
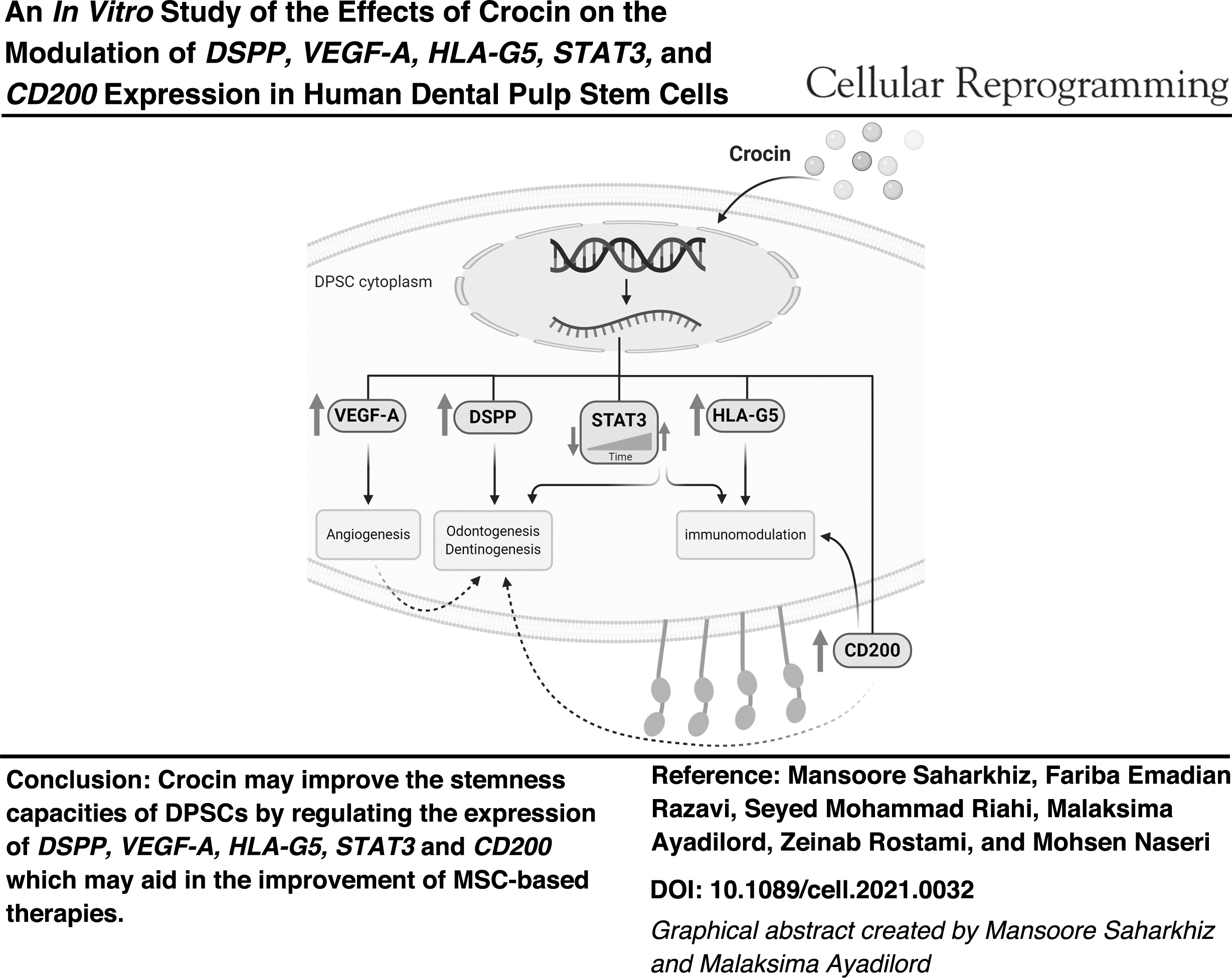
Abstract
Dental pulp stem cells (DPSCs) have been recommended as promising candidate for cell-based therapeutic applications due to high potentials in tissue repair/regeneration and modulation of immune responses. The gene expression change strategy by natural plant enhancers is an available opportunity to improve the stemness properties of these cells. The objective of this research was the evaluation of Crocin effects (saffron plant's bioactive compound) on immunoregulation and tissue regeneration-related biomarkers expression in human DPSCs. Based on the results of cell viability assay, application of 400 μM and lower concentrations of Crocin had no toxic effects on DPSCs; however, the time-dependent cytotoxic effects were observed at higher concentrations. This study, probably for the first time, detected the surface expression of CD200 in DPSCs with a slight time-dependent upward trend and reported that treatment with Crocin could increase expression of this macromolecule up to many times over. Also, it revealed that this carotenoid significantly led to the time-dependent upregulation of dentin sialophosphoprotein, vascular endothelial growth factor A, human leukocyte antigen-G5, and signal transducer and activator of transcription-3 messenger ribonucleic acids (mRNAs); however, this significant upregulation for STAT3 occurred, followed by a remarkable reduction. The results of this study indicated that cell treatment with Crocin may be effective in improving the stemness capacities of DPSCs. Therefore, the study provided basis for more insights into the biological effects of Crocin on DPSCs that it may aid in the future improvement of mesenchymal stem cell-based therapies.
Introduction
Adult mesenchymal stem cells (MSCs) are a heterogeneous population of fibroblast-like cells that can be isolated from majority of adult organs such as bone marrow and adipose tissues. These cells are characterized by self-renewal capacity, potential for multidifferentiation, positive expression of mesenchymal markers of CD90, CD73, and CD105, and negative expression of hematopoietic markers of CD14, CD34, and CD45 (Lin et al., 2013).
Recently, studies have reported that there are new populations of MSCs in the oral cavity, which are isolated from various dental tissues, including dental pulp stem cells (DPSCs), periodontal ligament stem cells, gingival stem cells, apical papilla stem cells, tooth germ stem cells, and stem cells from dental follicles (Gan et al.,, 2020). Among these cells, DPSCs were first recognized in 2000, able to differentiate to various lineages of mesenchymal tissues in vitro (Iohara et al., 2006) together with the potency to generate dental pulp-dentin complex (Gronthos et al., 2000) through the expression of macromolecules such as dentin sialophosphoprotein (DSPP) and vascular endothelial growth factor (VEGF), which involve in processes relating to angiogenesis, dentinogenesis, and odontogenesis in vivo (Nakashima et al., 2009; Zhu et al., 2012).
Recently, it has been shown that these cells possess a powerful immunomodulatory capability. This immunomodulation ability is due to humoral impacts mediated by cytokine, molecule, and growth factor secretion such as prostaglandin E2 (PGE2), indoleamine 2,3-dioxygenase (IDO), interleukin-6 and interleukin-10 (IL-6 and IL-10), nitric-oxide (NO), transforming growth factor-beta (TGF-beta), human leukocyte antigen-G5 (HLA-G5), and direct contact impacts with cells involved in innate and adaptive immunity through the surface expression of molecules such as programmed cell death ligand-1 and programmed cell death ligand-2 (PD-L1, PD-L2), membrane-bound HLA-G1 (Andrukhov et al., 2019; Hossein-Khannazer et al., 2019), and probably CD200 (OX2) similar to other MSCs (Kawasaki et al., 2007; Kim et al., 2018; Li et al., 2019). Furthermore, recent studies have highlighted the impressive role of signaling molecules such as STAT (signal transducer and activator of transcription) family of proteins, both in differentiation and in immune modulation process, by these cells (Vigo et al., 2019; Xu et al., 2018).
However, it has not been more than two decades from the isolation of first DPSCs, but because of the cell's unique properties and even higher potential compared to other MSCs in tissue repair/regeneration and modulation of immune responses and lower immunogenicity, higher proliferation rate, and easier access using teeth with indication for extraction due to orthodontic or other medical reasons (Sui et al., 2020), this short period of time was enough to introduce them as a candidate for therapeutic applications (Alhazzazi and Alghamdi, 2021; Relaño-Ginés et al., 2019), including cell-based therapy for inflammatory diseases (Andrukhov et al., 2019; Zayed and Iohara, 2020), autologous or allogeneic transplantations (Chen et al., 2015; Hata et al., 2015; Omi et al., 2016; Iohara et al., 2018), endodontic and periodontal restoration in tooth tissue (Shi et al., 2020), and extraoral tissue repair/regeneration (Yamada et al., 2019).
Recently, investigators are looking to discover new opportunities to optimize the therapeutic effectiveness and stemness status of MSCs before clinical applications (Sequeira et al., 2021). In this regard, many later studies have focused on increasing the expression of target genes (García-Bernal et al., 2021; Kornicka et al., 2017). The usage of herbs or their effective compound-based strategies has opened up new landscapes in this context (Kornicka et al., 2017).
Crocin is the most important carotenoid derived from saffron (Crocus sativus) plant, in which numerous pharmacological effects have been reported, including anti-inflammatory (Hashemzaei et al., 2020; Pashirzad et al., 2019) and tissue regenerative effects (Firouzi et al., 2010; Terraf et al., 2017; Tamaddonfard et al., 2013). Based on the GO (Gene Ontology) database and the previous studies, we selected the most common and important biomarkers related to immunomodulation and regeneration of dental pulp and dentin tissues. Then we assessed impacts of Crocin on relative expression of DSPP, VEGF-A, HLA-G5, and STAT3 genes as well as surface expression of CD200 glycoprotein in these cells. So this study was conducted to evaluate effects of Crocin at different time intervals on properties of the human DPSC cells to improve their efficiency in therapeutic applications.
Materials and Methods
Cell isolation and culture
The study was managed in accordance with the Declaration of Helsinki and approved by the ethics committee of the Birjand University of Medical Sciences (IR.BUMS.REC.1399.090). A human third molar tooth with complete root development was collected from a patient who had undergone teeth extraction due to orthodontic reasons in Rasool Akram dental clinic, Birjand, Iran. Then the DPSC isolation was performed by enzymatic digestion method according to a published protocol with a slight change (Gronthos et al., 2011).
Briefly, after transfer of the tooth to laboratory and exerting mechanical fractures on its surface, the pulp tissue was removed by means of forceps and then immediately immersed in phosphate-buffered saline containing 100 U/mL type I collagenase enzyme (Bio-Idea, Tehran, Iran) for 1 hour at 37°C. Digested pulp was put in Dulbecco's modified Eagle's medium/F12 (DMEM/F12; Gibco, Grand Island) complemented with 10% fetal bovine serum (FBS; Biowest, Nuaillé, France), 100 U/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (all Sigma-Aldrich, St. Louis, MO) and then incubated in 5% CO2 at 37°C for 5 days. When the primary cells reached a maximal growth (85%–90% confluence in 25-mm Petri dish), these were harvested using 0.05% trypsin/EDTA (Biowest). The early subcultures were started at 6 × 105 cells/mL in T-25 cell culture flasks. In this research, all experiments were carried out with cells between 3 and 5 passages.
Characteristics of DPSCs
Immunophenotyping and differentiation experiments contributed to characterize and confirm the isolated DPSCs.
Osteogenic and adipogenic differentiation assay
To induce osteogenic and adipogenic differentiation, the cells were cultured in the standard medium with 10% FBS in six-well plates at a density of 100,000 cells/well. After 3 days, the usual culture medium was exchanged with osteogenesis and adipogenesis differentiation medium, respectively, including 0.1 μM dexamethasone, 10 mM beta-glycerophosphate, 50 μM ascorbate-2-phosphate and 0.5 mM isobutyl-methyl-xanthine, 1 μM dexamethasone, and 200 μM indomethacin (all from Sigma-Aldrich).
The inducible culture media were refreshed each 2 days and it was continued for 21 days in adipo-differentiation and 28 days in osteo-differentiation. Finally, to evaluate the rate of differentiation to each of lineages, alizarin red and oil red staining were performed (Al-Habib and Huang, 2019). The stained cells were investigated by an inverted phase-contrast microscope for detection of synthesized mineral nodules and fat droplets.
Immunophenotyping assay
Flow cytometry method was done to detect the expression or lack of expression of specific MSC markers by staining with monoclonal human antibodies: CD105-PE, CD90-APC, CD73-PE CY7, and CD45-FITC (all from eBioscience, San Diego, CA).
Cell viability assay
The highest nontoxic dose of Crocin (Sigma-Aldrich) on the DPSCs was determined at 24, 48, and 72 h by MTT procedure. For this purpose, the cells were seeded in 96-well plates (104 cells/implant) and incubated to attach overnight at 37°C in 5% CO2. On the next day, they were treated with various amounts of Crocin (from 6.25 to 450 μM) for 24 hours. At the end of treatment, 20 μL solution of 3-(4, 5-dimethylthiazol-2-yl)-2 (MTT) (5 mg/mL, pH = 7.4) was added to each well and put at 37°C for 3 hours. The media were aspirated completely and formazan crystals became soluble in 100 mL dimethyl sulfoxide. Subsequently, the absorption (OD: optical density) of each well of the culture plate was measured at wavelengths of 570 and 630 nm using a scanning spectrophotometer (Biotek Epoch, Winooski, VT) (Vega-Avila and Pugsley, 2011).
Based on the results of statistical analysis of data in 24 h, concentrations of Crocin that did not have any significant effects on the cytotoxicity and growth of DPSCs were selected to investigate the cell viability after exposure with them for 48 and 72 hours. Finally, the highest concentration of Crocin, which had no significant effect on the cells' viabilities at 24, 48, and 72 h, was selected for subsequent experiments.
Quantitative real-time polymerase chain reaction (PCR) analysis
After treatment of DPSCs with Crocin, the RNA extraction was performed (Pars Tous, Tehran, Iran) and quantified at 260/280 nm with a NanoDrop (Biotek Epoch). Then cDNA was synthesized from 2 μg total RNA (Pars Tous). The relative expression of HLA-G5, VEGF-A, STAT3, and DSPP genes was evaluated by using SYBR Green assay in total volume of 25 μL as three replicates with two technical repeats. In the analysis, expression of target genes was normalized against GAPDH as an endogenous control using the 2−ΔΔCt method. The sequences of primers used are indicated in Table 1.
The Sequence of Primers Used in Quantitative Real-Time Polymerase Chain Reaction (PCR) Analysis
DSPP, dentin sialophosphoprotein; HLA-G5, human leukocyte antigen-G5; VEGF, vascular endothelial growth factor.
Flow cytometry analysis
The flow cytometric analysis of CD200 surface expression was performed on Crocin-treated and untreated cells. Approximately 1,000,000 cells were stained using the phycoerythrin (PE)-conjugated anti-human CD200 monoclonal antibody and PE-mouse IgG1κ as isotype control (both from Biolegend, San Diego) according to protocol optimized by Al-Habib and Huang (2019). Finally, the stained cells were counted with the flow cytometer (Sysmex®).
Statistical analysis
All tests were repeated at least for three experiments and reported as the mean ± standard deviation, after analyzing by SPSS software (version 26). One-way analysis of variance (ANOVA) followed by Tukey's test was performed to detect the highest optimal and nontoxic concentration of Crocin. The Rest software (t-test) was utilized to assess the gene expression results. Flow cytometry data were analyzed using FlowJo software, repeated measure ANOVA, and t-test. Also, in this study, GraphPad Prism software 8 was used for graphing and p < 0.05 was defined as statistical significance level.
Results
Cell isolation and culture
Human DPSCs were successfully isolated from a third molar tooth and expanded on the basis of high adhesion capacity to plastic surfaces. On the fifth day of isolation, the digested pulp had perfectly adhered to the floor of the culture dish and the fibroblast-like spindle cells had migrated out and poorly proliferated around it. These cells were heterogeneous in shape and size. During 2 days, a large cell colony was established, so the pulp tissue was taken out. After incubation for 5 days, the primary cells attained ∼80% monolayer confluence. The cells were harvested and used for subcultures (Fig. 1a–c).
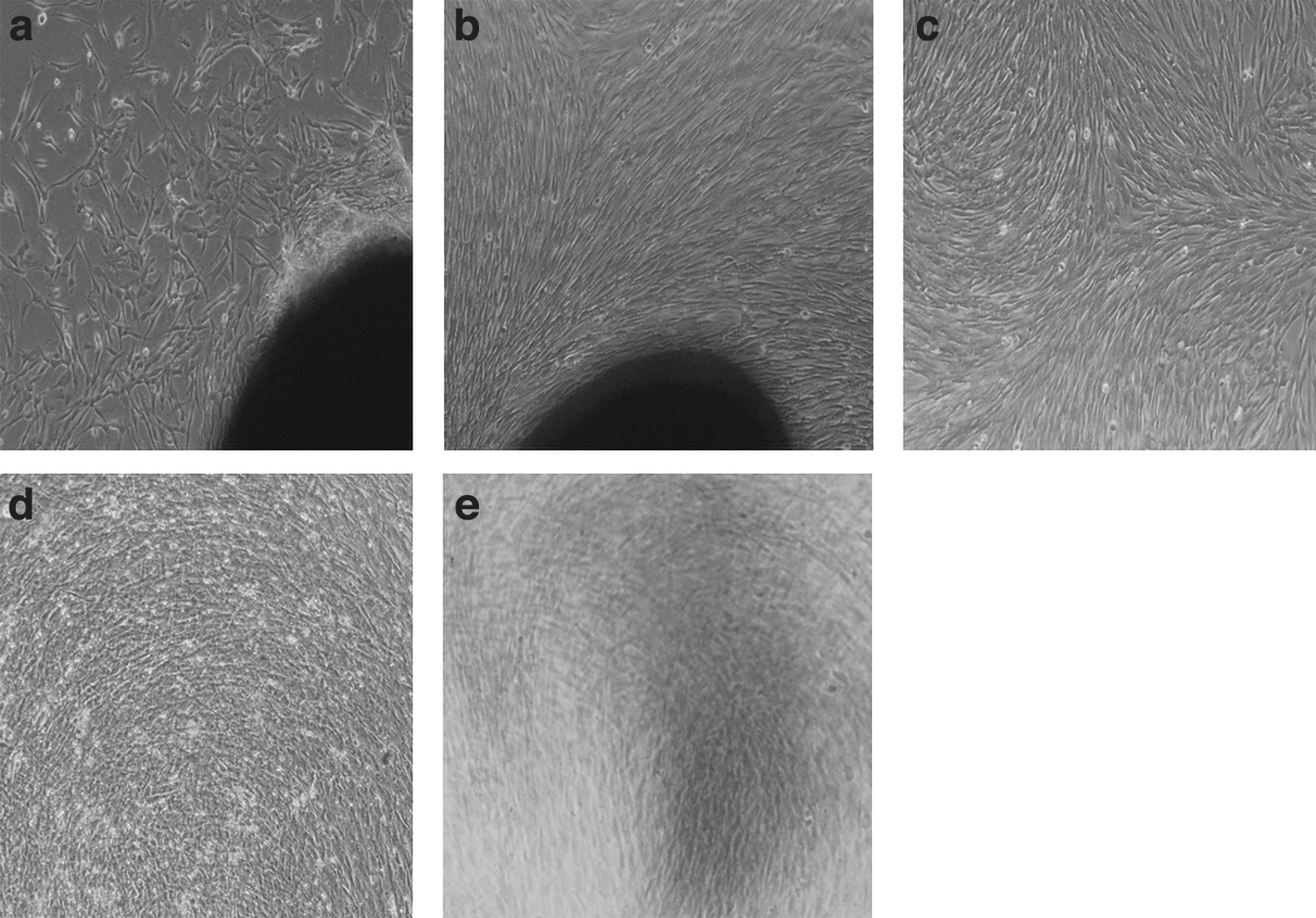
HDPSC isolation and differentiation to osteogenic and adipogenic lineages;
Differentiation toward osteogenic and adipogenic lineages
To characterize and confirm the DPSCs, the capacity to differentiate into osteoblasts and adipoblasts using the suitable culture and media conditions was investigated. Approximately 21 days after the onset of differentiation into adipose tissue, lipid droplets could be detected in cell cytoplasms, like adipocytes by oil red staining. On the other hand, after 28 days of osteogenesis culture, alizarin red staining approved the formation of mineralized nodules (Fig. 1d, e).
Immunophenotypic characterization of DPSCs
In flow cytometric analysis of cell surface markers, the isolated cells were strongly positive for specific mesenchymal markers CD73, CD90, and CD105, but negative for characteristic hematopoietic marker CD45 (Fig. 2).
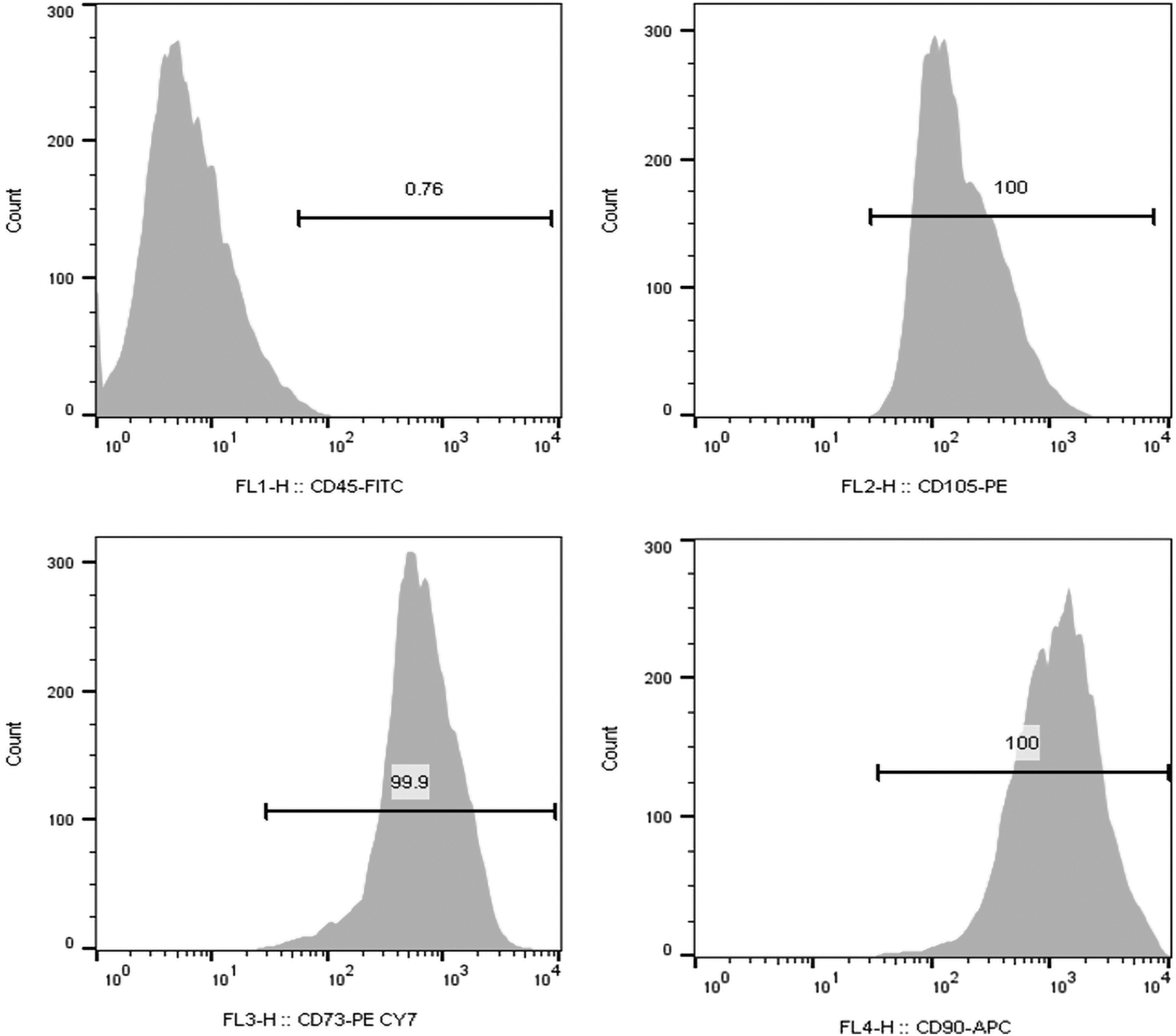
Immunophenotypic characterization of DPSCs. Flow cytometry analysis revealed that extracted cells were positive for specific mesenchymal markers CD90, CD73, and CD105, but negative for hematopoietic marker CD45.
Determination of the highest nontoxic concentration of Crocin on DPSCs
The DPSCs indicated a considerable growth rate under treatment with Crocin at concentrations less than or equal to 375 μM after 24 h (p < 0.05 and p < 0.001), while no statistically remarkable difference was found in terms of cell toxicity or proliferation at 400–450 μM concentration compared to control group (p > 0.05). To confirm the results, the experiment was repeated for concentrations of 375 to 425 μM. According to the results obtained, Crocin cytotoxicity in concentration of 425 μM occurred after 72 hours (p < 0.001). Thus, for subsequent experiments, the concentration of 400 μM of Crocin was selected as the highest nontoxic concentration in 24, 48, and 72 hours (p > 0.05) (Fig. 3).

Viability assay for DPSCs incubated with Crocin. The results were presented in mean absorption ± SD. Significant differences compared to control were indicated: *p < 0.001; **p < 0.05. SD, standard deviation.
Positive effects of Crocin on the expression of immunomodulatory and regenerative markers detected by quantitative reverse transcription polymerase chain reaction (PCR)
According to the Q real-time polymerase chain reaction (PCR) data, treatment with Crocin significantly resulted in a time-dependent upexpression of DSPP, VEGF-A, and HLA-G5 messenger ribonucleic acids (mRNAs), while this significant increase for STAT3 occurred after a statistically considerable reduction (all p < 0.001) (Fig. 4).
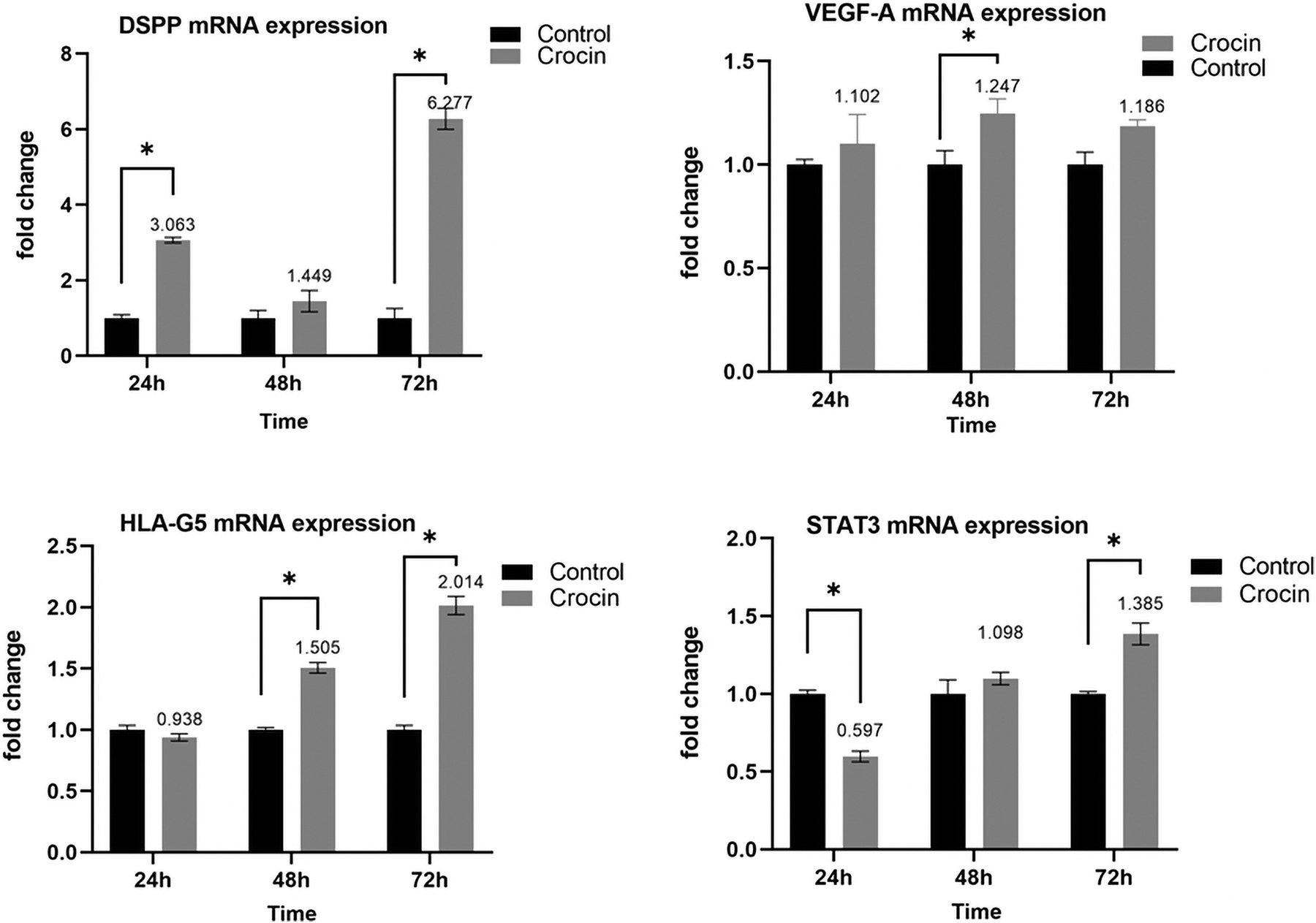
Quantitative real-time PCR analysis of DSPP, VEGF-A, HLA-G5, and STAT3 markers after Crocin treatment. Results were presented as mean ± SD. “*” Significant change (p < 0.001); DSPP, dentin sialophosphoprotein; HLA-G5, human leukocyte antigen-G5; PCR, polymerase chain reaction; STAT3, signal transducer and activator of transcription-3; VEGF-A, vascular endothelial growth factor-A.
Positive effects of Crocin on the cell surface expression of CD200 detected by flow cytometry
According to flow cytometric data, untreated DPSC cells expressed CD200 glycoprotein with a slight time-dependent upward trend (24 hours, 3.46%; 48 hours, 3.78%; and 72 hours, 7.75%), while treatment with Crocin considerably increased the positive cell population to 4.97%, 12.36%, and 20.78% after 24, 48, and 72 hours, respectively (Fig. 5a). Statistical analysis of the data with repeated measure ANOVA test revealed that the CD200 expression linearly enhanced over time (p < 0.001) and Crocin caused a significant difference in the percentage of positive cells, compared to the control groups (on average 12.345% in treatment group vs. 4.853% in control group, p = 0.003, partial eta squared = 0.995) (Fig. 5b, c). Also, Mauchly's test was performed to check the assumption of sphericity (Mauchly's W = 0.259, df = 2, p = 0.509).
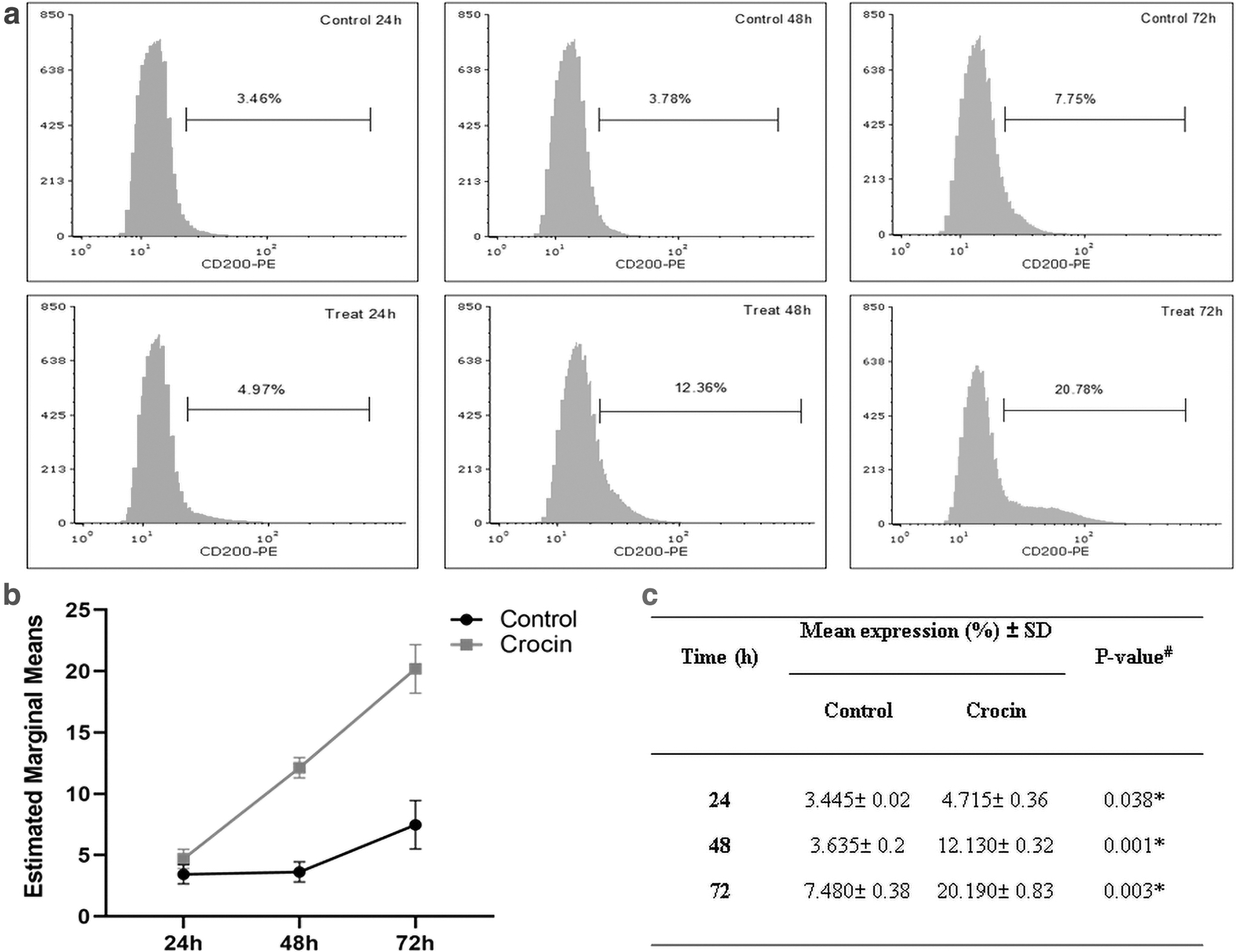
Flow cytometric analysis of CD200 surface expression in DPSCs;
Discussion
MSC-based therapy is being intensely considered a promising strategy for the treatment of degenerative and immune-related diseases in recent years. Among various sources of MSCs, cells isolated from dental pulp have opened up a new opportunity in medicine and especially in dentistry due to their unique capabilities.
The aim of this study was to provide preliminary data about the effects of Crocin on the DPSCs with the hope that the results of the existing studies along with other studies in the future will introduce new opportunities to optimize the therapeutic effectiveness and stemness status of MSCs. Therefore, in this study, the immunoregulation and tissue repair/regeneration-related biomarkers expression were evaluated in Crocin-treated DPSCs on the assumption that this carotenoid can significantly upregulate them. For this purpose, first, we established the DPSC culture from a human third molar tooth and demonstrated that these cells were compliant with ISCT (International Society for Cell & Gene Therapy) criteria in terms of morphology, surface marker specification, and differentiation potential.
Almost all studies agree that the reaction of CD200 with its receptor on the surface of myeloid cells reduces inflammatory responses. Therefore, immune inhibition enforced by CD200/CD200R can be an effective step in allogeneic transplants and treatment of diseases related to the overactivity of the immune system (Holmannová et al., 2012).
It has been reported that most cancer stem cells and some MSCs express CD200 glycoprotein on their surface (Kawasaki et al., 2007; La Rocca et al., 2012). Based on the information obtained, no study has been performed to evaluate expression of this macromolecule on dental stem cells so far. Hence, possibly for the first time, we successfully detected positive expression of this marker on surface of the DPSCs and showed that treatment with Crocin can increase it up to many times over; however, other experiments are needed to confirm the effectiveness of this amount of increase.
In this regard, Kong and coworkers found that MSC cells taken from placenta amniotic membrane enhanced CD200 expression up to 2.5-fold under hypoxic conditions and indicated that co-culture of these cells with microglia cells inhibited expression of inflammatory cytokines. In addition, in vivo experiments showed that transplantation of CD200-positive cells has neuroprotective effects in stroke model rats. So their results could indicate impactability of the repair process from the inflammatory microenvironment by the modified MSCs (Kong et al., 2018).
Researchers demonstrated that MSC differentiation/immunoregulatory potentials were enhanced following the forced induction of CD200 (Kim et al., 2018), and it was agreed with the findings presented by Pontikoglou. It was shown that CD200-positive cells sorted from bone marrow more effectively upregulated gene markers of differentiation into bone. Also, at the in vitro level, these cells were more susceptible to formation of a mineralized matrix (Pontikoglou et al., 2016).
On the other hand, we revealed that Crocin can significantly upgrade the angiogenesis and immunoregulatory properties of DPSCs by increasing the expression of VEGF-A and HLA-G5 biomarkers, respectively.
Selmani and colleagues evidenced that HLA-G5 expression by MSCs is necessary for the immunomodulatory purposes in suppressing allogeneic T lymphocyte proliferation and natural killer cell-related functions, and to induce expansion of regulatory T cells (Selmani et al., 2008). In confirmation of the results, an article of 2020 highlighted the tolerogenic potency of HLA-G molecules as novel therapeutic targets for transplant rejection and other disorders caused by immune dysregulation, and it suggested that this molecule can be a candidate biomarker for positive identification of MSCs with potent immunomodulatory capacity (Ajith et al., 2020). Also, in vitro differentiation experiments, while emphasizing the immunoregulatory functions of MSCs due to HLA-G5 secretion, indicated that expression of this molecule was preserved even after cell differentiation (Hossein-Khannazer et al., 2019; Montespan et al., 2014).
Previous studies indicated that VEGF probably is a necessary factor to increase the advancement of cell differentiation and tissue repair processes and suggested that increasing the expression of this marker may provide new opportunities to improve treatment methods in regenerative medicine approaches (D'Alimonte et al., 2011; Nakashima et al., 2009; Reis-Filho et al., 2012; Zhang et al., 2016). In this regard, Ishizaka et al. (Ishizaka et al., 2013) had demonstrated that DPSCs are more preferable to other sources of MSCs for therapeutic applications and this is because of their higher potential in the expression of angiogenic and neurotrophic factors such as VEGF-A and NGF (nerve growth factor) at the in vitro level. On the other hand, they had shown that pulp stem cells promote neovascularization, angiogenesis, and tissue regeneration more effectively than adipose and bone marrow stem cells in the in vivo experiments, in ischemic hindlimb and ischemic cerebrum models as well as ectopic dental root transplantation model.
Moreover, our results revealed the potential effects of Crocin in improving expression of pulpal differentiation marker DSPP after 24, 48, and 72 hours of cell treatment; however, the enhancement occurred to a lesser extent in 48 hours, which was also statistically insignificant. Since the expression of this biomarker is controlled by various molecular mechanisms (Hao et al., 2020; He et al., 2004; Wan et al., 2016), therefore, the study of time-dependent effects of this carotenoid on different molecular signaling pathways may be helpful to justify the results in DPSCs. Similar to the results, a recent study reported that the treatment of pre-odontoblast cells with DSPaa183–219 peptide upregulated the expression of DSPP in a time-dependent manner. In this study, the DSPP gene expression increased with different extents after 12, 24, and 48 hours of cell treatment, so that its expression at 12 and 48 hours was much higher than 24 hours (Wan et al., 2016). Also, the importance of DSPP protein had been shown that expression of this marker is necessary for dentinogenesis and regeneration of pulp-dentin complex (Kim et al., 2020; Zhu et al., 2012) so that its knockout leads to develop defective dentin mineralization in mice (Sreenath et al., 2003).
In line with the objectives of this study, Kalalinia et al. investigated the effects of two important pigments of saffron on osteogenic differentiation of bone marrow MSCs (BMSCs). They found that Crocin more effectively increased the expression of bone differentiation markers compared to Crocetin (Kalalinia et al., 2018).
A recent study at functional levels in vivo demonstrated that Crocin administration decreased histological damages and infiltration of immune cells in periodontitis models of Wistar rat (Kocaman et al., 2019), and it was in agreement with findings of Terraf et al. about using scaffold containing endometrial stem cells and Crocin for regeneration of hemisected spinal cord (Terraf et al., 2017). A research conducted by Zhu et al. showed that Crocin reduced inflammation caused by titanium particles through induction of M2 macrophage polarization both in vitro and in vivo in murine air pouch models. Also, co-culturing BMSCs and macrophages under condition of treatment with Crocin indicated that this carotenoid facilitated differentiation into bone by creating an immunoregulatory microenvironment (Zhu et al., 2019).
Furthermore, we disclosed that Crocin markedly up-regulated the expression of the STAT3 signaling molecule after a significant downregulation in it, which may be justifiable based on previous studies. It had been indicated that STAT3 can be involved in boosting the stemness potential of stem cells (Nicolaidou et al., 2012) by participating in the LIF (Leukaemia inhibitory factor) and BMP (bone morphogenic protein) signaling pathways (Chen et al., 2008).
Furthermore, the therapeutic importance of miR-21/STAT3 signal had recently been perused in DPSCs. According to the findings, the upregulation STAT3 signal dramatically improved the tendency of DPSCs toward odontoblastic lineage (Xu et al., 2018). In another study (Vigo et al., 2019), researchers extended previous findings in this field. They demonstrated that interferon beta could boost immune regulation capacity of MSCs through phosphorylation of STAT1/3. However, a recent research suggested that Crocin may decrease chronic inflammation of the cancer microenvironment by downregulating the IL6/STAT3 pathway, which is related to proliferation and invasion as well as angiogenesis in cancer cells (Kim and Park, 2018). Thus, it may be concluded that Crocin purposefully affects DPSCs in a time-dependent manner by regulating of STAT3-related pathways, which may have clinical applications.
Conclusion
This study presented preliminary data about the effects of Crocin on modulation of the immunomodulatory and tissue regeneration-related marker expression in DPSCs and suggested that Crocin may be effective to improve the stemness capacities of the cells. Therefore, the data allow to provide a basis for more insights into the biological effects of Crocin on DPSCs, which may aid in the improvement of MSC-based therapies. Although these results are promising, in future, more experiments using in vitro and in vivo models are required to prove the effectiveness of Crocin as an inducer for enhancing stemness characteristic of MSCs.
Ethical Approval
All procedures performed in this study are in accordance with the ethical standards of the institutional Birjand University of Medical Sciences (IR.BUMS.REC.1399.090).
Availability of Data Statement
All data of the study are available from the corresponding author upon reasonable request.
Consent to Participate
Informed consent was obtained from participants included in the study.
Footnotes
Acknowledgments
The authors thank the participants in this study and the research assistant of Birjand University of Medical Sciences, Birjand, Iran, for support to the study.
Author Disclosure Statement
The authors declare they have no financial conflicts of interest.
Funding Information
This study was funded by Birjand University of Medical Sciences, under grant No. 456182