
Abstract
POU5F1 (POU class 5 homeobox 1) is a transcription factor that is critically involved in the self-renewal of undifferentiated embryonic stem cells. In this present study, we have developed our study to analyze the expression of the POU5F1 in the neonatal and adult mice testis section and isolated spermatogonial stem cells (SSCs). We also examine POU5F1 protein localization by three various kinds of antibodies. In this experimental research, to enhance our understanding of the POU5F1 expression levels, protein localization, and function in testicular germ cell, we used immunohistochemistry, immunocytochemistry, and Fluidigm real-time polymerase chain reaction (RT-PCR) analysis in the mouse testis section and neonatal and adult SSCs, and also we used protein-protein network analysis and gene enrichment analysis for genes involved in testicular development. Counting POU5F1-positive cells represented significantly higher expression (p < 0.05) of POU5F1 in the adult testis in comparison to the neonate. Finally, Fluidigm RT-PCR showed a significant expression (p < 0.05) level of germ cells gene POU5F1 in neonate SSCs (1–2 week) than 16–24 week SSCs. The illustrated results identify POU5F1 as a necessary transcription factor of testicular germ cells and can be supportive for the investigation of the development and differentiation of SSCs.
Introduction
Spermatogenesis embraces a complex network of processes that aims to mature male gametes and occurs in the seminiferous tubules. In these processes, primitive spermatogonial stem cell (SSC) divides either to replace their number (stem cell renewal) or spermatogonia proliferate and then differentiate into spermatocytes; spermatocytes produce spermatids during mitotic division, and eventually, mature spermatids are released. Previous studies have shown that some factors are critical to the natural process of spermatogenesis. Therefore, various researches have been done to find suitable markers of germ cells. One of these markers is POU class 5 homeobox 1 (POU5F1) (Azizi et al., 2020a, 2021; Tabar et al., 2021).
POU5F1, also known as Oct4 (Octamer-binding transcription factor 4), expresses by primordial germ cells and SSCs, which is implicated in the regulation and maintenance of self-renewal (Zeineddine et al., 2014). SSCs are the most primitive and one of the rarest spermatogonia in the testis that play an essential role in maintaining spermatogenesis by self-renewal and the production of daughter cells to generate differentiated cells, spermatozoa, so they are very effective in transmitting genetic information to the next generation (Kubota and Brinster, 2018). It has been shown that these unipotent stem cells, which are located on the basal membrane in seminiferous tubules, can be cultivated on mouse embryonic fibroblast feeder layers in the presence of growth factors for long-term culture (Azizi et al., 2019b, 2020a, 2020b).
In our previous experiments, we have shown that derivation of embryonic stem cell-like (ES-like) cells from SSCs seems to be age-dependent (Azizi et al., 2016, 2019a). The conversion of adult SSCs into ES-like cells coincides with a reduction in expression of the important genes involved in the regulation of pluripotency such as POU5F1, Nanog, and Sox2 in SSCs with age (Azizi et al., 2016). POU5F1 is critical for reprogramming and pluripotency and it is a key transcription factor for the maintenance of the undifferentiated state of cells during early development and is a regulator of germ cell differentiation (Malik et al., 2019). Marti et al. (2013) have shown that POU5F1 has two isoforms POU5F1A and POU5F1B. Unlike POU5F1A, POU5F1B isoform is not related to stemness and pluripotency (Wang and Dai, 2010).
POU5F1B seems to be associated with the cellular stress response (Liu et al., 2017). The basic cellular localization of two isoforms POU5F1A and POU5F1B is different. POU5F1A is localized in the nucleus, while POU5F1B is mainly expressed in the cytoplasm (Cauffman et al., 2006).
This experiment aims to enhance our understanding of the POU5F1 expression levels, spatiotemporal dynamics, and its function in testicular germ cells, which can help to a more accurate understanding of fertility and the development of infertility treatments.
Materials and Methods
Digestion of testis
We divided 12 C57BL/6 mice into 4 age groups: 1 week, 2 weeks, 16 weeks, and 24 weeks and the cells of the testes isolated after treatment with enzymatic digestion solution contained collagenase IV (0.5 mg/mL; Sigma Aldrich), DNase (0.5mg/mL; Sigma Aldrich) and Dispase (0.5 mg/mL; Sigma Aldrich) at 37°C for 8 minutes. The digestion solution enzymatic reaction was paused with 10% fetal bovine serum (FBS; Invitrogen), and digested tissue slowly pipetted for obtaining suspension of single cells. The tissue samples of the digested testis were washed with Dulbecco's modified Eagle's medium/F12 (Invitrogen) and, after passing the cell suspension through a 70 μm mesh filter, centrifuged at 1500 rpm and it took 10 minutes (Azizi et al., 2016).
Culture of testicular cells
The neonate and adult testicular cell suspension was plated onto 0.2% gelatin-coated dishes in SSCs culture medium, which is included of StemPro-34 medium, 1% N2-supplement (Invitrogen), 6 mg/mL D+ glucose (Sigma Aldrich), 5 μg/mL bovine serum albumin (BSA; Sigma Aldrich), 1%
Gene expression analyses on the Fluidigm Biomark system
Dynamic Array chips (Fluidigm) were handled to measure the expression of the gene Pou5f1 (Mm03053917_g1), which was used for normalization of SSCs. In each individual sample, nearly 50 cells were manually elected from the various cells with a micromanipulator, lysed with special lysis buffer containing 9 μL RT-PreAmp Master Mix (5.0 μL Cells Direct 2 × Reaction Mix [Invitrogen], 2.5 μL 0.2 × assay pool, 0.2 μL RT/Taq Superscript III [Invitrogen], and 1.3 μL Tris EDTA (TE) buffer and directly frozen and stored at −80°C). The number of targeted transcripts was quantified using TaqMan real-time polymerase chain reaction (RT-PCR) on the Biomark Real-Time quantitative PCR (qPCR) system (Fluidigm). All samples were examined in two technical replicates. The Ct values achieved from the Biomark System were analyzed by GenEx software from MultiD analysis (Azizi et al., 2019a).
Immunocytochemical staining
Cells were cultured in 24-well plates and fixed with 4% paraformaldehyde. After washing with phosphate-buffered saline (PBS), the specimens were permeabilized with 0.1% Triton/PBS and stopped with 1% BSA/PBS. When blocking solution was removed, the cells were incubated overnight with primary antibody Abcam (ab19857), Santa Cruz (SC-8628) and StemGent (09-0023). After rinsing, the process was continued by incubation with species-specific secondary antibody, which was conjugated with fluorochrome; the marked cells were counterstained with 0.2 μg/mL DAPI (4′,6-diamidino-2-phenylindole) for 3 minutes at room temperature and fixed with Mowiol 4-88 reagent. Deletion of each primary antibody in the sample was used as a negative control for all markers. Labeled cells were examined with a confocal microscope Zeiss LSM 700, and images were photographed with a Zeiss LSM-TPMT camera (Azizi et al., 2020a).
Tissue processing for immunohistofluorescence staining
Testis tissue was removed from male animals, followed by washing in PBS and fixed in 4% paraformaldehyde for about 24 hours at room temperature. Dehydrated testis was embedded in Paraplast Plus. Testis tissue blocks were cut with a microtome at about 10 μm thickness. Parts were mounted on Superfrost Plus slides and kept at room temperature until usage.
All sections were deparaffinized with xylene and rehydrated in an ethanol series before staining. Afterward, heat-mediated antigen retrieval (10 mM sodium citrate buffer, pH 6 or 1 mM ethylenediaminetetraacetic acid, pH 8) at 95°C for 20 minutes, nonspecific binding was inhibited with 10% serum/0.3% Triton in PBS, and immunofluorescence staining went on as described above (Azizi et al., 2016, 2020a).
Protein–protein interaction network analysis
Search Tool for the Retrieval of Interacting Genes/Proteins (STRING version 11.5, https://string-db.org/) is a biological database and web resource of known and predicted protein–protein interactions (PPIs) (Lu et al., 2020). STRING app in Cytoscape Software version 3.8.2 was used to generate a PPI network. We used STRING: PubMed query as our data source to generate a network among the first 100 proteins involved in “testis development” in Mus musculus, and minimum confidence (score) cutoff was set 0.40. Then, the PPI network was constructed using Cytoscape Software. We identified the first neighbors' nodes with POU5F1 and created a new network with them.
Gene set enrichment analysis
To confirm the biological roles of the genes involved in the PPI network of first neighbors' nodes with POU5F1, we have performed functional gene enrichment analysis using the STRING Enrichment analysis in the Cytoscape. We reviewed a number of functional enrichments related to our laboratory data without considering a specific false discovery rate value.
Statistical analysis
The trials were replicated at least three times. The average genes expression in all groups was calculated, and the groups were evaluated using one-way analysis of variance (ANOVA), covered by the Tukey's post hoc tests. The expression of genes was compared with nonparametric Mann–Whitney's test. The variation between groups was considered statistically reliable, if a value of p < 0.05 had been acquired. PPI networks were analyzed based on relevant databases or online data analysis tools.
Ethical statement
In the current investigation, animal experiments were approved (Ir.ausmt.rec.1400.04) by the Ethics Committee of Amol University of Special Modern Technologies.
Results
POU5F1-GFP signals were compared in sections of seminiferous tubules of neonate mice (Fig. 1A1) and 16-week-old mice (Fig. 1B1), and the confocal scanning ultraviolet-laser microscope delineated that the expression of POU5F1 in neonate mice was much higher than 16-week-old mice (Fig. 1).
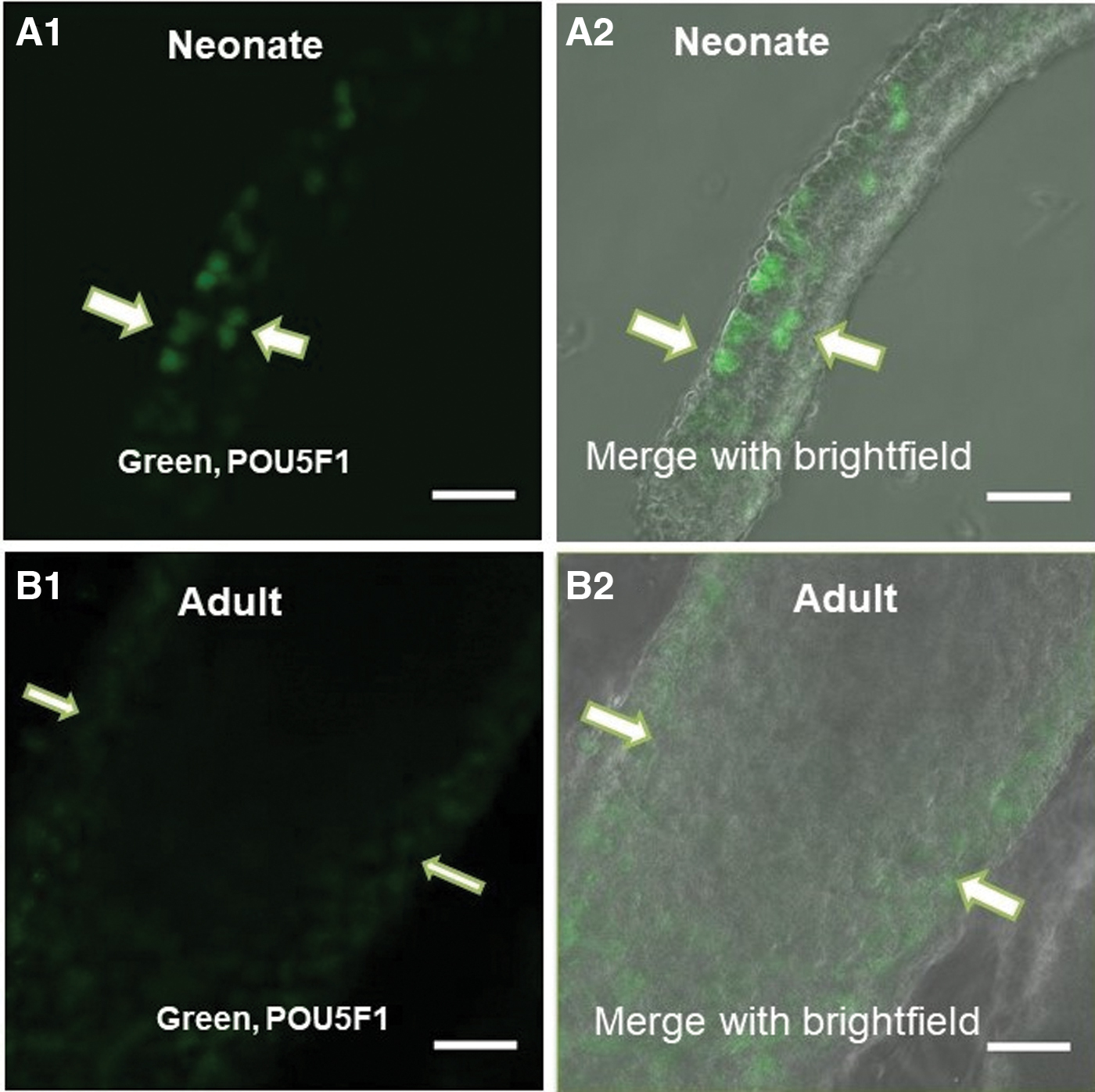
The comparison of expression of POU5F1 in Neonate and adult mouse testis in seminiferous tubules by confocal scanning UV-laser microscope. The high expression of POU5F1 in the seminiferous tubule of neonate mice
In the next step, by a confocal microscope, we analyzed POU5F1 protein localization in adult and neonate SSCs against POU5F1 protein from Abcam (ab19857), Santa Cruz (SC-8628), and StemGent (09-0023) by immunocytochemistry and immunohistochemistry (Figs. 2 and 3).
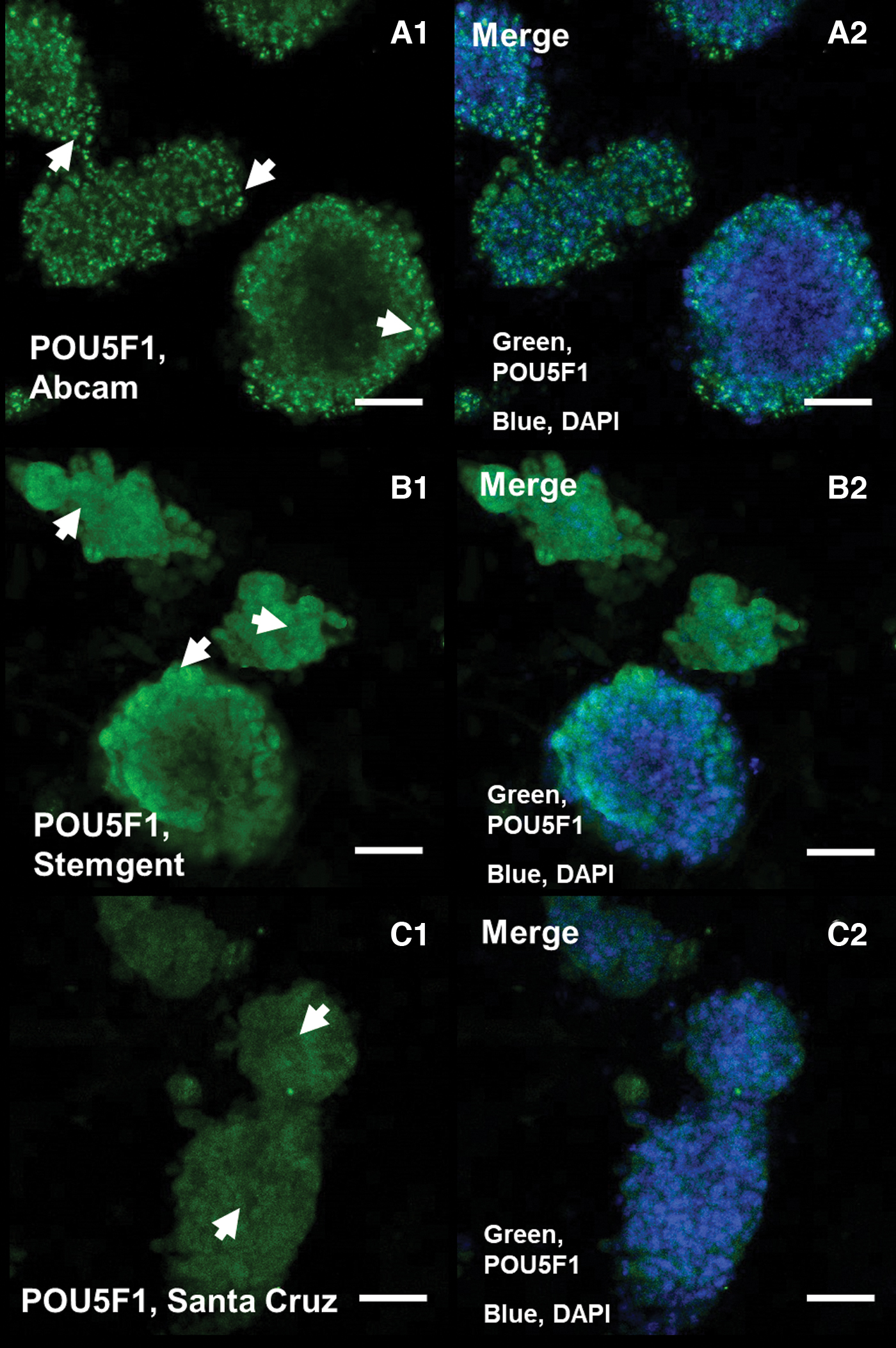
Immunocytochemical analysis of POU5F1 in adult SSCs by using of three antibodies (Abcam, Santa Cruz, and StemGent). POU5F1 high nuclear expression by ab19857
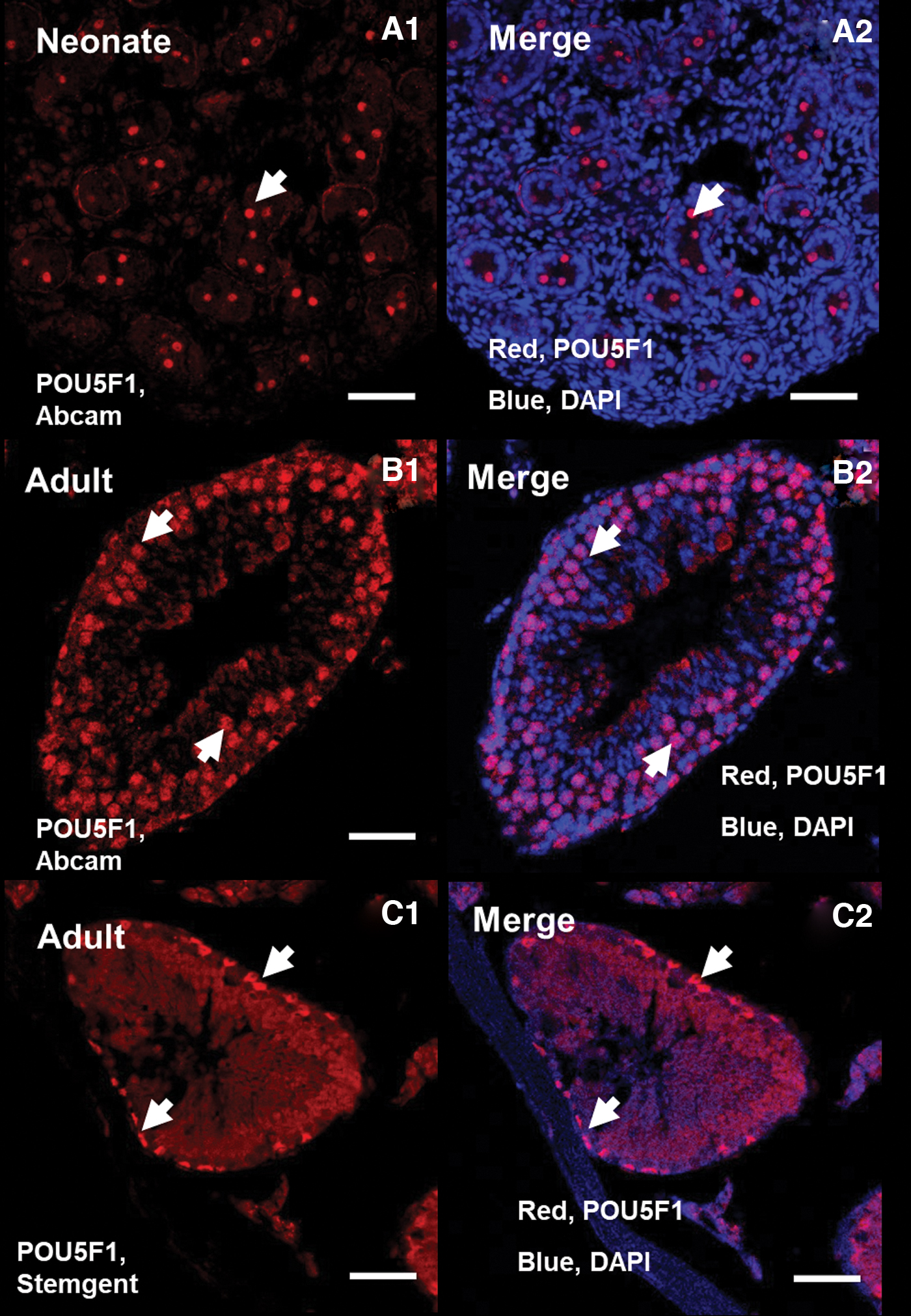
IHC analysis of POU5F1. The difference of expression of POU5F1 in adult and neonate testis by two color blending (Abcam and Stemgent). Nuclear and cytoplasm expression of POU5F1 from ab19857 in neonate
Immunocytochemistry analysis in adult SSCs confirmed high nuclear expression of POU5F1 protein by ab19857 (Fig. 2A1), while high cytoplasmic expression of POU5F1 was observed by 09-0023 in the SSCs (Fig. 2B1), and also low cytoplasmic POU5F1 expression was detected by SC-8628 (Fig. 2C1). Further, immunohistochemistry analysis by confocal microscopy shows both nuclear and cytoplasm expression of POU5F1 from ab19857 in adult and neonate testis and nuclear and cytoplasm expression of POU5F1 by 09-0023 in adult seminiferous tubule (Fig. 3).
Counting of POU5F1-positive cells from ab19857 in the seminiferous tubules of the neonate and adult testis displayed that almost 7% of the cells in the neonate and 27% of the cells in the adult testis were positive. Therefore, a higher number of POU5F1-positive cells were observed in the adult testis (Fig. 4).
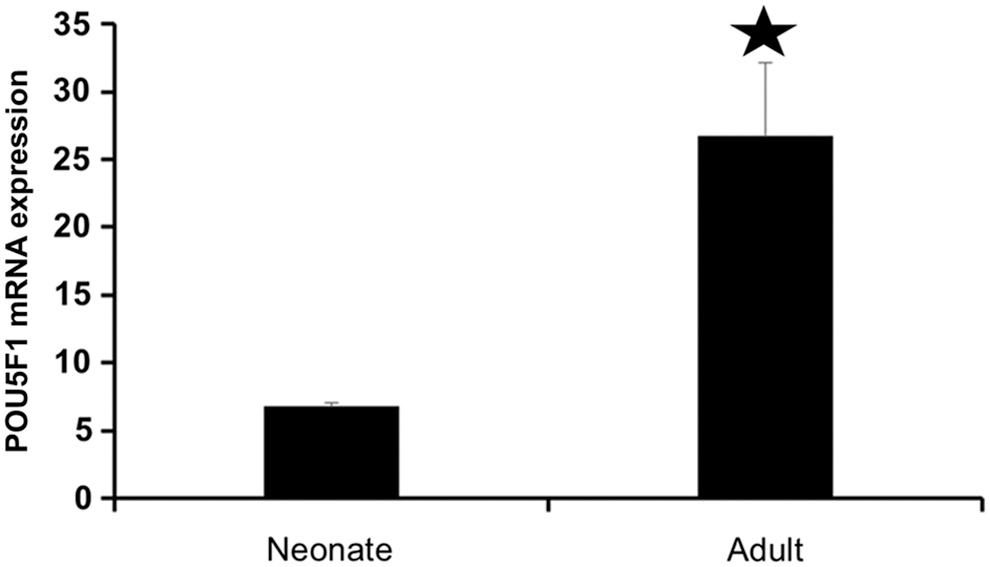
Counting polymerase chain reaction analysis for POU5F1 expression in neonate and adult mouse. Significant increase (*) in mRNA expression of POU5F1 in adult mouse (p < 0.05).
Fluidigm RT-PCR analysis indicated although neonatal germ cells significantly expressed POU5F1 than adult germ cells, the number of POU5F1-positive cells in adult testis is more than neonate testis sections, and also the expression of POU5F1 downregulated during differentiation of SSCs into sperm cells. qPCR analysis using single cells revealed expression of POU5F1 gene in neonate SSCs were significantly (p < 0.05) upper than 16- and 24-week SSCs (Fig. 5).
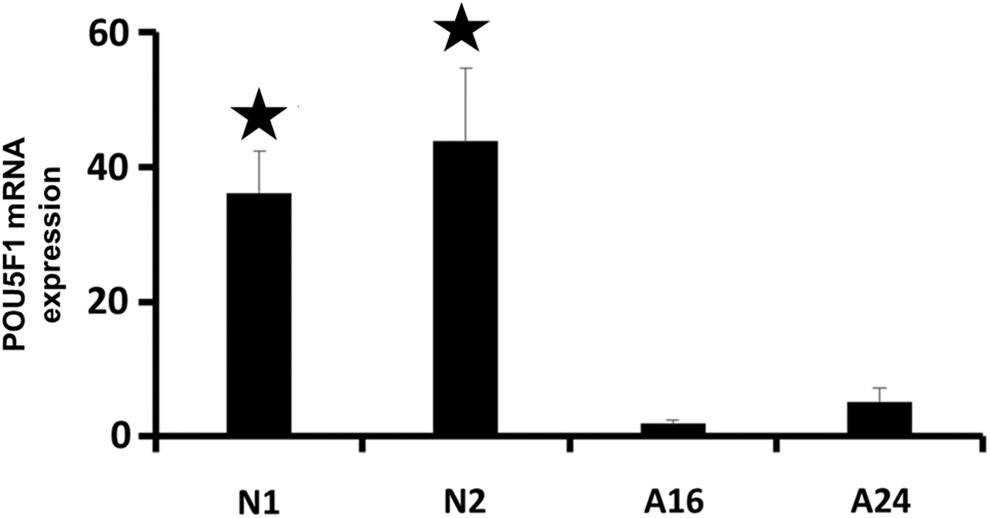
Fluidigm real-time RT-PCR analysis for POU5F1 expression. Determining the mRNA expression of POU5F1 in neonate (neonate 1 and 2 weeks) and adult SSCs (adult 16 and 24 weeks) showing significant difference (*) between neonates and adults (p < 0.05). RT-PCR, real-time polymerase chain reaction.
Furthermore, the PPI analysis was performed by STRING database to evaluate the physical and functional associations of proteins involved in “Testis development” (Fig. 6). Network nodes represent proteins and edges represent protein-protein associations (associations are meant to be specific and meaningful, that is, proteins jointly contribute to a shared function). According to the data obtained in the network, POU5F1 is also involved in this process and has many connections with other involved proteins. We used red for the POU5F1 node to make it easier to see (Fig. 6). In the next step, we identified the first neighbors' nodes with POU5F1, which includes 34 nodes, and highlight their interaction with POU5F1 (Fig. 7).
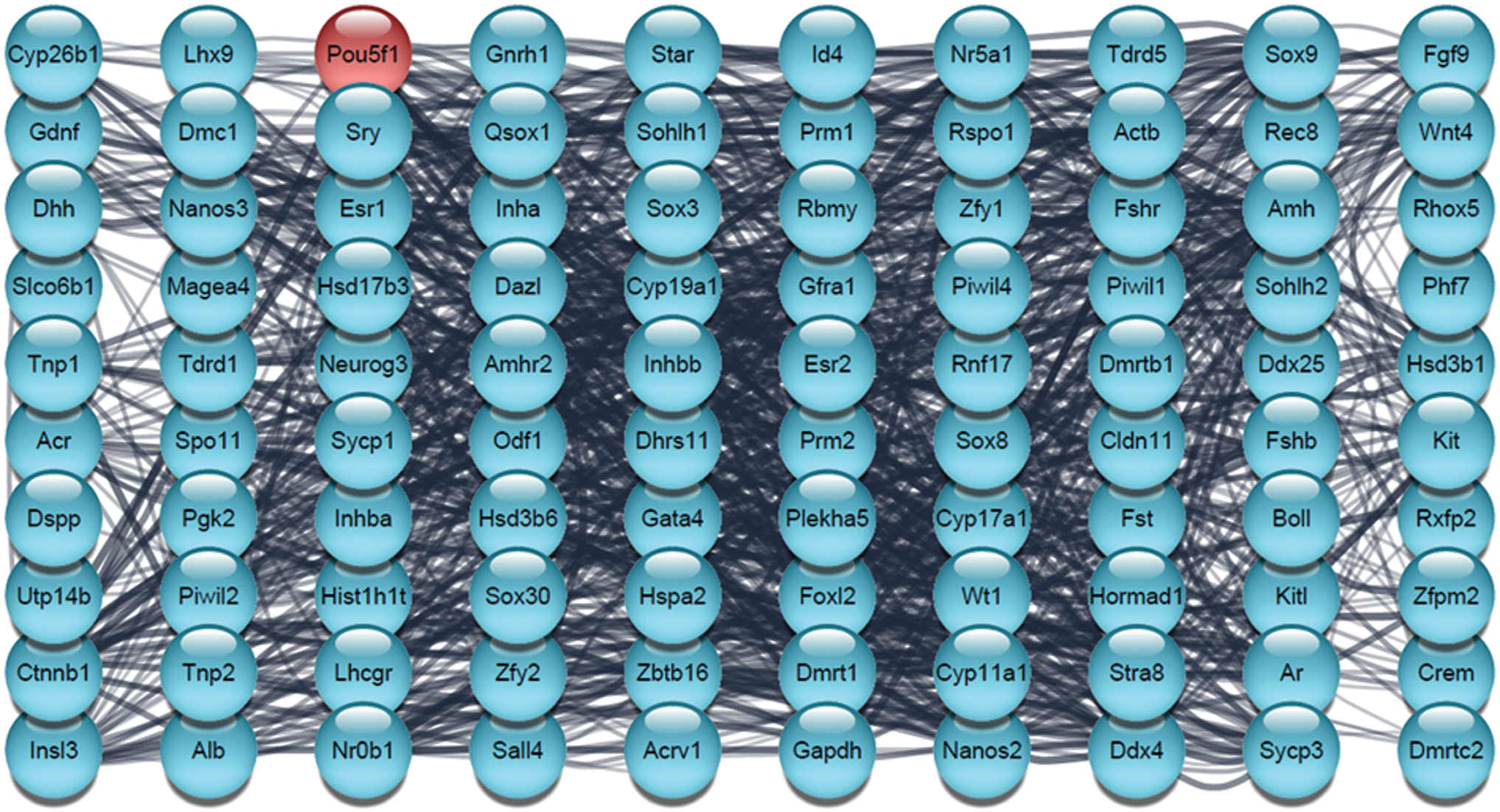
The PPI network analysis involved in testis development. The network was constructed by Cytoscape based on the PubMed query from the STRING database. The maximum number of proteins was set as 100 with confidence score cutoff >0.40. The red node represents POU5F1. PPI, protein–protein interaction. Color images are available online.

The interactions of the first neighbors' nodes of POU5F1. The red node represents POU5F1. Color images are available online.
Enrichment analysis was performed to identify enriched biological processes, molecular functions, and cellular locations associated with POU5F1 and its first neighbor proteins (Fig. 8). We selected some biological processes based on the purpose of our experiment, including germ-line stem cell population maintenance, growth, developmental growth, ectoderm development, blastocyst growth, cell differentiation, positive and negative regulation of developmental process, developmental induction, and tissue development. Furthermore by TISSUES analysis, we have studied the localization of POU5F1in spermatid, pluripotent stem cell, germ cell, blastocyst, and embryonic stem cell. Also, we used UniProt keywords to identify if the POU5F1 is a developmental protein (Fig. 8).
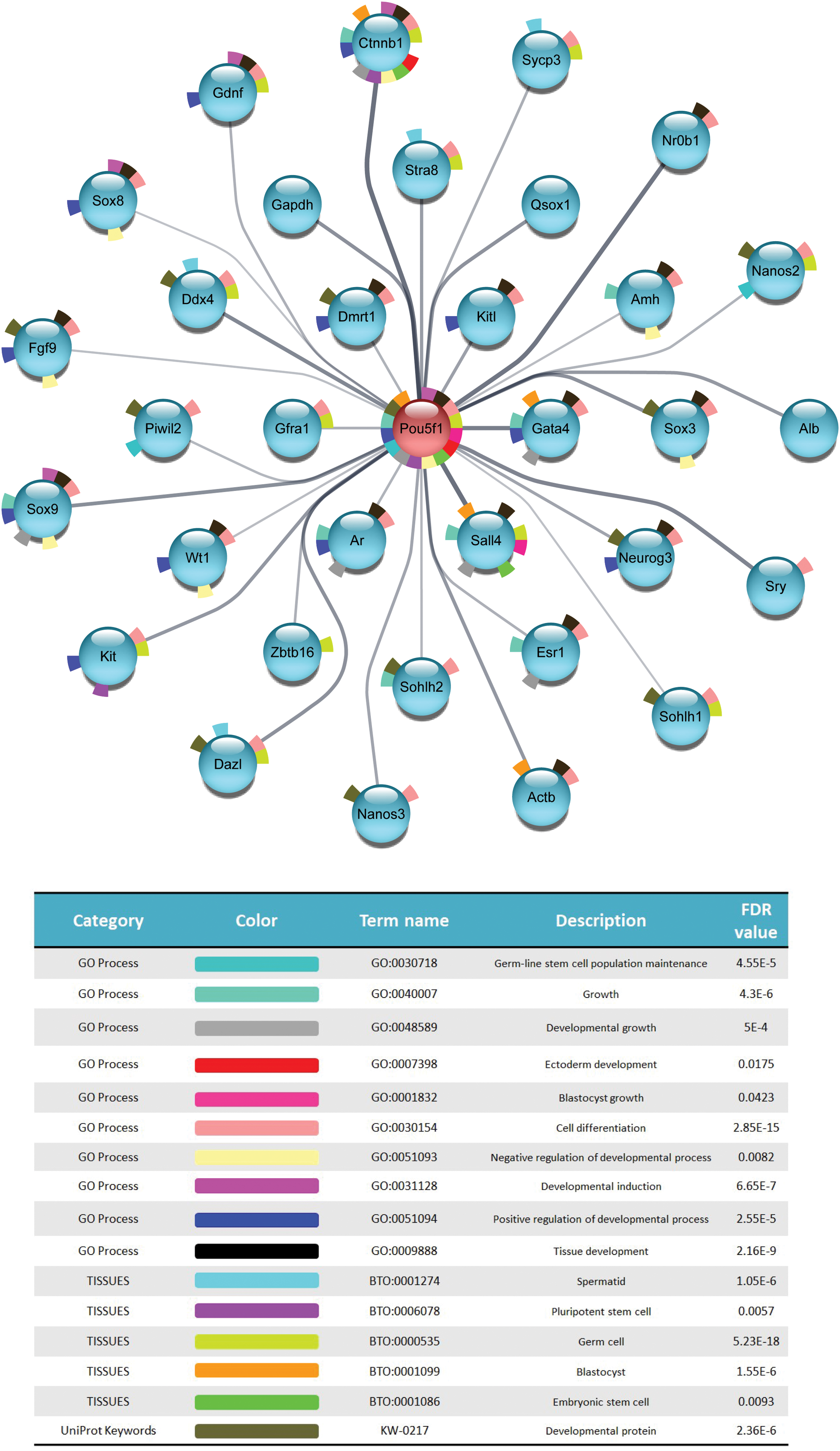
STRING Enrichment analysis. Each node represents a protein of our selected genes and edges indicate PPIs, with line thickness being indicative of evidence strength for a predicted interaction. Color images are available online.
Discussion
We provided a strong indication for the expression of POU5F1 in the seminiferous tubules of neonate and adult testis in the POU5F1-GFP transgenic mouse. Confocal laser scanning microscopy indicated the reduction of the signal density of POU5F1-GFP in the seminiferous tubules of the adult mouse than neonate. Zheng et al. (2016) have showed that the expression of POU5F1 has a chiefly nuclear localization in undifferentiated germ cells such as PGCs, gonocytes, and spermatogonia (Ohbo et al., 2003). Subsequently, Sohni et al. (2019) have showed that the decrease of POU5F1-GFP signal might be related to a reduction of gonocytes and primary undifferentiated Spermatogonial population in the testis of infants and older animals.
Also, according to the findings of Goel et al. (2008) the expression of pluripotent markers in the germ line are stage-specific and also they depend on the age of the species. This result was apparently confirmed by Fluidigm RT-PCR analysis, which demonstrated that expression of POU5F1 gene in neonate SSCs was higher than 16–24 weeks SSCs.
We kept the study of POU5F1 expression in testis portions by immunohistochemistry, it was distinguished that POU5F1 protein expression is in the basement membrane of the testis and decreases during spermatogenesis. It was also observed that in the neonate, unlike the adult, POU5F1 is expressed in the center of the seminiferous tubules. It seems that these germ cells were prospermatogonia, which capture at G0 mitotic stage (Payne, 2013; Yang et al., 2020). During early development, POU5F1 is expressed in pluripotent ES cells, blastomeres, and in the inner cell mass (ICM) of the blastocyst. Also after gastrulation, the POU5F1 is downregulated in trophectoderm and primitive endoderm but maintained in PGCs (Li, 2017; Pesce and Schöler, 2000). PGCs maintain the expression of POU5F1 until the initiation of the spermatogenesis in males (Pesce and Schöler, 2000).
Thus, the reduction of POU5F1 is an essential step in differentiation during the process of spermatogenesis (Li, 2017). Dann et al. (2008) have proven in their research that the POU5F1 expressed in undifferentiated SSCs is demanded for keeping of SSCs during in vitro culture and is also important for colonization activity following cell transplantation.
With anti-POU5F1 antibody from Abcam, Santa Cruz, and StemGent, we identify spatiotemporal dynamics of POU5F1 protein localization in SSCs and testis section. Researchers have done many studies on POU5F1, and in different articles, these three antibodies have been used the most. Hence, we decided to examine if there is any difference between these three. Immunohistochemistry analysis results demonstrated protein expression of the POU5F1 in the testis section with the Abcam and StemGent, but not from Santa Cruz antibody.
With alternative splicing and alternative translation, three mRNA isoforms of POU5F1 genes, including POU5F1A (OCT4A), POU5F1B (OCT4B), and POU5F1B1 (OCT4B1), and four protein isoforms, including POU5F1A, POU5F1B-190, POU5F1B-265, and POU5F1B-164, are present in the human cells (Mehravar et al., 2021). The isoforms of POU5F1 may exhibit different functional properties. The researchers examined the expression patterns between POU5F1A and POU5F1B in human ES cells and all stages of development before implantation by immunocytochemistry. It has been shown that POU5F1A was expressed in all compressed fetal nuclei and blastocysts, while OCT4B was expressed in the cytoplasm of all cells from the four-cell stage onward (Wang and Dai, 2010).
POU5F1A is a transcription factor that confirmed the stemness and pluripotency state of ES cells (Sneha et al., 2019). However, POU5F1B is mainly localized in the cytoplasm of nonpluripotent cells, which cannot maintain the self-renewal of ES cells and it is reduced in nonpluripotent stem cells, but may respond to cell stress (Liu et al., 2017). During coexpression of POU5F1A and POU5F1B, activation of transcriptions by POU5F1A was not repressed by POU5F1B and parallel overexpression of POU5F1B was not activated by POU5F1A (Wang and Dai, 2010). Guo et al. (2012) have identified two variants of the POU5F1 gene, including POU5F1A and POU5F1B in mouse cells. POU5F1A exists only in one variant, which is the basic regulator of pluripotent stem cells, while three isoforms of POU5F1B include POU5F1B-247aa, POU5F1B-190aa, and POU5F1B-164aa and were found in somatic cells of the mouse (Guo et al., 2012).
Warthemann et al. (2012) demonstrated that POU5F1 from Abcam (ab19857) is common to both POU5F1A and POU5F1B, while the Santa Cruz antibody (Sc-5279) is directed against POU5F1A. The confirmation of this result is downregulation of POU5F1 expression during spermatogenesis. This could be related to the production of different differentiation factors by Sertoli cells in the two different compartments surrounded and separated by these cells, the stem cell and the differentiation compartment (Huleihel and Lunenfeld, 2004).
In addition, the data obtained from the network analysis show a strong association of POU5F1 with other genes involved in the process of testicular development. It has been shown that this gene plays an important role in the regulation of the developmental processes in different embryonic stages such as blastocyst and ectoderm. It is also present in cell lines such as embryonic stem cells, germ cells, blastocyst, and pluripotent stem cells in general, but not in spermatids. This result may explain why POU5F1 decreases in the differentiation process during spermatogenesis. Hambiliki et al. (2012) have shown by immunostaining that POU5F1 is expressed in the blastocyst stage and is more expressed in the ICM than trophectoderm.
On the contrary, Takahashi and Yamanaka (2006) have proved that POU5F1 is one of the key factors in maintaining pluripotency in stem cells and has been used to induce pluripotency in the somatic cell line. High levels of POU5F1 in the ICM as well as embryonic stem cells are necessary to maintain pluripotency, so the level of this factor is tightly linked to cell fate (Dann et al., 2008). Also, Li et al. (2015) have shown in their experiments that after birth, the expression of POU5F1 is limited only to undifferentiated spermatogonia in the stages before puberty. Yoshimizu et al. (1999) have stated that POU5F1 plays a crucial role in the development of germ cells because it is specifically expressed in most stages of the germ cell line development.
In addition, Vansandt et al. (2016) have shown that POU5F1 expression is age-dependent. Zeng et al. (2016) using high-throughput RNA-Seq have shown that the expression of POU5F1 and NANOG was required in germ cells development and they control multiple downstream transcription factors and epigenetic modifiers that maintain pluripotency of stem cells. Overall, according to the data obtained, POU5F1 is one of the developmental proteins that induce development.
Conclusion
Our approach provides POU5F1 protein localization, which is highly expressed in both neonatal and adult testicular niches, and also in isolated SSCs from testicular cultures. While high POU5F1 gene and protein expression was observed in the neonate testis, the number of POU5F1-positive cells in the seminiferous tubules of adult testis was greater than neonate. In the future, it would be very important to understand why POU5F1 expression is downregulated during the differentiation of SSCs into sperm. This new result provides several important milestones that should be overcome for in vitro differentiation of stem cells into germ stem cells and for infertility treatment.
Footnotes
Authors' Contributions
Z.N.N.: wrote the article and the data analysis. H.A.: carried out and designed the experiment, assembly of data, and data analysis. K.S.: designed PPI network and enrichment analysis, wrote the article and data analysis, and edited the final document. The authors read and approved the final article.
Acknowledgments
The present article extracted from master dissertation of microbial biotechnology at Amol University of Special Modern Technologies, Iran in 2021. I first thank my supervisor, Dr. Hossein Azizi whose expertise was invaluable in formulating the research questions and methodology. His insightful feedback pushed me to sharpen my thinking and brought my work to a higher level. I also acknowledge my colleagues for their wonderful collaboration.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
No funding was received for this article.