
Abstract
Different development stages of porcine embryos have different tolerance to low temperature. Therefore, we took the porcine embryos after parthenogenetic activation (PA) as the model, to explore the optimal development stage for vitrification during morula (D4), early blastocyst (D5), and expanded blastocyst (D6) after PA (D0). Embryos were observed with microscope and analyzed by different staining after cryo-recovery for 24 hours. The quality of embryos was damaged after vitrification, including embryonic nuclei, DNA, cytoskeleton, and organelles. The re-expansion rate at 24 hours of D5 embryos was significantly higher than those of D4 and D6 embryos (D5 vs. D4 vs. D6, 27.620 ± 0.041 vs. 7.809 ± 0.027 vs. 13.970 ± 0.032, p < 0.05). Therefore, D5 embryos were selected as research objects to explore the effect of vitrification on lipid in vitrified embryos. The results showed that the expression levels of perilipin PLIN3 messenger RNA (mRNA) and triacylglycerol synthesis-related genes AGPAT1 and DGAT mRNA are significantly reduced (p < 0.05). Vitrification affected lipid synthesis, which might have an irreversible impact on embryonic development. In conclusion, our data demonstrated that the optimal stage of vitrification was D5 for early blastocysts.
Introduction
The parthenogenetic activation (PA) embryo is a special biological phenomenon in which simple chemical or physical stimulation of the oocyte allows it to develop preimplantation, and both morphologically and preimplantation development reflect normal conditions well (Jolliff and Prather, 1997; King and De Sousa, 2006). PA embryos have been used as a substitute to test new embryo technologies, for example, vitrification (Esaki et al., 2004) and new culture systems (Nguyen et al., 2010). PA embryos have been widely used because of their great application and research (Wu et al., 2016).
Vitrification has been performed in various animals for genetic resource preservation and animal breeding (Dobrinsky, 2002), which could be used to preserve embryos derived from in vitro procedures such as genetic engineering and cloning biotechnology and biomedicine (Castillo-Martin et al., 2014a).
Although the survival rate and conception rate after transplantation have been improved in porcine embryos after vitrification, the survival rate still remains at a relative low level in pig. Many factors should be considered, one of the crucial factors is the higher content of lipid droplets (LDs) in porcine oocytes and embryos (McEvoy et al., 2000), which is highly sensitive to the freezing of porcine embryos (King and De Sousa, 2006).
LDs are dynamic cellular organelle that are present in most eukaryotic cells (Sun et al., 2013), which serve as repositories of fatty acids and sterols and are used for the synthesis of membrane and hormone (Martin and Parton, 2006), in which triglycerides (TAG) are mainly stored and represent important source of energy during embryo development (Sturmey et al., 2006). The developmental ability of embryos comprised because of the content of LDs, which withstand conventional cryopreservation.
Therefore, delipidation was performed in different studies with centrifugation and subsequent removal of polarized LDs to improve survival rates after vitrification (Beeb et al., 2002), but the physical treatment on embryos might cause negative effects on embryonic development. Previous studies have reported that LD content varies in the preimplantation embryonic development stage (Kikuchi et al., 2002).
In addition, embryonic characteristics, including the developmental stages, the size and number of blastomeres, thickness of the zona pellucida (ZP), the permeability of plasma membrane to water, osmotic swelling, and so on, will directly affect the success of cryopreservation. Therefore, the embryonic developmental stage should be considered a critical factor when the vitrification would be performed for cryopreservation of porcine embryos (Jin et al., 2013). LDs in early-stage porcine embryos are abundant and large, but with the development of embryos, the size and content of LDs will decline gradually (Kikuchi et al., 2002; Norberg, 1973). The survival cryopreservation of porcine late-stage embryos with low LD content and smaller-sized LDs is better than early-stage embryos (two, four, and eight cells) (Jin et al., 2013; Nagashima et al., 1992).
Although vitrification has been applied in human (Chen et al., 2016; Hiraoka et al., 2004; Vanderzwalmen et al., 2002), mouse (Ling et al., 2009; Ruihuan et al., 2021), and cattle (Do et al., 2019) with rapid freezing rates (Chen and Tian, 2005) and reduced ice crystals (Rall et al., 1987), various s caused by vitrification in embryos still need to be overcome, especially in pig.
After vitrification, it was reported that the oxidation–reduction (redox) status is disturbed by both decreasing glutathione content and increasing intracellular reactive oxygen species (ROS) levels in porcine embryos (Gupta et al., 2010). Hence, this imbalance between ROS production and cellular antioxidant defenses may cause damage to biomolecules such as DNA, proteins, and membrane lipids, leading to mitochondrial dysfunction, and further induce apoptotic responses and reduce embryo viability (Castillo-Martin et al., 2014b; Zhao et al., 2016).
Therefore, this study was designed to investigate the proper embryonic developmental stage for vitrification of porcine embryos and further to explore the possible effect of vitrification on porcine embryos, while porcine embryos produced by PA were used.
Materials and Methods
Antibodies and chemicals
All materials and methods complied with department policies and procedures regarding conflicts of interest and government ethics. All chemicals were purchased from Sigma-Aldrich Co., Inc. (St. Louis, MO), unless otherwise indicated. All manipulations were performed on a heated stage adjusted to 38.5°C, unless otherwise indicated.
Oocytes and in vitro mature
Ovaries were obtained from a local abattoir and were transported to the laboratory in 0.9% NaCl at 35°C. Ovaries were washed twice and kept at 35°C. Follicular fluid (2–6 mm in diameter) was collected using a disposable syringe with 18-gauge needle and allowed to sediment in a 50-mL conical tube. Cumulus oocyte complexes (COCs) were selected according to their morphologic characteristics, showing at least three layers of compact cumulus cells (CCs) and evenly granulated ooplasm.
COCs were washed thrice in HEPES-buffered tissue culture medium 199 (TCM-199) plus 0.8 mM L-glutamine and 2% (v/v) fetal bovine serum (FBS); random parts of COCs were cultured in groups of 50–60 in four-well dishes (Nunc; Thermo Fisher, Carlsbad, CA) in TCM-199 supplemented with 10% (v/v) FBS, 10% (v/v) porcine follicular fluid, 0.8 mM L-GlutaMAX, 0.242% (v/v) gentamicin, 15 IU/mL serum gonadotropin, and 15 IU/mL human chorionic gonadotrophin (hCG) at 38.5°C in a 5% CO2/20% O2 atmosphere at maximum humidity.
Production of PA embryos
After in vitro maturation for 42 hours, COCs were treated with hyaluronidase (1 mg/mL in TCM-199 culture medium) for about 1 minute to obtain denuded oocytes (DOs). DOs were washed twice in an activation solution consisting of 0.3 M mannitol, 0.1 mM MgSO4, 0.05 mM CaCl2, and 0.01% polyvinyl alcohol. They were then electrically activated in a BTX microslide 0.5-mm fusion chamber (model 450; BTX, San Diego, CA) using a single direct current pulse of 0.86 kV/cm for 80 μs, followed by chemical activation with 5 mg/mL cytochalsin B (CB) in bovine serum-free porcine zygote medium (PZM-3 medium) for 4 hours. Oocytes were then cultured in PZM-3 medium in a 4-well plate, both at 38.5°C in a 5% CO2/20% O2 atmosphere at maximum humidity.
Vitrification and warming of embryos
The handling medium (HM) was HEPES-buffered TCM-199 with 20% FBS. All the vitrification media were prepared with HM. Vitrification and warming steps were performed at 38.5°C. Steps and solution preparation of vitrification were performed as previously described (Xu et al., 2020).
The day of PA was recorded as D0; we collected embryos on morula (D4), early blastocyst (D5), and expanded blastocyst (D6) for vitrification, which was performed by using the Cryotop carrier (Kitazato Biopharma, Shizuoka, Japan). In brief, each handing step, 3–5 embryos were transferred into HM for equilibration, and then embryos were transferred into equilibration solution consisting of 10% ethylene glycol (EG) and 10% dimethyl sulfoxide (DMSO) in HM for 30 seconds; subsequently, COCs were transferred into vitrification solution consisting of 20% EG, 20% DMSO, and 0.5 M sucrose in HM for 20 seconds. Embryos were loaded on Cryotop and plunged into liquid nitrogen for frozen.
All warming solutions were performed at 38.5°C. Vitrified embryos were warmed by transferring them to a warming solution 1 (W1) composed of HM with 0.3 M of sucrose for 1 minute. Subsequently, embryos were transferred into warming solution 2 (W2) composed of HM with 0.2 M sucrose for 3 minutes. After being washed in HM for 5 minutes, embryos were transferred into the original PZM-3 to recover for 24 hours, which was called cryo-recovery.
Assessment of embryos after cryo-recovery
Embryos in the control group at each stage continued to develop without any treatment. D9 embryos were embryos that developed in vitro to the 9th day after PA. After cryo-recovery, the re-expanded vitrified embryos were assessed by morphologic observation under a stereomicroscope. The blastomeres of D4 embryos are uniform, complete, and tightly connected; the edges of D4 are clearly visible. The edge of D5 and D6 embryos is smooth and complete, and the blastocyst cavity is re-formed or there is a tendency to re-form the blastocyst cavity. Embryos were selected for staining by MitoTracker Red (Beyotime), BODIPY®493/503 (Thermo Fisher), Phospho-Histone H2A.X, and DyLight™594 Phalloidin (Cell Signal Technology). Those embryos with complete nuclei, cytoskeleton, and organelles were determined as recovered embryos.
Immunofluorescence staining and confocal microscopy
Embryos were collected and fixed in 4% paraformaldehyde for 30 minutes at room temperature (RT). After washing thrice in washing buffer, embryos were permeabilized in 0.5% Triton X-100 for 1 hour at RT. They were washed thrice in washing buffer and were blocked with blocking buffer PB1 (1% BSA in phosphate-buffered saline) for 1 hour at RT to suppress the nonspecific binding of immunoglobulin G (IgG). Embryos were then incubated with 2 μg/mL primary antibodies overnight at 4°C; after being washed thrice, embryos were incubated with 5 μg/mL secondary antibody for 1 hour at RT. Finally, 10 μg/mL Hoechst 33342 was used to stain nuclei for 15 minutes at RT.
Samples were mounted on glass slides and examined with a confocal laser-scanning microscope (LSM 800 META; Zeiss, Oberkochen, Germany). There were no less than 5 embryos in each group, and each group was repeated 3 times.
Gene expression of vitrified embryos
D5 embryos were used as research objects; the aim was to explore the effects of vitrification on perilipin PLIN3 and TAG synthesis-related genes.
First, RNA was extracted from a pool of 30 embryos in each group using the Dynabeads® mRNA DIRECT™ Kit (Ambion, Invitrogen Life Technologies) following the manufacturer's protocol. The total extracted RNA was loaded into the complementary DNA (cDNA) reaction, and the first-strand cDNA was synthesized using the HiScript® RT SuperMix for qPCR kit (Vazyme Biotech Co., Nanjing, China) following the instruction. Quantification of each messenger RNA level was performed with a real-time polymerase chain reaction using AceQ® qPCR with SYBR® Green Master Mix (Vazyme Biotech Co.) in a reaction volume of 20 μL. 18S was used as internal controls. Primers for relevant genes were designed by the Primer premier 6.0 software as shown in Table 1.
Summary of Quantitative Polymerase Chain Reaction Primers of Target and Reference Genes in Porcine Embryo
F, forward; R, reverse.
Statistical analysis
At least three replicates were performed for each experiment. Data were analyzed by GraphPad Prism 7.0 (Graph Pad Software Inc., San Diego, CA) and were presented as mean percentages (mean ± standard error of the mean). Intergroup differences were compared using t-tests. p-Values lower than 0.05 were considered statistically significant.
Results
The optimal vitrified stage is D5 embryos
As shown in Figure 1 and Table 2, the re-expansion rate of D5 embryos was significantly higher compared with D4 and D6 (27.602% of D5 vs. 7.809% D4 and 13.970% of D6 embryos; p < 0.05), and the re-expansion rate of D4 embryos was the lowest.
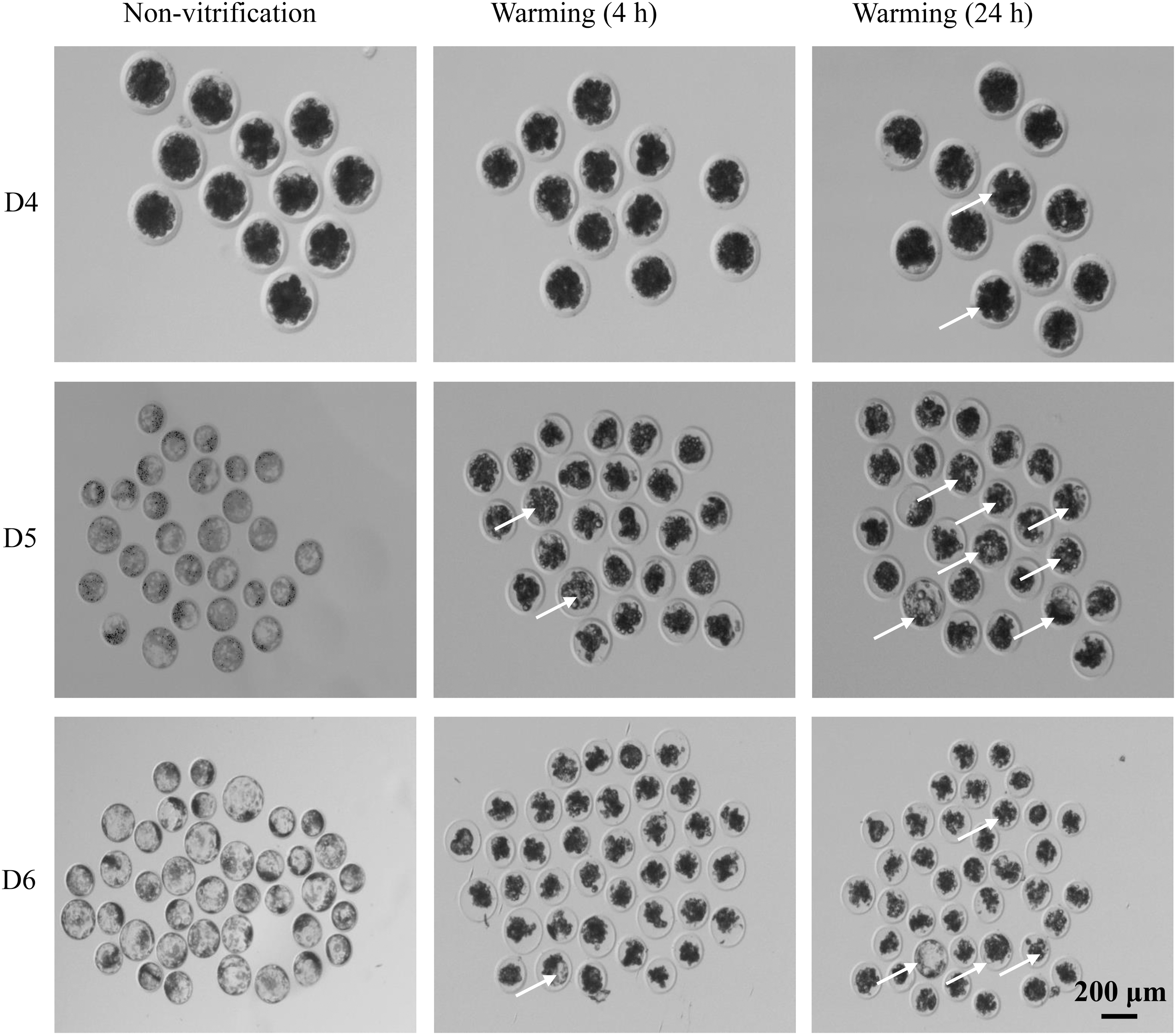
The state of embryos before and after vitrification. The status of embryos before vitrification and cryo-recovery for 4 and 24 hours. D4: morula; D5: early blastocyst; D6: dilated blastocyst. The arrows point to the re-expansion embryos.
Re-Expansion Rate of Porcine Embryos 24 Hours After Vitrification
Different superscripts in the same column indicate significant differences (p < 0.05).
Effect of vitrification on embryonic quality
As shown in Figures 2A and 3A, the nuclei of D4, D5, and D6 embryos were seriously damaged after cryo-recovery. The distribution pattern of the nuclei had changed from uniform distribution to aggregation, and double-strand breaks (DSBs) occurred in embryonic nuclei. After cryo-recovery, the number of nuclei was reduced (p < 0.01, Fig. 2B) and the ratio of embryonic nuclei with DSBs to the total nuclei was significantly increased (p < 0.01, Fig. 3B); the integrity of the cytoskeleton was destroyed, but blastomeres with normal nucleus had complete cytoskeleton (Fig. 4).

The number of embryonic cells in control group and vitrified group. Embryonic nuclei in control group and D4, D5, and D6 embryos after cryo-recovery for 24 hours
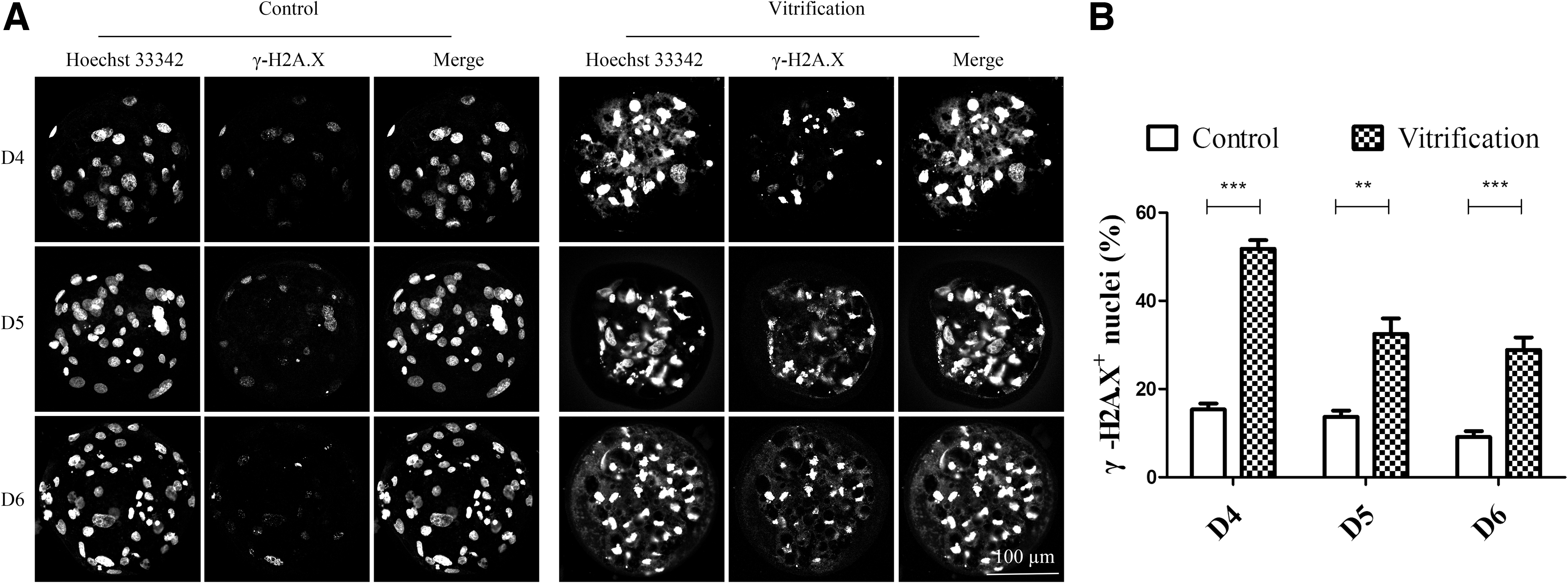
DNA damage in the control group and the vitrified group. Effect of vitrification on induction of DNA damage in D4, D5, and D6 embryos after cryo-recovery for 24 hours
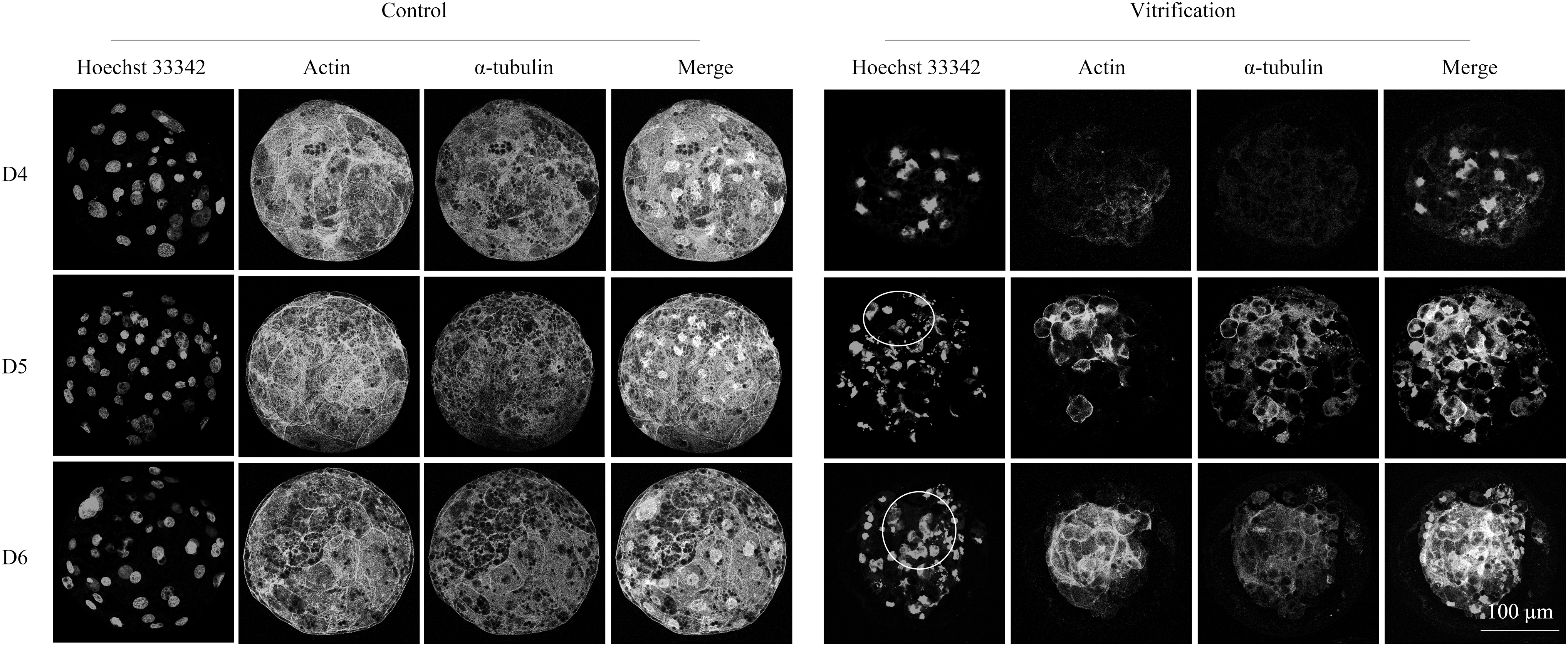
The integrity of the cytoskeleton of the control group and the vitrified group. Microtubules are stained with Actin and spindles are stained with α-tubulin and nuclei. The circles indicate normal cytoskeleton.
Vitrification affected the distribution of mitochondria and LDs in embryos
As shown in Figures 5 and 6A. The cytoplasmic mitochondria and LDs of the embryos of the control group were evenly distributed. After cryo-recovery, the mitochondria and LDs were severely damaged and distributed unevenly in embryos. LDs in vitrified embryos aggregated and became large, which was similar to the LDs morphology of naturally apoptotic D9 embryos (Fig. 6B). However, the normal D9 embryos showed only a decrease in LD content, but no change in morphology.

The distribution of mitochondria in the control group and the vitrified group. Mitochondrial were stained and nuclei were stained with Hoechst 33342.

The distribution of LDs in embryos at different stages. The effects of vitrification on LD distribution in D4, D5, and D6 embryos after cryo-recovery for 24 hours
The effect of vitrification on PLIN3 gene and TAG synthesis-related genes of D5 embryos
As shown in Figures 7 and 8, the gene expression level of PLIN3, AGPAT1, and DGAT in the control group was significantly higher than those in vitrified group (p < 0.05), indicating that vitrification affected lipid synthesis.

Distribution
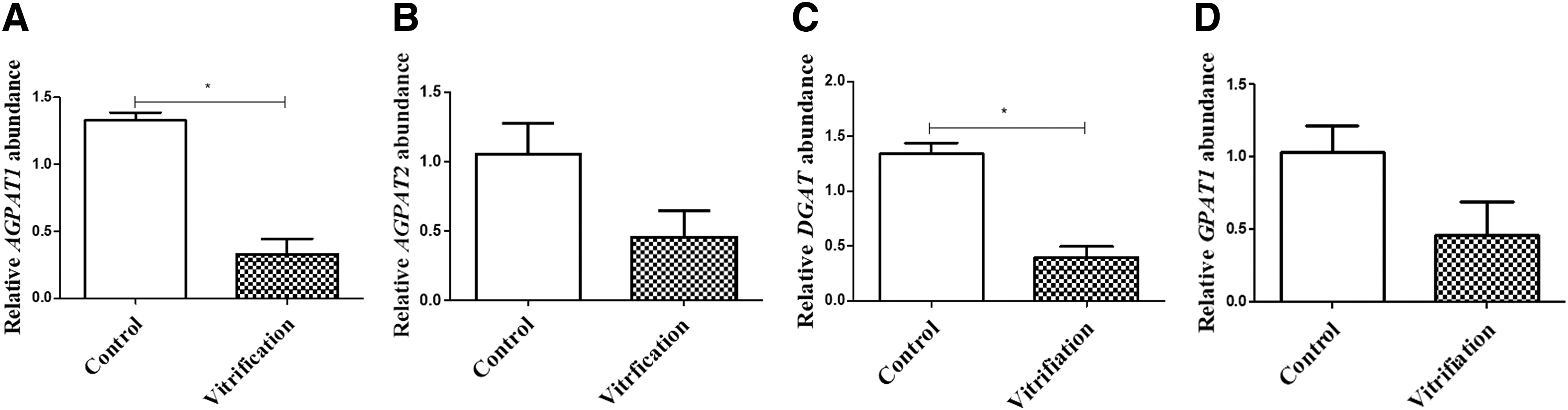
Expression levels of TAG synthesis-related genes in embryos of control group and vitrified group. Embryos in the vitrified group were D5 embryos after cryo-recovery for 24 hours. A–D: The relative expression of genes AGPAT1
Discussion
Although different damages, including nuclei, DNA, cytoskeleton, mitochondria, and LDs, presented in porcine embryos after vitrification-thawing, this study confirmed that D5 (early blastocyst) was the proper developmental stage for porcine embryo cryopreservation by vitrification. The main reason for the damage of porcine embryos might not only be due to the high content of LDs as reported previously (Nagashima et al., 1994) but also due to developmental stage, which affects the cryotolerance of embryos (Morato et al., 2016).
Both embryos produced in vitro (Somfai et al., 2009) and in vivo (Cuello et al., 2008) are very sensitive to low temperature. Embryos in different stages have different numbers of blastomeres, LD content, morphological size, and thickness of ZP, so the vitrification of tolerance is different. Therefore, the choice of embryo stage is very important for vitrification (Jin et al., 2013). Studies have shown that the vitrification effect of morula and later embryos is better than other stages, which may be related to the higher number and the smaller size of cells (Saberi et al., 2018), and improve chilling tolerance when compared to the early-stage embryos with few and larger cells (Szell and Shelton, 1986).
Morula and blastocysts had been widely selected for embryo vitrification with relative acceptable successful rate in pig (Beeb et al., 2002; Fujino et al., 2008; Li et al., 2012; Misumi et al., 2003), human (Hiraoka et al., 2004), and mouse (Zander-Fox et al., 2013). In this study, D4, D5, and D6 embryos are morula, early blastocysts, and expanded blastocysts, respectively. The optimal stage of porcine embryos to vitrify was D5 embryos, as the efficiency decreased when D6 and especially D5 embryos were used. Although previous research had shown that the optimal stage for vitrification is D4 embryos, which includes morula and blastocysts (Li et al., 2012), however, it is not easy to make sure which developmental stage was better.
Also, another study proved that porcine D6 blastocysts showed the higher rates of survival compared with D5 blastocysts after warming (Morato et al., 2016), but the percentages of apoptotic cells in D6 were higher than D5 blastocysts. Study found that the permeability shown by porcine embryos toward glycerol, EG, or water is lower at the early blastocyst stage than at the expanded blastocyst stage (Jin et al., 2013), which can reduce the toxic concentration of cryoprotectants in early embryos. Expanded blastocysts have bigger blastocoelic fluid, in which more ice crystals may form during cooling (Hiraoka et al., 2004). The efficiency of vitrification is negatively correlated with the expansion of the blastocoel (Hiraoka et al., 2004). That might be the reason D5 embryos have a higher survival rate of blastocysts, such as for human embryos (Mukaida et al., 2003).
Vitrification caused damage to the embryos, including ultrastructure, mitochondria (Fabian et al., 2005), and broken cells (Wu et al., 2016). In our results, a part of blastomeres was damaged, which could be one of the reasons why the nuclei were reduced. Another reason is that the frozen embryos need time to recover; some cells underwent mitosis, so the number of nuclei was significantly reduced compared with the control group.
The cytoskeleton is a complex network structure composed of microfilaments, microtubules, and intermediate filaments (Goode et al., 2000). Low temperature induces damage to the cytoskeleton as evidenced by abnormal spindle microtubule configuration, disrupted chromosomes, and disorganized microfilaments (Morato et al., 2008). In this experiment, the cryo-recovery embryos showed that microfilaments and microtubules were destroyed, and this damage mostly appeared in the blastomeres with damaged nuclei, which further impaired embryo development (Gao et al., 2020).
The metabolism and formation of LDs are a dynamic equilibrium process during embryonic development. LDs are mainly composed of neutral lipids, including TAG and cholesterol ester, and provide substrates for energy production, membrane components, and signaling lipids (Welte, 2015). LDs interacting with several other organelles can help the cell adapt to the nutrient environment (Valm et al., 2017), which have been associated with several new biological functions, including embryogenesis, and in response to oxidative stress (Bailey et al., 2015; Welte and Gould, 2017). We found that the LD morphology of damaged embryos by vitrification is similar to that of naturally apoptotic D9 embryos. The changes of LDs are also a factor of embryonic apoptosis.
PAT proteins are known to regulate LD store (Bickel et al., 2009), including PLIN1 (perilipin), PLIN2 (adipose differentiation-related protein), PLIN3 (tail interacting protein of 47 kDa), PLIN4 (S3–12), and PLIN5 (oxidative tissue-enriched PAT protein) (Kimmel et al., 2010). PLIN3 plays an important role in regulating lipolysis and TAG storage in the adipose tissue (Diaz and Pfeffer, 1998), while it can be recruited to LDs, participating in the biosynthesis of new LDs, and its suppression affects LD formation. The lack of PLIN3, probably at an early stage of LD biogenesis, might involve a change in cellular fatty acid uptake, neutral lipid synthesis, or liberation, and further interfere the formation of mature LDs (Bulankina et al., 2009). PLIN3 is target primarily to nascent LDs and remains stable in the cytoplasm when not associated with LDs.
It has emerged as a regulator of LD biogenesis and degradation (Bulankina et al., 2009). It is found that PLIN3 is distributed along the cytoplasmic side of the double-layer endoplasmic reticulum, and the absence of PLIN3 will reduce the size and number of cytoplasmic LDs (Skinner et al., 2009). When RNAi interferes with PLIN3, the storage of TAG is inhibited. After vitrification, LDs became larger and gathered, and further studies had found that the expression levels of PLIN3, AGPAT1, and AGPAT2 were reduced, which affected lipid synthesis. Therefore, in future experiments, it is possible to reduce LDs by reducing the expression of embryonic PLIN3 protein, in combination with cytochalasin (cytoskeleton stabilizers) and laser drilling (a method for causing the blastocyst cavity to collapse and blastocyst fluid to flow out), thereby improving the effect of vitrification.
In conclusion, our findings suggest that vitrification of porcine embryos by Cryotop method is useful; the optimal stage for vitrification was D5.
Footnotes
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This work was supported by the joint found of Nanjing Agricultural University and Xinjiang Agricultural University (KYYJ201907) and Modern Agriculture project of Jiangsu Province (BK2017327).