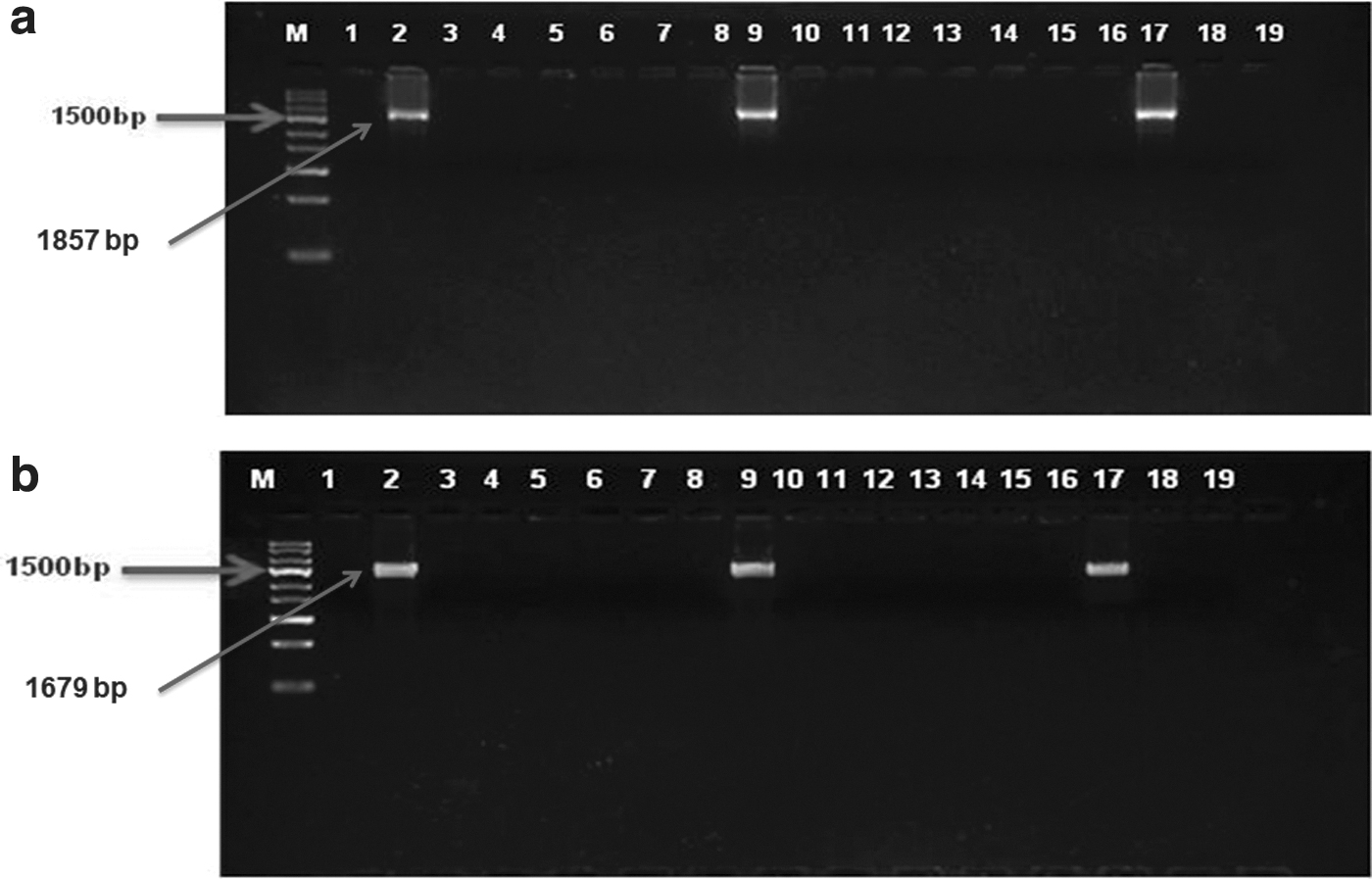
Screening of goat transgenic embryos for targeted modification:
Discussion
In the present study, we identified the Rosa 26 locus in the goat genome and generated transgenic handmade cloned embryos following TALEN-mediated targeting of GFFs with a donor vector containing a GFP gene cassette. The identified caprine Rosa 26 locus exhibited very high homology with the porcine and ovine Rosa 26 sequences, which confirmed its bonafide identity. After that, we targeted the intron 1 region of the Rosa 26 loci for knockin of the GFP gene cassette considering the porcine Rosa 26 target site (Kong et al., 2014). The Rosa 26 locus supported stable and ubiquitous expression of the GFP gene in the donor GFFs and transgenic cloned embryos.
Numerous earlier studies have demonstrated the pre-eminence of TALENs over other genome-editing tools because of their greater targeting efficiency, higher specificity, and lower cytotoxicity (Kasparek et al., 2014; Mussolino et al., 2014; Shi et al., 2017; Zhu, 2015). Therefore, we chose TALENs for site-directed transgene integration at the Rosa 26 locus and generated a Rosa 26 targeted expression vector (donor vector- P5′KC3′) containing the GFP gene, which any desired transgene could further replace. The GFP gene cassette in the vector was flanked by 864 bp 5′ and 1283 bp 3′ homology arms to facilitate efficient homology-directed repair as the use of longer homology arms results in a higher frequency of homologous recombination (Cui et al., 2015).
In the current study, fetal fibroblast cells were chosen as they grow rapidly and serve as an efficient nuclear donor for SCNT (Liu et al., 2013; Mehta et al., 2018, 2019). Several studies have demonstrated that nucleofection, an improved version of electroporation, exhibit very high transfection efficiency and less cytotoxic effects for various cell types, including fetal and adult fibroblasts, embryonic stem cells, and spermatogonial stem cells (George et al., 2011; Mehta et al., 2018; Sharma et al., 2020). Therefore, we chose nucleofection to deliver the constructed donor vector along with TALEN vectors in GFFs. The transfection of GFFs by TALEN vectors resulted in 13.30% indel frequency at Rosa 26 locus, which confirmed the functionality of TALENs and was subsequently used for cotransfection with Rosa 26 targeting vector.
The frequency of homologous recombination for integrating the GFP gene cassette at Rosa 26 locus was higher for the linear donor vector fragment than the complete circular construct. It could be due to the larger construct size of the circular vector than the linear vector fragment, as reported by Kasparek et al. (2014). Contrastingly, some previous studies have shown no difference in integration between circular and linearized vectors (Cui et al., 2011; Hermann et al., 2012; Meyer et al., 2010).
We found only monoallelic modifications in all the correctly targeted colonies. Since it is challenging to control the mono- and biallelic targeting ratio, the only viable option is to screen the maximum number of colonies. However, improvement in certain factors, such as transfectability, the efficacy of nucleases, and the quality of donor vector, might yield a higher efficiency of homology-directed repair, ultimately resulting in higher biallelic modifications (Luo et al., 2014).
The lower blastocyst rate for the transgenic cloned embryos than the cloned embryos used as control suggests that the presence of transgene or G418 selection might have an adverse effect on their developmental competence. These results agree with several earlier studies (Gui et al., 2012; Jura et al., 2013; Mehta et al., 2019; Xiong et al., 2011). Two out of 15 embryos (13.30%) showed the desired genome editing. The genome-editing efficiency may be further improved by developing a better strategy to select the culture of cells with targeted integration (Luo et al., 2014). After that, using these cells to generate transgenic handmade cloned embryos could culminate in the production of 100% correctly targeted embryos. Finally, based on our results, it can be inferred that the identified goat Rosa 26 locus is amenable to TALEN-mediated gene knockin, and it supported the stable expression of the GFP gene in donor GFFs and transgenic cloned embryos.
The present study was confined to identify a safe locus in the goat genome and optimize TALEN-mediated GFP gene targeting in GFFs and embryos. However, the strategies for targeted cell colony selection and embryo production need to be further improved. In this context, the seeding of transfected cells at a low density before G418 selection could offer better results than the cloning ring method used to obtain single cell-derived colonies. The generation of single cell-derived knockin colonies would ensure that all the transgenic cloned embryos generated using these cells would have the same genotype. Future studies should focus on improving the developmental competence of transgenic handmade cloned goat embryos and generating transgenic animals followed by their transfer to surrogate females.
In conclusion, we identified Rosa 26 locus in the goat genome and demonstrated the transgene insertion in GFFs through TALEN-mediated homologous recombination. The GFP gene incorporated at Rosa 26 locus was expressed ubiquitously in both GFFs and transgenic handmade cloned embryos. Hence, this study provides headway for targeted integration of medicinally/pharmaceutically important genes to generate transgenic goats.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.