
Abstract
Zinc is an essential trace element for bone growth and bone homeostasis in the human body. Bone mesenchymal stem cells (BMSCs) are multipotent progenitors existing in the bone marrow stroma with the capability of differentiating along multiple lineage pathways. Zinc plays a paramount role in BMSCs, which can be spurred differentiating into osteoblasts, chondrocytes, or adipocytes, and modulates the formation and activity of osteoclasts. The expression of related genes also changed during the differentiation of various cell phenotypes. Based on the important role of zinc in BMSC differentiation, using zinc as a therapeutic approach for bone remodeling will be a promising method. This review explores the role of zinc ion in the differentiation of BMSCs into various cell phenotypes and outlines the existing research on their molecular mechanism.
Introduction
As one of the essential trace elements for the human body, zinc plays a pivotal part in crucial physiological processes, such as individual growth, development (Gao et al., 2015), reproductive inheritance (Nasiadek et al., 2020), the maintenance of immune function (Read et al., 2019), and endocrine (Baltaci et al., 2019). It is also a component of above 50 metalloenzymes involved in enzymatic catalysis, cell growth, metabolic differentiation, intracellular signal transduction, and regulation of nucleic acid structure (Narayanan et al., 2020). At the molecular level, zinc also acts a vital role in the synthesis of proteins, receptors, and hormones (Barroso et al., 2018).
Moreover, over the last two decades, researchers have done a lot of research on zinc, which acts a pivotal part in normal bone growth and bone homeostasis. Qi et al. (2020) harbored the idea by in vivo studies that diabetic rats with zinc supplementation present increased bone mineral density, improved bone tissue morphology, and decreased bone loss. Accordingly, nutritional zinc can prevent the occurrence of osteoporosis because of its stimulation function for skeleton formation (Yamaguchi, 2010). Furthermore, zinc possesses insulin properties, which could have positive effects on bone remodeling and spinal fusion (Koerner et al., 2016). It is noteworthy that the entry of zinc ions into cells to perform bone formation functions is tightly controlled by cell receptors, including different zinc transporters (ZnTs), osmotic channels, and metallothioneins (Fukada et al., 2011).
Among the process, membrane ZnT proteins include the Zrt- and Irt-like protein (ZIP) family with 14 mammalian members and the ZnT family with 10 mammalian members (Huang et al., 2020), which positively regulate the movement of Zn2+ into cytoplasm and transport of Zn2+ out of the cytoplasm, respectively (Zhu et al., 2017). Zinc can affect a host of signal transduction pathways, including nuclear factor kappaB (NF-κB) (Haddad, 2009; Jarosz et al., 2017), AKT (Vahidi Ferdowsi et al., 2020), insulin/IGF-1 (Kumar et al., 2016; Norouzi et al., 2018; Novakovic et al., 2020), GM-CSF (Aster et al., 2019), cGMP, and PKC (Beyersmann and Haase, 2001). All in all, the physiological amount of zinc mainly promotes growth through increasing bone mass and suppressing bone resorption.
Mesenchymal stem cells (MSCs) are a type of pluripotent stem cells, which can be shown everywhere in the body and possess features of self-renewal ability (Friedenstein et al., 1976; Pittenger et al., 1999). They can be separated from bone marrow (Friedenstein et al., 1976), adipose tissue (Fraser et al., 2006), peripheral blood (Cao et al., 2005), the dental pulp (Perry et al., 2008), the lung (Gong et al., 2014), and placenta, amnion, umbilical cord, and cord blood (Hass et al., 2011). Since bone marrow is its main source, they are collectively called bone mesenchymal stem cells (BMSCs). At present, we often designate BMSCs as the gold standard, which is commonly still the most examined stem cell type (Friedenstein et al., 1976).
Numerous reports demonstrate that BMSCs are differentiated into various cell phenotypes containing chondrocytes, osteoblasts, and adipocytes (Oryan et al., 2017; Richardson et al., 2016). Research in vivo and ex vivo has shown that BMSCs enjoy profound potential in bone restoration and bony reconstitution.
However, the mechanism of BMSC treatment in bone repair has not been fully elucidated (Fu et al., 2019). Besides, BMSCs also act crucial roles in articular cartilage repair. For instance, recently, Liu et al. (2020) confirmed that BMSCs, modified by SUMO (small ubiquitin-like modifier), rehabilitate articular cartilage in rat. Interestingly, in vivo and in vitro investigations have shown that hypoxic culture and appropriate mechanical stimulation enhance chondrogenic differentiation and formation of BMSCs (Bornes et al., 2015, 2018; Xie et al., 2019). In addition, BMSCs can differentiate into chondrocytes during osteoporosis.
The adipogenic differentiation and osteogenic differentiation of BMSCs antagonize each other (Chen et al., 2016). Further research on its molecular mechanism manifested that the GDF11-FTO-Pparg axis promotes the transformation of osteoporotic BMSCs into adipocytes and suppressed bone formation (Shen et al., 2018). Moreover, further study has found that BMSCs are differentiated into adipocytes rather than osteoblasts during osteoporosis, which partly gives rise to osteoporosis. This perhaps provides new methods of osteoporosis therapy (Hu et al., 2018).
The skeleton of zinc accounts for a large proportion of the body's total load, which is crucial to the growth and development of healthy bones (Herzberg et al., 1990). Zinc is distributed to intracellular organelles after being taken up into BMSCs (Hojyo and Fukada, 2016). The endoplasmic reticulum is a mobilizable zinc store (Adulcikas et al., 2019). Zinc as a cofactor of numerous enzymes and transcription factors regulates cellular enzyme activity (Bellomo et al., 2014; Vallee and Auld, 1995). The intracellular zinc level is regulated by Zn transporters; its' coordinated activity directly affects the absorption and maintenance of Zn (Hojyo and Fukada, 2016).
A report indicated that ZIP1, located in the cytoplasmic membrane, induces osteogenesis of MSCs by promoting the zinc influx (Tang et al., 2006). ZnT7 can impair the osteogenesis of MSCs and prevent osteoblasts from apoptosis (Liang et al., 2013; Liu et al., 2013). The underlying function of ZnTs in the roles of zinc ion on BMSCs requires to be further studied.
In the last two decades, much of the research in containing zinc ion biomaterials has been examined. The multiplication and differentiation of BMSCs are significantly increased after coating Zn2+ on scaffolds (Amiri et al., 2016; Heras et al., 2019). In vitro, it was found that there is high alkaline phosphatase (ALP) activity, biological mineralization, and expression of osteogenic specific genes in zinc-induced osteogenic differentiation of BMSCs. Interestingly, the biomaterials can also provoke angiogenesis and effectively inhibit bacteria (Telgerd et al., 2019; Zhou et al., 2020), which can better result in BMSCs differentiating into osteogenic (Borys-Wojcik et al., 2019). These advantages will tremendously augment the survival rate of transplanted BMSCs and improve their potential for tissue regeneration (Zhang et al., 2011).
It is worth noting that zinc oxide nanoparticles may induce apoptosis of BMSCs by mediating cyclooxygenase and lipoxygenase (Kim et al., 2019). Besides stimulating osteogenic differentiation, zinc ion effectively inhibits osteoclastic and adipocytic differentiation of BMSCs to promote bone mass (Li et al., 2015). Furthermore, ZnO-containing composite scaffolds promote osteochondral differentiation of BMSCs due to zinc possesses' well-known insulin-mimetic properties (Khader and Arinzeh, 2020). According to recent studies, the role of aging on the differentiation of MSCs is very important.
Aging is one of the most significant elements in the proliferation and differentiation capacity of MSCs. Its molecular mechanisms are associated with progressive telomere shortening (Wright and Shay, 2005). The element zinc acting as an antioxidant or anti-inflammatory is crucial to prevent cellular aging (Prasad, 2014). Farahzadi et al. (2017) found in vitro that ZnSO4 promotes the lengthening of telomeres by augmenting the telomere-related gene expression and telomerase activity, and changing the hTERT promoter methylation status of human adipose-derived MSCs. Furthermore, studies in vivo showed that zinc sulfate significantly enhances the telomerase gene expression and decreases the aging of rat adipose tissue-derived MSCs under exposure to electromagnetic fields (Fathi and Farahzadi, 2018; Fathi et al., 2017). However, the application of aged MSCs remains to be explored further.
Therefore, in this review, we summarized the role of zinc ion on MSC differentiation, which can provide a theory for researchers to better use zinc as a therapeutic approach for bone remodeling. In addition, the review will be the first summary to combine the molecular mechanism and biomaterial role of zinc on BMSC differentiation; it could potentially be beneficial for several clinical osteopathic of zinc supplement with BMSCs.
The Effect of Zinc-Containing Biomaterials on BMSCs
The effect of zinc ion on BMSC behavior is investigated by means of several biomaterials such as bioactive glass, nanofiber scaffolds, coatings, and alloys. Multitudes of researches demonstrated that zinc-containing biomaterials play a significant role in the viability, proliferation, and differentiation of BMSCs. For instance, Hu et al. (2012) confirmed in vitro that the adhesion, proliferation, and differentiation of BMSCs are significantly augmented by zinc-doped coating compared with those without zinc. Amiri et al. (2016) demonstrated that the Zn2SiO4-coated fibrous scaffold significantly enhanced the proliferation and osteogenic differentiation of BMSCs, ALP activity, and biomineralization. And the expression of osteoblast-specific genes and proteins is significantly enhanced during differentiation of BMSCs. Moreover, zinc-containing biomaterials also possess antibacterial, anti-inflammatory, and angiogenic properties.
Hu et al. (2012) found that the growth of Staphylococcus aureus and Escherichia coli was obviously inhibited on the Zn-doped TiO2 coating, and the bacteriostatic ability of the coating gradually improved with the increase of zinc content in the coating. An experiment in vivo by establishing the rabbit bone defect model reported that the graphene oxide foam/polydimethylsiloxane/zinc silicate (ZS) scaffold group showed a significant new bone and bone trabecular formation in comparison to the group without zinc at 12 weeks postimplantation, and no infection was observed at the graft site (Li et al., 2020). These properties make zinc-containing biomaterials candidates for orthopedic and dental implants.
The first stage of BMSCs on the surface of zinc-containing biomaterials involved in osseointegration is their adhesion and diffusion (Mendonça et al., 2008). The stage is extremely critical because the quality of the stage directly affects the ability of BMSCs to proliferate and differentiate when in contact with the zinc-containing biomaterials (Anselme, 2000). A multitude of the literature suggests that the zinc-containing biomaterials are conducive to the adhesion and diffusion of BMSCs (Dhand et al., 2016; Hu et al., 2012; Zhang et al., 2018). Jin et al. (2014) advocated that zinc ion is the main reason for the abnormal stretching behavior of BMSCs in vitro, which facilitates cell proliferation.
Then Yu et al. (2017b) further in vitro demonstrated that the surface of Zn/Mg-PIII promotes the initial adhesion and spread of BMSCs by upregulating the integrin α1/β1 gene expression. Afterward, it was found by Maimaiti et al. (2020) that during the early stage of osteogenesis, the active sites on the surface of ZnO-NPs promote protein interactions, and then increase the activity of osteoblasts. Recent studies by Chopra et al. (2020) suggested that the adhesion of BMSCs to the zinc-containing rGO plates may be due to their favorable adsorption of proteins. In addition, the adhesion, proliferation, and differentiation activities of BMSCs are influenced by the surface properties of the biomaterial, including surface chemistry, surface morphology, and other properties (Burg et al., 2000). Zinc ion and biomaterials have a synergistic effect on BMSC adhesion.
The next stage is the proliferation and differentiation of BMSCs (Zhu et al., 2018). Zinc has a considerable protective effect on the transplantation of BMSCs in vivo. For instance, BMSCs have higher survivability on nanocoated surfaces containing zinc ion compared with without zinc ion. After being cultured on HAP-ZAg and HAPZAg-Cu surfaces for 28 days, the cell survival rate was higher than 95%. Conversely, the cell survival rate was less than 70% on day 28 on HAP-COM (Bostancioglu et al., 2017).
Aaron et al. confirmed that the faster degrading ZSr41A and ZSr41B alloys showed a dramatically lower number of viable BMSCs adhered to their surfaces (Cipriano et al., 2017). It has been shown that the number of BMSCs that survived decreases with the faster degradation rate of zinc ion in biomaterials. However, the supporting mechanism has not been explored in detail. It was found by Wang et al. (2019) that zinc gluconate and Zn2+-passivated carbon dots promoted the survival of BMSCs when zinc ion was at a certain solubility. Moreover, the adhesion of BMSCs on the surface of the new zinc-doped cement increased with the increase of zinc ion content in vitro (Cama et al., 2017).
In addition, supplementation with physiologic amounts of zinc gives rise to marked attenuation of cell apoptosis, enhanced cell viabilities, and increased vascular endothelial growth factor (VEGF) release, which may improve the survival rate of transplanted BMSCs (Zhang et al., 2011). Therefore, zinc supplementation prevents BMSCs from apoptosis, increases the survival rate of BMSC transplantation, and improves its tissue regeneration potential. Moreover, it further promotes bone formation in the body and accelerates bone healing. All in all, BMSCs treated with zinc have a significant effect on promoting bone healing.
Zinc Effects on the Osteoblast Differentiation of BMSCs
Zinc can promote osteoblast formation
Based on the paramount role of zinc in bone development and growth, considerable literature has suggested that zinc ion can promote BMSCs to osteogenic formation. A shared finding was that zinc promotes the ALP activity, proliferation, and calcific deposition of BMSCs in vitro (Cama et al., 2017; Fernandes et al., 2020; Oh et al., 2011; Zhang et al., 2012; Zhu et al., 2017) and in vivo (Hadley et al., 2010; Hie et al., 2011). The second shared finding was that zinc supplementation augments the expression of osteogenic specific genes and proteins in differentiated BMSCs, such as Runx2, ALP, type 1 collagen (Col-I), osteocalcin (OCN), and osteopontin (OPN) (Amiri et al., 2016; Chopra et al., 2020).
The third shared finding was that the effects of zinc on osteogenic differentiation processes show a time and dose-response. The differentiation process of MSCs can be divided into three stages: proliferation, maturation, and mineralization. During this process, a variety of genes are upregulated/downregulated according to different stages of differentiation (Liao et al., 2016). In the early stage of osteogenesis of BMSCs, zinc increases the expression of several osteogenic specific genes in vitro, such as ALP, Runx2, and Col-I (Fernandes et al., 2020; Qiu et al., 2018; Zhang et al., 2012).
During the middle and late differentiation stage, it has been shown that OCN and OPN significantly enhance zinc-stimulated osteogenic differentiation of BMSCs (Table 1) (Oh et al., 2012; Qiu et al., 2018; Zhang et al., 2012). For instance, Chopra et al. (2020) demonstrated that the expression of ALP, Col-I, and BMP-2 significantly increased range from 7 to 14 days, while being downregulated at 21 days. Moreover, the expression of OCN and OPN at 21 days significantly increased compared with 7 and 14 days. In addition, zinc ion concentration also has a biphasic reaction on osteogenic differentiation of BMSCs.
The Role and Molecular Mechanism of Zinc During Bone Mesenchymal Stem Cell Differentiation
+, upregulate; −, downregulate; /, none; ALP, alkaline phosphatase; Col-I, type 1 collagen; ERK, extracellular signal-regulated protein kinases; MAPK, mitogen-activated protein kinase; NF-κB, nuclear factor kappaB; OCN, osteocalcin; OPG, osteoprotegerin; OPN, osteopontin; RANKL, nuclear factor receptor activator-κB ligand; Runx2, runt-related transcription factor 2; TGF-β, transforming growth factor beta 1; TRAP, tartrate-resistant acid phosphatase.
Xiong et al. (2019) confirmed by ex vivo cell culture experiments that the presence of 10.91–27.15 μM of Zn ion promoted the upregulation of the osteogenesis-related gene expression; doses above 128.58 μM significantly reduced the ALP activity of BMSCs. The Zn concentration exceeding 130 μM was cytotoxic on the proliferation of MC3T3-E1 cells (Liang et al., 2012), which might be associated with the rapid release of zinc ion (Ishikawa et al., 2002; Ito et al., 2000). Thus, whether and how the release rate and dose of zinc play a synergistic role in promoting osteogenic differentiation of BMSCs and the release rate can replace the concentration of zinc ion in osteogenic differentiation remain to be elucidated.
Numerous studies found that Runx2 expression is significantly increased after adding zinc to BMSC culture (Chandramohan et al., 2021; Fernandes et al., 2020; Yusa et al., 2016). Runx2 is the early major transcription factor for osteoblastogenesis (Hayrapetyan et al., 2015), which could positively regulate the proliferation, differentiation, and expression of osteoblast genes (Fig. 1) (Jensen et al., 2010). Moreover, Runx2 also is responsible for the activation of several osteoblast-specific genes such as ALP, Col-I, OCN, and OPN to promote osteoblast maturation (Grafe et al., 2018; Kim et al., 2020).

Zinc promotes the osteogenic differentiation of BMSCs. Runx-2, as the earliest transcription factor for osteogenic differentiation, can activate the expression of multiple late-stage osteogenic-related genes. Increased expression of Runx-2 can promote the differentiation of BMSCs into precursor chondrocytes. And the increased expression of Osterix can promote the maturation of subsequent osteoblast development. BMSC, bone mesenchymal stem cell.
Similarly, a study found that the expression of ALP, OPN, and OCN genes is not present in Runx2 gene knockout mice (Komori et al., 1997). Runx2 also can promote the proliferation and differentiation of osteoblast-lineage cells by activating multiple cellular signaling pathways. Komori (2020) confirmed that Hedgehog, Fgf, Wnt, and Pthlh signaling pathways stimulate Runx2 gene expression, promoting the proliferation and osteogenic differentiation of BMSCs; Runx2 interacts with these signaling pathways to control osteogenic differentiation, rather than their cascade. However, the transcriptional regulation of the Runx2 gene in osteogenic differentiation needs to be elucidated.
Moreover, the addition of zinc could significantly promote the expression of ALP, Col-I, CON, and OPN on BMSC culture. Hu et al. (2012) showed that ALP, as an early osteoclastogenic gene, participates in the premineralization process and promotes the formation of mineral nodules. Gang et al. (2006) note that Col-Iα has a handsome effect on the early deposition of inorganic molecules and by mediating integrin β1 to promote adhesion behavior. Qian et al. (2015) observed that the upregulation of OCN leads to the deposition of minerals in the extracellular matrix and new bone mineralization. These osteogenic genes play vital roles in various stages of osteogenic differentiation of BMSCs. However, the transcriptional regulation of these genes has not been fully understood.
Of particular note, physiologic amounts of zinc also inhibit BMSC apoptosis. Zhang et al. (2011) showed by establishing a mouse ischemic hindlimb model that replenishment of physiologic quantity of zinc markedly attenuates BMSC apoptosis and improves their viabilities by activating Akt. Kim et al. (2019) found that zinc significantly attenuates ZnO NP-induced toxicity and apoptosis in human BMSC cultures. Apoptosis is associated with the increased production of reactive oxygen species and the activation of caspase-3/7, which participate in mitochondrial dysfunction (Kim et al., 2019; Orazizadeh et al., 2015; Syama et al., 2014).
Moreover, Liang et al. confirmed that zinc would protect osteoblast cells from apoptosis and oxidative damage by mitogen-activated protein kinase (MAPK) and PI3K/AKT systems. Interestingly, zinc deficiency could induce preosteoblasts MC3T3-E1 cells apoptosis through a mitochondrial-mediated pathway (Guo et al., 2012). Zinc could protect osteoblasts from apoptosis (O'Connor et al., 2020). Thus, whether zinc protects osteogenic differentiation of BMSCs from apoptosis requires further exploration in detail.
Zinc-stimulated osteoblast differentiation molecular mechanisms
As mentioned above, zinc promotes the osteogenic differentiation of BMSCs, and several cellular special genes upregulate/downregulate in related signaling pathways during the process. A considerable number of research in vivo and ex vivo have investigated the mechanisms of osteoblastogenic effects induced by zinc. According to numerous literatures, zinc may promote osteoblastogenic differentiation of BMSCs through various cellular molecular pathways, such as MAPK pathway, Wnt pathway, BMP/transforming growth factor beta 1 (TGF-β) pathway, and cAMP pathway.
MAPK signal transduction is considered to be one of key signal pathways in the process of osteogenic differentiation, which includes extracellular signal-regulated protein kinases (ERK), p38, c-Jun N-terminal kinase (c-JNK), and ERK5 (Hipskind and Bilbe, 1998; Schneider et al., 2001; Seger and Krebs, 1995). The activated kinase would activate the downstream genes by translocating to the nucleus, triggering the activation of Runx2, and thus altering the differentiation of BMSCs (Ge et al., 2007).
Although the elegant work by Matsuguchi et al. (2009) strongly suggests that MAPK/JNK plays a critical role in the process of osteoblast differentiation, the support mechanism is by far the least understood. In addition, Gao et al. (2021) confirmed in vivo that the MAPK/ERK cascade is significantly activated at the transcriptional level, suggesting the MAPK/ERK pathway might be more dominant in the osteogenic differentiation of BMSCs than other MAPK pathways. Kawashima et al. (2020) noted that surface reaction-type prereacted glass ionomer (S-PRG) could be internalized into osteoblastic cells, thereby activating MAPK pathways, mainly the ERK and p38 pathway, thus inducing osteogenic differentiation and mineralization.
Kang et al. (2018) showed that zinc/aluminum-layered double hydroxides (Zn-Al LDH) upmodulated osteoblastic classical marker genes such as Runx2 and Osterix, thus promoting osteoblast differentiation by activating JNK and ERK pathways. In cell culture, most MAPK/ERK signal through Ras, Raf, MEK, and ERK1/2 to promote osteoblast differentiation (Fig. 2). A study has found that extracellular Zn2+ enters the BMSCs by endocytosis, after internalization, activating the ERK pathway and eventually promoting osteogenesis (Gao et al., 2021). Cation diffusion facilitator protein is functionally similar to vertebrate ZnT1, which controls the Ras–Raf–MEK–ERK pathway by promoting Zn2+ efflux and reducing the concentration of cytosolic zinc ion (Bruinsma et al., 2002).
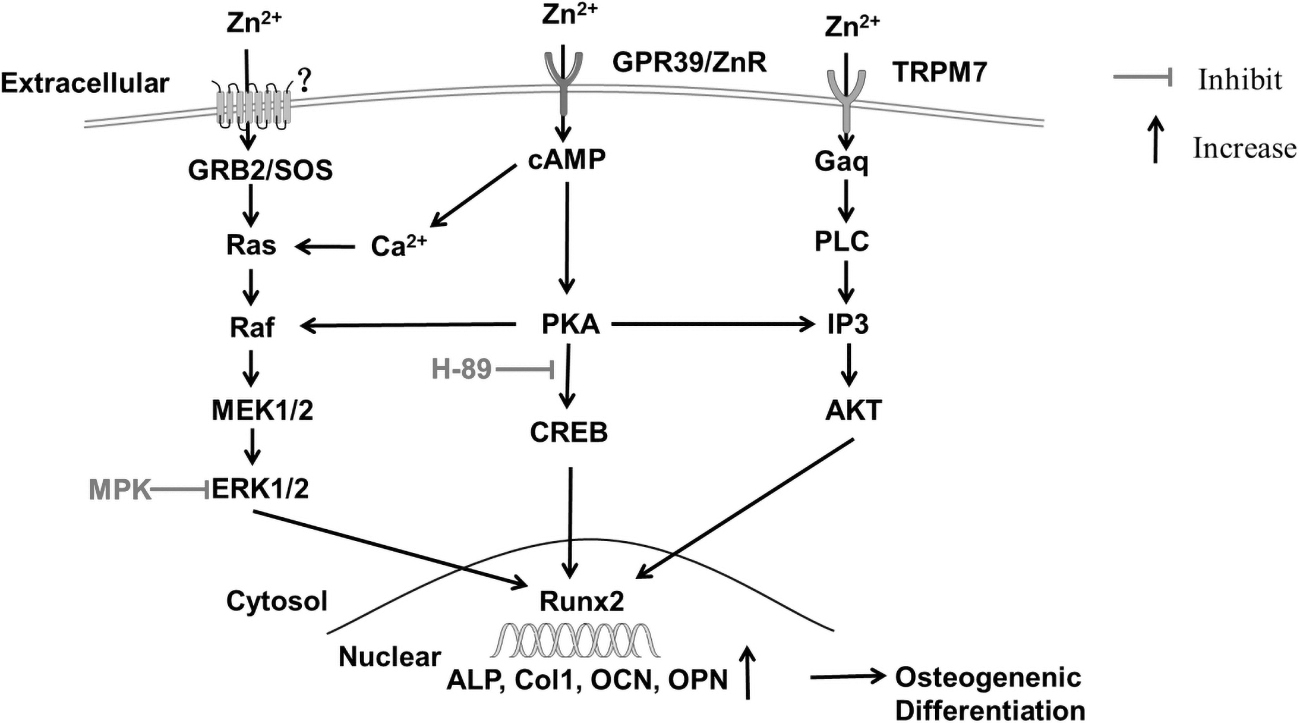
MAPK/ERK1/2 and cAMP/PKA signaling pathway of zinc effect on osteoblast differentiation of BMSCs. The extracellular Zn2+ binds to the receptor on the membrane, and activates GRB2 to bind to SOS, further activating Ras by consuming GTP, thereby activating the downstream Raf, MEK1/2, and ERK1/2. The activated ERK1/2 transfers to the nucleus and induces osteogenesis-related marker genes such as Runx2, ALP, Col-I, OCN, and OPN, and these genes are expressed to produce osteogenic differentiation. In addition, MPK, an inhibitor of ERK1/2, can decrease the expression of Runx2 in the nucleus and inhibit the osteogenic differentiation of BMSCs. Zn2+ can activate the cAMP/PKA pathway through the membrane receptor GPR39 or TRPM7, and cause the increasing expression of osteogenic genes in the nucleus. Zn2+ can also activate AKT and affect the osteogenic differentiation of BMSCs. In addition, the extracellular Zn2+ cloud is transferred to the cell and activates the cAMP-PKA pathway, triggering the Ca2+ response in the cytoplasm, thereby activating MAPK. ALP, alkaline phosphatase; Col-I, type 1 collagen; ERK, extracellular signal-regulated protein kinases; MAPK, mitogen-activated protein kinase; OCN, osteocalcin; OPN, osteopontin.
Moreover, ZIP1, a ZnT in BMSCs, appears to promote osteoblastogenesis by increasing the expression of specific osteoblast-associated genes (Tang et al., 2006). It is found that the expression of bone formation marker genes, including Runx2, ALP, OCN, and OPN, decreases by using the MEK-specific inhibitor U0126 blocking ERK activation, suggesting that the ERK pathway plays a vital role in osteoblast differentiation (Gao et al., 2021). This result is consistent with using special ERK1/2 inhibitor PD98059 (Fathi et al., 2017).
Besides the MAPK pathway, zinc can involve in the cAMP-PKA pathway to promote osteogenesis of BMSCs. For instance, Zhu et al. (2017) confirmed that extracellular Zn2+ is transferred into cells by cellular receptors and then activates the cAMP-PKA pathway and triggers Ca2+ responses in the cytoplasm, leading to the activation of the MAPK pathway. Further study found that the major cellular receptors might be TRPM7 and GPR39, which can facilitate Zn2+ entry into BMSCs (Fig. 2). BMP9 can induce osteogenic differentiation of MSCs by activating the PKA/CREB pathway (Zhang et al., 2015).
The cAMP-PKA pathway containing cAMP, PKA, and CREB upregulates the expression of osteogenic genes and eventually promotes the differentiation and mineralization of BMSCs (Chuang et al., 2020). Using PKA inhibition shPKA, it is found that ALP activity and mineralization are significantly decreased in zinc-induced osteogenic differentiation of BMSCs (Park et al., 2018). Remarkably, it has been reported that the PKA inhibition by using H-89 blocks the nuclear phospho-CREB induced by ZnSO4, and, interestingly, suppresses phosphorylation of AKT and β-catenin induced by ZnSO4 (Park et al., 2018).
After entry into cytoplasm, zinc also triggers the Gαq signaling pathways; then the activated Gαq activates the PLC and AKT signaling pathways, which also reflect osteogenic differentiation (Wang et al., 2020; Zhu et al., 2017). The Gαq inhibition YM254890 and the PLCβ inhibitor u73122 largely attenuate the Zn-dependent Ca2+ response (Zhu et al., 2017). Future studies should further elucidate the detailed cross talk between PKA/CREB signaling and other signaling molecules in osteogenic differentiation of BMSCs and bone formation.
Wnt/β-catenin signaling pathway also exerts a considerably essential role in osteogenic differentiation of BMSCs induced by zinc (Gao et al., 2021; Gaur et al., 2005; Park et al., 2018). Wnt/β-catenin pathway-specific genes comprise WNT5a, FZD4, LRP5, and AXIN2 during the process of osteoblast differentiation of BMSCs. In this pathway, the Wnt ligand binds to the transmembrane frizzled (Fz) receptor and forms a complex with the low-density lipoprotein receptor-related protein (LRP) co-receptor, originating a Wnt/Fz/LRP complex, thereby inhibiting the β-catenin cytoplasmic degradation (Wang et al., 2014).
Then β-catenin modulates the transcription of Runx2 by accumulation and transport in the nucleus, which promotes the osteogenic differentiation and mineralization (Fig. 3) (Fernandes et al., 2020; Shares et al., 2018; Wang et al., 2014). A study reported that zinc deficiency leads to a decreased osteogenic differentiation due to reduced Runx2 expression through the inhibition of the Wnt/β-catenin signaling by the suppression of glycogen synthase kinase 3β (GSK3β) inhibition and Akt activation preceded by the reduced level of SP-1 protein (Hie et al., 2011).
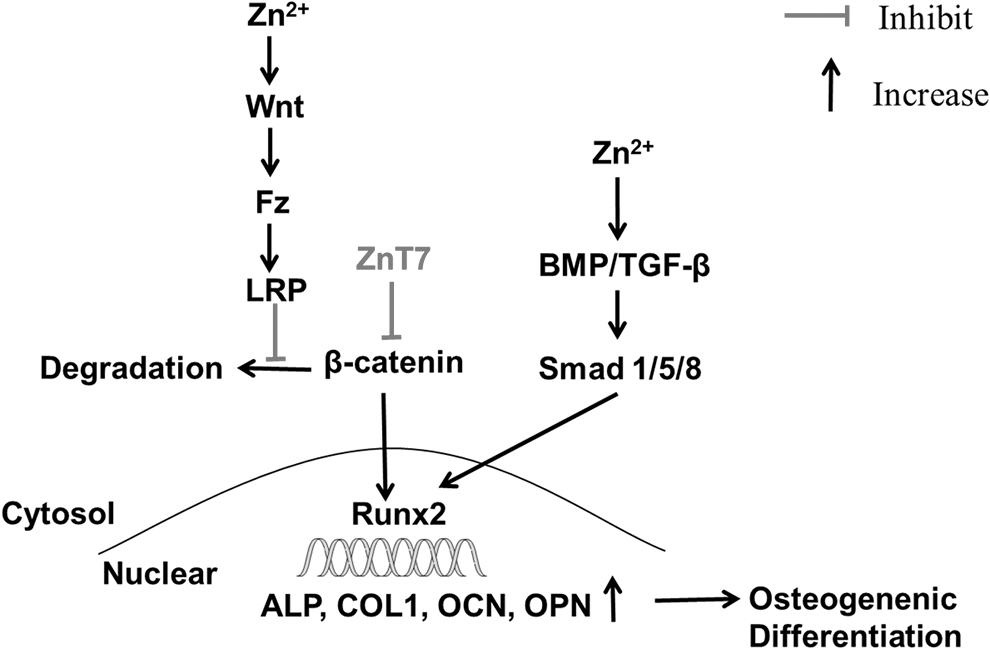
Wnt/β-catenin and BMP/Smad signaling pathway of zinc effect on osteoblast of BMSCs. Intracellular zinc ions can activate Wnt. Wnt ligands bind to transmembrane coiled (Fz) receptors and form complexes with LRP co-receptors to produce Wnt/Fz/LRP complex, which inhibits the cytoplasmic degradation of β-catenin according to specific signal transduction. Then β-catenin accumulates and transfers to the nucleus to regulate the transcription of osteoblast-specific genes. ZnT7 may also participate in the Wnt/β-catenin signaling pathway to inhibit the osteogenic differentiation of BMSCs. Moreover, Zn2+ participates in the BMP/Smad signaling pathway to enhance the expression of osteogenic differentiation genes, thereby promoting osteogenesis of BMSCs. Fz, frizzled; LRP, low-density lipoprotein receptor-related protein; ZnT, zinc transporter.
However, neither Akt inhibitor nor Wnt inhibitor (DKK1) completely reverses zinc-induced osteogenic effects (Park et al., 2018). Thus, the Wnt/β-catenin pathway may not mainly be a cellular signaling pathway on osteogenic differentiation of BMSCs with the present zinc, which is in line with a study found by Fernandes et al. (2020) that the bone regeneration might be regulated, at least in part, by the augmented signaling through the canonical Wnt pathway.
The TGF-β/BMP signaling pathway can regulate differentiation from early BMSCs to mature matrix-secreting osteoblasts induced by zinc (Gao et al., 2021; Grafe et al., 2018; Yusa et al., 2016). For instance, Gao et al. (2021) found by Kyoto Encyclopedia of Genes and Genomes that the TGF-β signaling pathway is overexpressed during osteogenic differentiation of BMSCs (Fig. 3). In vitro study by Yusa et al. (2016) had observed that eluted zinc ions (EZ) stimulate TGF-β signaling pathway-related mRNA expression.
Furthermore, using BMP antagonist noggin, these genes are downregulated in EZ-treated culture. TGF-β family members include BMPs, TGF-βs, activins, and inhibins (Grafe et al., 2018). TGF-β activates their receptors and induces the phosphorylation of intracellular transcription factor Smads; then the activated Smad complex can translocate to the nucleus to trigger the transcription of osteoblast marker genes Runx2, ALP, and Col-I, and eventually promotes osteogenic differentiation and mineralization of BMSCs (Heldin et al., 1997; Yu et al., 2017a). Yu et al. confirmed that Zn-modified calcium silicate coatings significantly upregulate the expression of TGF-β1 and Smad2/3, and zinc induced expression Runx2, Smad2/3, and p-Smad2/3 protein levels are reduced by using the TGF-β inhibitor SB431542, suggesting that zinc induced Runx2 expression through a TGF-β/Smad2/3 mediated process.
Moreover, TGF-β/BMP signaling also stimulates osteoblastic proliferation and differentiation of BMSCs through the Smad1/5/8 pathway (Chen et al., 2012; Yang and Moses, 1990). A study reported that zinc importer protein ZIP13 is involved in the BMP/TGF-β signaling pathway during connective tissue development, a process that includes osteogenesis and cartilage development (Fukada et al., 2008). ZIPs can import extracellular zinc ion into the cytoplasm, which have great prospects in BMSC maturation. However, ZIPs' effects remain to be elucidated on BMSCs' mature process, which includes osteoblast and chondrocyte differentiation.
Zinc Effects on the Chondrocyte Differentiation of BMSCs
As the earliest phase of skeletal development, chondrocytes are differentiated from bone marrow MSCs and exert a crucial role in endochondral ossification (Goldring et al., 2006; Nishimura et al., 2014). Zinc ion can modulate the expression of chondrogenesis-related genes, including Sox-9, aggrecan, chondroadherin, Col-II, and Col-X (Hozain et al., 2021). The transcription factor, Sox-9, is a key regulator of chondrocyte differentiation of BMSCs (Hozain et al., 2021). Sox-9, an early chondrogenesis-related gene, secures chondrocyte lineage commitment and augments cell survival, importantly activating cartilage-specific markers to promote chondrocyte differentiation (Lefebvre and Dvir-Ginzberg, 2017). Col-II and Col-X are the late markers of hyaline cartilage, thus marking these proteins as a functional indicator of mature chondrogenesis (Khader and Arinzeh, 2020; Zheng et al., 2009).
Moreover, chondroadherin, as a mature marker of cartilage, plays a role in the interaction of collagen fibril organization and cell matrix (Table 1) (Zhang et al., 2010). The effects of zinc on chondrocyte differentiation of BMSCs are in a dose-dependent manner (Khader and Arinzeh, 2020; Ovesen et al., 2004). Khader and Arinzeh (2020) found that the chondrogenic differentiation is enhanced where BMSCs on the low percentage of ZnO, which manifests increased Sox-9 and Col-II production and expression of cartilage-specific genes. Nevertheless, osteogenic differentiation is promoted where BMSCs on a high percentage of ZnO, demonstrating the highest ALP, Col-I, and expression of bone-specific genes.
The mechanisms by which zinc affects chondrocytes and cartilage formation are not well defined. Hozain et al. (2021) found that ZnCl2 treatment on ATDC5 cells could be inducing chondrocyte differentiation and hypertrophy through VEGF-mediated signaling. Insulin-like growth factor 1 (IGF-1) has been proved to enhance the proliferation and expression of chondrocyte-related genes during chondrocyte differentiation of MSCs independently through TGF-β signaling (Longobardi et al., 2006). Although zinc has insulin-mimetic properties to promote chondrogenesis (Khader and Arinzeh, 2020), its molecular mechanisms require further clarification.
Reciprocal Effects of Zinc and BMSCs on Osteoclast Differentiation
The complicated interaction between zinc and osteoclast
As an essential trace element, zinc is necessary for osteoclastogenesis. Bone remodeling is mainly composed of osteoblast-induced bone formation and osteoclast-induced bone resorption (Yusa et al., 2016). Zinc exerts a crucial role in bone metabolism by regulating the behavior of (pre)osteoblasts and osteoclasts. The effects of Zn on both the activity and the formation of osteoclasts are complicated. Several previous reports generally suggested that zinc inhibits osteoclastic differentiation (Roy et al., 2013; Yamada et al., 2008). For instance, Yamada et al. (2008) noticed that the number of apoptotic osteoclasts is significantly 6.3 times higher with Zn-incorporated (0.633 wt.%) tricalcium phosphate ceramic (TCP) than with the pristine TCP after 1-day culture. Xiong et al. (2017) showed that reduced graphene oxide/ZS/calcium silicate extracts suppress the RANKL-induced osteoclastic differentiation of preosteoclast RAW264.7 cells.
However, these results are not in line with the finding of Luo et al. (2014), who observed that the proliferation and tartrate-resistant acid phosphatase (TRAP) activity of RAW264.7 cultured on 45 mmol Zn-TCP are significantly higher than those cultured on zinc-free TCP, suggesting that zinc has a positive influence on osteoclastic differentiation. Although the existence of a complex connection between zinc and osteoclastogenesis has been reported previously, the molecular mechanism accounting for this connection remains to be elucidated at length.
The effect of zinc on osteoclast differentiation may be stimulative or suppressive, depending on the dosage of zinc in the environment. For instance, Luo et al. (2014) manifested that zinc in low concentration (i.e., 0.18 and 1.8 ppm) stimulates osteoclastogenic differentiation. The result is partly in line with a study by Yamaguchi and Weitzmann (2011), who demonstrated that zinc dose in the range of 10–250 μM (i.e., 0.65 and 16.25 ppm) suppresses osteoclast formation. However, only very low doses of zinc (>1 μM) are found to be an inhibition of osteoclastogenesis (Hie and Tsukamoto, 2011). Thus, further studies are required to explain osteoclastogenesis in a zinc concentration-dependent manner.
Osteoclasts are differentiated from monocyte-macrophage cell lines rather than bone marrow MSCs. On EZ-induced dental pulp stem cell culture, it is found that the expression of osteoclast-related gene RANKL and osteoprotegerin (OPG) mRNA have no significant changes (Yusa et al., 2016). Therefore, zinc may not differentiate BMSCs into osteoclasts. However, of particular note, zinc promotes the maturation of osteoclasts through the stimulation of osteoblast proliferation (Gurban and Mederle, 2011; Kang et al., 2018; Meng et al., 2019).
For instance, Kang et al. (2018) confirmed that Zn-Al LDH increases the expression of OPG and RANKL mRNA, suggesting that zinc-induced osteoblasts are able to modulate osteoclastogenesis by investigating both RANKL and OPG. Osteoclastogenesis-related genes include RANKL, OPG, RANK, TRAP, NFATc1, Car2, and MMP-9, which are closely associated with osteoclastic activity (Table 1) (Hie et al., 2011; Xiong et al., 2017). Osteoblasts can produce two osteoclastogenesis-related molecules: RANKL and OPG (Yusa et al., 2016).
RANKL binds to its receptor RANK, and induces the development and activation of osteoclasts, while OPG functions as a decoy receptor and blocks RANKL-RANK binding and osteoclast differentiation (Boyce and Xing, 2008; Stern et al., 2007; Yusa et al., 2016). TRAP activity and giant cell fusion are markers of osteoclastogenic differentiation of mononuclear precursors (Luo et al., 2014). Interestingly, Meng et al. (2019) found by establishing co-culture of BMSCs induced by zinc and preosteoclast RAW 264.7 cell that the expressions of TRAP5b and interleukin (IL)-1 are significantly enhanced at 8- and 15-day co-culture, and nucleus integration and formation of the multinucleated cells are moderately stimulated at 14-day co-culture, suggesting interacted effects of zinc and BMSCs on the osteoclastic activity. Nevertheless, the underlying mechanism of the highly reciprocal role of Zn and BMSCs requires to be further explored in detail.
Effects on zinc osteoclast differentiation molecular mechanisms
On preosteoclast monoculture induced by zinc, the NFATc1 signaling pathway is crucial for osteoclastic differentiation (Park et al., 2013; Zhao et al., 2010). Park et al. (2013) confirmed that the inhibitory effect of zinc on osteoclast differentiation attributed to the suppression of the NFATc1 signaling pathway. Yamaguchi and Weitzmann (2011) found that zinc blocks osteoclastic differentiation by inhibiting the NF-κB signaling pathway. Several previous reports found that the binding of RANKL to RANK activates the NF-κB signaling pathway that is capable of increasing the expression of the NFATc1 gene (Asagiri et al., 2005; Zhao et al., 2010).
Similarly, zinc suppresses the expression of RANKL and RANK during osteoclastogenesis, which inhibits NFATc1 expression level by blocking the activation of NF-κB, and eventually leads to osteoclast activity attenuation (Hie and Tsukamoto, 2011; Park et al., 2013; Yamaguchi and Uchiyama, 2004). NFATc1, as a master regulator in regulating osteoclastogenic differentiation, is capable of controlling the osteoclast-related gene expression, including TRAP, MMP-9, and Car2 (Kim et al., 2014). It is worthy to note the decreased osteoclastogenesis by suppressing RANK expression through a reduction of MITF protein caused by Zn deficiency (Hie et al., 2011).
When osteoblasts and osteoclasts co-culture in the presence of zinc, the RANK/RANKL/OPG pathway plays a crucial role in bone remodeling regulation (Gurban and Mederle, 2011). Osteoblast metabolism is found to affect osteoclast differentiation largely due to the RANKL and OPG produced by osteoblast (Kang et al., 2018).
More in vivo and ex vivo studies have shown that the interaction of osteoblast and Zn modulates the osteoclastic differentiation by regulating the RANK/RANKL/OPG pathway (Fig. 4) (Gurban and Mederle, 2011; Kobayashi et al., 2009; Meng et al., 2019). Meng et al. (2019) confirmed that TRAP5b and IL-1 were significantly increased at 8- and 15-day co-culture, although osteoclastic activity was reduced at 1-day co-culture. Furthermore, IL-1 might play a crucial role in the effect of zinc on osteoclast function in the presence of osteoblasts (Meng et al., 2019). IL-1 regulates a downstream factor TRAF-6 by NF-κB and JNK pathways, and TRAF-6 can modulate osteoclast formation (Wright et al., 2009).

The effect of osteoblast and adipocyte differentiation of BMSCs induced by zinc on osteoclasts. Zinc can directly inhibit osteoclasts of BMSCs by suppressing the expression of RNAKL and RNAK, by blocking the activation of NF-κB to inhibit NFATc1 expression level. Moreover, zinc is capable to indirectly modulate osteoclast differentiation through spurring formations of osteoblast and adipocyte products. NF-κB, nuclear factor kappaB.
OPG produced by osteoblasts acts by binding to neutralizing both the soluble RANKL, which prevents osteoclast formation, and osteoclastic bone resorption by inhibiting the RANKL-RANKL receptor interaction (Udagawa et al., 2021). Moreover, various cytokines synthesized and released by adipocytes, such as tumor necrosis factor-α and IL-6, can also stimulate bone resorption (Fig. 4) (Ailhaud et al., 1992; Mundy et al., 1996). Thus, how osteoblasts inhibit differentiation of osteoclasts requires further exploration in detail.
Zinc Effects on the Adipocyte Differentiation of BMSCs
Several pathological conditions, including advanced age, osteoporosis, or osteonecrosis, have shown to enhance differentiation of BMSCs into adipocytes, which results in the increase of adipocytes and the decrease of bone mass (Valenti et al., 2016). As pluripotent progenitors, BMSCs are capable of differentiating into osteoblasts, adipocytes, and chondrocytes (Caplan, 1991).
Since both osteoblasts and adipocytes are derived from the common cellular precursors, named BMSCs, a reciprocal affiliation exists between adipogenesis and osteogenesis of BMSCs, which often is observed in human and animal models (Georgiou et al., 2012; Morita et al., 2006), consistent with the stimulation of proliferation and differentiation of osteoblasts and the suppression of adipocytic differentiation of BMSCs in the presence of zinc (Li et al., 2015). Similarly, bone marrow adiposity is opposite correlated with bone mass in postmenopausal osteoporosis (Patsch et al., 2013).
Thus, a hypothesis was suggested by Li et al. (2015) that BMSCs diverse toward osteoblast formation is a “switch-like” to promote osteoblastic differentiation and suppress adipocytic formation. A growing body of evidence supports that the severity of osteoporosis is indicated by observing the degree of adipocyte differentiation of BMSCs (Li et al., 2014). Therefore, the inverse relationship between adipogenesis and osteoblastogenesis may be valuable for better understanding the pathogenesis of postmenopausal osteoporosis and the prevention of fracture.
The effects of zinc ion on adipogenesis showed a biphasic dose–response and incubation time dependence. Wang et al. (2007), by in vitro, found that zinc ion concentration (at 10−11 and 10−10 M) suppressed adipocyte differentiation of BMSCs, and the effect was reversed to promote or even be removed along with the increasing zinc ion (at 10−9 M). Moreover, the suppression effects were attenuated with the increase in incubation time. A study on the molecular mechanism of effects of zinc on adipocyte differentiation of BMSCs is later to be reported.
Liu et al. (2013) established an obese mice model and found that zinc deficiency increases the production of leptin and the infiltration of macrophages into adipose tissue. The prospective trials suggested by Li et al. (2015) require clarifying the time sequence of gene expression as the differentiation of BMSCs to adipocytes in vitro, to explore potential targets for interventional zinc therapy.
Conclusion
Zinc ion can promote bone marrow MSCs to differentiate into various cell phenotypes, including osteoblasts, chondrocytes, and adipocytes, and the increase of osteoclast formation along with BMSC co-culture. The four main directions of studies support zinc as a therapeutic agent to promote bone regeneration. Several signaling pathways such as MAPK pathway, Wnt pathway, BMP/TGF-β pathway, and cAMP pathway are involved in the process of osteoblastogenesis of BMSCs. Moreover, the proliferation and differentiation of osteoblast-lineage cells are controlled by the reciprocal regulation of four signaling pathways, but not by their independence. During the process of chondrocyte differentiation of BMSCs induced by zinc, Sox-9 is the most important gene, which is an indicator for chondrocyte formation, and VEGF and IGF-1 are related to this process.
The effect of zinc on osteoclast differentiation is complicated, influenced by the dosage of zinc in the environment. Zinc interacting with osteoblasts can promote osteoclastic differentiation by regulating the RANK/RANKL/OPG pathway. In addition, zinc can physiologically suppress adipocyte formation of BMSCs, which is associated with leptin and the infiltration of macrophages into adipose tissue.
Future Directions
Zinc, as an essential trace element, exerts crucial roles in bone metabolism. The formation of bone mass and bone resorption maintains skeleton metabolism. Osteoporosis attributes to an out-of-balance between osteoblast and osteoclast activity, which eventually gives rise to bone loss (Tanaka et al., 2005). Zinc is a component of bone and a fundamental cofactor of many cellular proteins involved in cellular and bone homeostasis (Huang et al., 2020). The cellular and bone homeostasis are modulated in the presence of supplemental zinc ion by stimulating osteoblastic bone formation and preventing osteoclastic bone resorption by regulating protein synthesis and gene expression. Zinc and genistein can restrain osteoclastogenesis induced by various bone-resorbing factors in bone marrow culture by OPG-RANKL-RANK system and Ca2+ signaling.
In addition, zinc stimulates bone formation by increasing production of IGF-I and TGF-β1, and the combination of zinc and genistein increases protein synthesis by directly activating aminoacyl-tRNA synthetase, which is a rate-limiting enzyme at translational process in osteoblastic cells (Yamaguchi, 2012). Zinc ion serving as a first or second messenger triggers signaling pathway, and eventually regulates cellular physiological functions, including proliferation, differentiation, and ion transport (Levaot and Hershfinkel, 2018).
The zinc-containing biomaterials are late model of foreign transplant, possessing the promoting behavior of BMSCs, antibacterial, anti-inflammatory, and angiogenic properties. They are capable of significantly enhancing the survival rate and vitality of BMSCs.
The adhesion and proliferation of BMSCs are affected by the internal properties of biomaterials, such as the surface wetting ability (de Luca et al., 2014), Zeta-potential (Qiu et al., 2018), morphology, and surface chemistry (Burg et al., 2000); grain size and crystallinity of ceramic (Wei et al., 2008); and sustained and slow zinc ion release, and the increase in expression of integrin α1/β1 gene upregulated by zinc is in favor of BMSC behavior (Qiu et al., 2018; Yu et al., 2017b). Thus, the synergistic effect between zinc ion and biomaterials promotes the adhesion and proliferation of BMSCs. Further research on the reciprocal of zinc ion and biomaterials is considered crucial for promoting the survival ability of BMSCs.
An approach is applied to the prevention and treatment of osteoporosis through suppressing the activity of osteoclasts and augmenting the osteoblast differentiation (Park et al., 2018). Furthermore, zinc ions are the promoter of bone regeneration to correct bone disorder by the RANKL/RANK/OPG pathway in postmenopausal osteoporosis (Gurban and Mederle, 2011). Future research directions will probably be related to clarifying the cellular and molecular mechanisms that zinc affects osteoblastogenesis and osteoclast formation to promote bone regeneration. Supplement of zinc alone is proved to be capable to promote antidiabetic action because it possesses insulin mimetics (O'Connor et al., 2020; Vardatsikos et al., 2013).
Insulin and IGF-1 share the common receptor family and cellular transduction pathways (Taniguchi et al., 2006). IGF-1 is demonstrated to induce differentiation of chondrocytes through the PI3k/Akt, PKC/p38, or ERK/MAPK pathway (Oh and Chun, 2003). Therefore, whether these pathways also exist in zinc promoting chondrocyte differentiation of BMSCs requires to be elucidated.
The therapeutic approaches for applying zinc ion agent to delay the occurrence of osteoarthritis, which involves the degeneration of articular cartilage, will be a future study direction. A previous report has shown that zinc inhibits osteoclastogenesis by reducing the secretion of cytokines that are produced by adipocytes (Liu et al., 2013). Several kinds of literature support the stimulation effect on preadipocyte to osteoclast differentiation (Benayahu et al., 1994; Sakaguchi et al., 2000). Thus, the reciprocal interaction of osteoblasts, osteoclasts, and adipocytes may be valuable for better understanding the mechanism of effect of zinc ion on bone. Since zinc ion can increase bone mass by augmenting osteoblastogenesis, zinc-containing grafts can apply to the treatment of fractures to promote bone healing. Further trials in large animal models are required to bring zinc therapeutics to clinical use.
The differentiation potential of stem cells derived from adipose differs from BMSCs upon zinc supplementation. Besides osteogenic differentiation (Haimi et al., 2009), zinc possesses other functions in adipose-derived MSCs. Moon et al. (2018) found that zinc promotes differentiation toward a neuronal fate in adipose-derived MSCs. What is noteworthy is that ZnSO4 reduces adipose-derived MSC aging under the exposure of extremely low-frequency electromagnetic field by increasing telomerase reverse transcriptase TERT gene expression (Fathi et al., 2017). The role of zinc on the differentiation of adipose-derived MSCs will be a novelty.
Footnotes
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This work was supported by grants from the Health Commission of Hunan Province (20200476, 20201907, 202102080160), Fund Project of University of South China for Prevention and Control of COVID-19 (2020SK3010), and Teaching reform research project of University of South China (2020YB-XJG106).