
Abstract
Reprogramming of human dermal fibroblasts (HDFs) into induced cardiomyocyte-like cells (iCMs) represents a promising strategy for human cardiac regeneration. Different cocktails of cardiac transcription factors can convert HDFs into iCMs, although with low efficiency and immature phenotype. Here, GATA4, MEF2C, TBX5, MESP1, and MYOCD (GMTMeMy for short) were used to reprogram HDFs by retrovirus infection. We found that the exogenous expression stoichiometry of GATA4 (GATA4 stoichiometry) significantly affected reprogramming efficiency. When 1/8 dosage of GATA4 virus (GATA4 dosage) plus MTMeMy was used, the reprogramming efficiency was obviously improved compared with average pooled virus encoding each factor, which measured, by the expression level of cardiac genes, the percentage of cardiac troponin T and alpha-cardiac myosin heavy-chain immunopositive cells and the numbers of iCMs showing calcium oscillation or beating synchronously in co-culture with mouse CMs. In addition, we prepared conditioned maintenance medium (CMM) by CM differentiation of H9 human embryonic stem cell line. We found that compared with traditional maintenance medium (TMM), CMM made iCMs show well-organized sarcomere formation and characteristic calcium oscillation wave earlier. These findings demonstrated that appropriate GATA4 stoichiometry was essential for cardiac reprogramming and some components in CMM were important for maturation of iCMs.
Introduction
Cardiomyocytes (CMs) have little regeneration capacity after birth (Kikuchi and Poss, 2012), so severe chronic heart failure after myocardial infarct can only rely on orthotopic heart transplant, which is limited by compatible donor organs. Cardiac reprogramming of human fibroblasts represents a promising avenue to solve this problem.
In the past decade, ectopic expression of five factors, GMTMeMy was reported to initiate cardiac genes expression in human cardiac fibroblasts (HCFs) and human dermal fibroblasts (HDFs), which made the reprogrammed cells exhibit contraction and action potentials in co-culture with mouse CMs (Wada et al., 2013). Moreover, GATA4, HAND2, TBX5, MYOCD, MiR-1, and MiR-133 were reported to allow the reprogrammed HCFs to exhibit spontaneous contractions by extending the culture period (Nam et al., 2013). Other groups reported success of reprogramming H9 human embryonic stem (hES) cell-derived fibroblasts and HDFs with seven factors, GMTMeMy plus ESRRG and ZFPM2 (GMTMeMyEZ for short) (Fu et al., 2013); then they also found that TGF-β and WNT inhibitors could enhance this seven-factor reprogramming of HCFs (Mohamed et al., 2017).
Around the same time, HAND2 was found to help GMTMeMyEZ to facilitate the early progress of reprogramming of HDFs and H9Fs (Bektik et al., 2017), and MiR-590 could promote reprogramming of HCFs (Singh et al., 2016). However, the reprogramming efficiency and maturity in HDF-derived induced cardiomyocyte-like cells (iCMs) were poorer than those in HCF-derived iCMs (Fu et al., 2013; Nam et al., 2013). HDFs, as an easy obtainable source of autologous cells for reprogramming, seem to be more resistant to the cardiac reprogramming process, so additional manipulations are urgently needed to push HDFs toward the fate of CMs.
A balanced exogenous stoichiometry and a logical introduction sequence of transcription factors (TFs) were considered critical factors of cardiac and pluripotent reprogramming (Liu et al., 2013; Wang et al., 2015). Moreover, some cardiogenic compounds involved in CM differentiation of pluripotent stem cells (PSCs) were used to improved the quality of cardiac reprogramming of mouse fibroblasts (Yamakawa et al., 2015). Therefore, in this study, we adjusted GATA4 dosage to promote cardiac reprogramming of HDFs, prepared conditioned maintenance medium (CMM), and compared the role of traditional maintenance medium (TMM) and CMM in maturation of iCMs. We found that the use of 1/8 GATA4 dosage or later introduction of GATA4 virus markedly improved the reprogramming efficiency; meanwhile, CMM accelerated maturation of iCMs.
Materials and Methods
Differentiating of H9 hES cell line into cardiomyocytes and preparation for CMM
Animal experiments were registered and approved by the Ethics Committee of the First Affiliated Hospital of Harbin Medical University.
H9 hES cells were cultured in mTeSR medium (STEMCELL). When the confluence reached about 70%, the medium was switched to RPMI supplemented with B27 without insulin (Gibco) and 6 mM CHIR-99021 (Selleck). After 48 hours, the medium was changed to RPMI-B27 without insulin for 48 hours, then RPMI-B27 without insulin supplemented with 5 mM IWR-2 (Selleck) for 48 hours, and then changed back to RPMI-B27 without insulin for 48 hours. On day 8, RPMI-B27 (Gibco) supplemented with 2% fetal bovine serum (FBS) (Gibco), 0.05% bovine serum albumin (BSA) (Sigma), and Vitamin C (50 μg/mL; Sigma) was used as TMM for long-term maintenance culture. TMM was collected every 36 hours and then filtered through a 0.45 μm filter (Millipore) to prepare CMM.
Retroviral infection and cell culture
To construct the pMXs retroviral vectors, we amplified the coding regions of GATA4, MEF2C, TBX5, MYOCD, and MESP1 by polymerase chain reaction, subcloned them into pMXs vectors, and then transfected the recombinants and packaging vector into 293T cells with calcium phosphate to produce retroviruses. The supernatant containing viruses was collected 48 and 72 hours after transfection and filtered through a 0.45 μm filter (Millipore). Adult HDF cell line (BJ fibroblasts) was purchased from ATCC.
About 2 × 104 of BJ per well were seeded onto a 24-well plate 24 hours before infection and transduced with freshly prepared virus mixture containing 8 mg/mL polybrene every 24 hours, twice in total. Fluorescence images of BJ induced by five factors and DsRed showed a high infection efficiency of DsRed 48 hours after infection (Supplementary Fig. S1A). The medium was switched to induced medium (10% FBS/20% M199/DMEM; Gibco) supplemented with Vitamin C (50 μg/mL), SB431542 (1 μM; Selleck), and XAV939 (2 μM; Selleck) 24 hours after the last infection and changed every 3 days. TMM or CMM was used 2 weeks after induction and changed every 2 days.
Fluorescence-activated cell sorting analysis
For cardiac troponin T (cTnT) positive assay, cells were fixed with 4% paraformaldehyde (PFA) for 20 minutes, permeabilized with saponin, and stained with primary antibody (anti-troponin-T PE, REAfinity; Miltenyi Biotec, 5201006420) for 10 minutes at room temperature. Cells were run on C6 flow cytometer (Becton Dickinson). Data analyses were performed by the software FlowJo10.
Immunofluorescence
Cells were fixed in 4% PFA for 15 minutes at room temperature, blocked, and incubated with primary antibodies, including rabbit anti-alpha-cardiac myosin heavy chain (α-MHC) (1;100; Proteintech, 22281-1-AP), rabbit anti-myosin light chain 2a (1;50; Proteintech, 17283-1-AP), mouse anti-cTnT (1:100; Abcam, ab8295), and mouse anti-Sarcomeric Alpha Actinin (1;100; Abcam, ab9465), followed by the appropriate secondary antibodies conjugated with either Alexa Fluor 488 or Alexa Fluor 546 (Molecular Probes), and finally with DAPI (Invitrogen).
qRT-polymerase chain reaction
Total RNAs were prepared with TRIzol. For quantitative polymerase chain reaction (qPCR), purified RNA was reverse transcribed into cDNA using ReverTra Ace (Toyobo) and oligo (dT) (Takara), and then analyzed by qPCR with Premix Ex Taq (Takara). Relative mRNA levels were normalized to those of glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Primer sequences for RT-qPCR are provided in Table 1.
Overview Primers Used for Quantitative Polymerase Chain Reaction
Co-culture of iCMs with mouse neonatal CMs
On day 14, iCMs were treated with trypsin, and then replated onto neonatal mouse CMs at a ratio of 1:8 with DMEM/M199/10% FBS medium. Neonatal mouse CMs were isolated from P3.5 mouse pups with Collagenase II (Sigma) and 0.05% Trypsin (Gibco) (2:1). After preplating on uncoated dishes for 2.5 hours to remove adherent non-CMs, the nonadherent CMs were counted and plated with DsRed-labeled iCMs together. Forty-eight hours after co-culture, medium was switched to TMM.
Imaging calcium oscillation
For detecting calcium oscillation,5 μM Fura 2-AM (Dojindo Laboratories) supplemented with 0.05% Pluronic F-127 (Dojindo Laboratories) was used to label BJ and iCMs for 45 minutes; the cells were washed and allowed to de-esterify the Fura-2 AM in HBSS (Cienry) at 37°C for 30 minutes. Ca2+ imaging was performed by the Eclipse Ti-U Ca2+ Imaging System. About 45 mM KCl solution was used to stimulate cells. We detected [Ca2+]i in individual cell clusters by exciting Fura-2 alternately at 340 and 380 nm. Data analyses were performed by the MetaFluor soft.
Transmission electron microscopy
iCMs and H9-CMs were fixed with 2.5% glutaraldehyde for 30 minutes, then washed five times with phosphate-buffered saline (PBS), and then osmium acid was used to fix cells for 20 minutes. Samples were dehydrated by an ethanol gradient and embedded in resin, and then left at 37°C for 20 minutes and at 20°C in dry environment for a night. Next day, the sample was embedded, percolated at 40°C for 1 day, and polymerized at 60°C for 1 day; the samples were then visualized on a transmission electron microscope (Tecnai G2 Spirit) after trimming, sectioning, and staining.
Results
GATA4 stoichiometry markedly affected cardiac gene expression in 5F-induced HDFs
In this study, GMTMeMy (5F)-induced HDFs were cultured with modified reprogramming medium containing TGF-β inhibitor, WNT inhibitors, and Vitamin C, which were previously reported to enhance cardiac reprogramming (Mohamed et al., 2017; Talkhabi et al., 2015). Medium was replaced by maintenance medium 2 weeks after reprogramming (Fig. 1A). First, we analyzed cardiac gene expression profiles of 5F-induced HDFs by qPCR after 1 week of reprogramming. 5F initiated strong expression of multiple cardiac genes, including key cardiac marker and TNNT2, while MYH6, as another important cardiac marker, was quite poorly induced (Fig. 1B). The exogenous expression stoichiometry of each TF showed that the GATA4 stoichiometry was obviously higher than MEF2C or MYOCD stoichiometry (Fig. 1C).
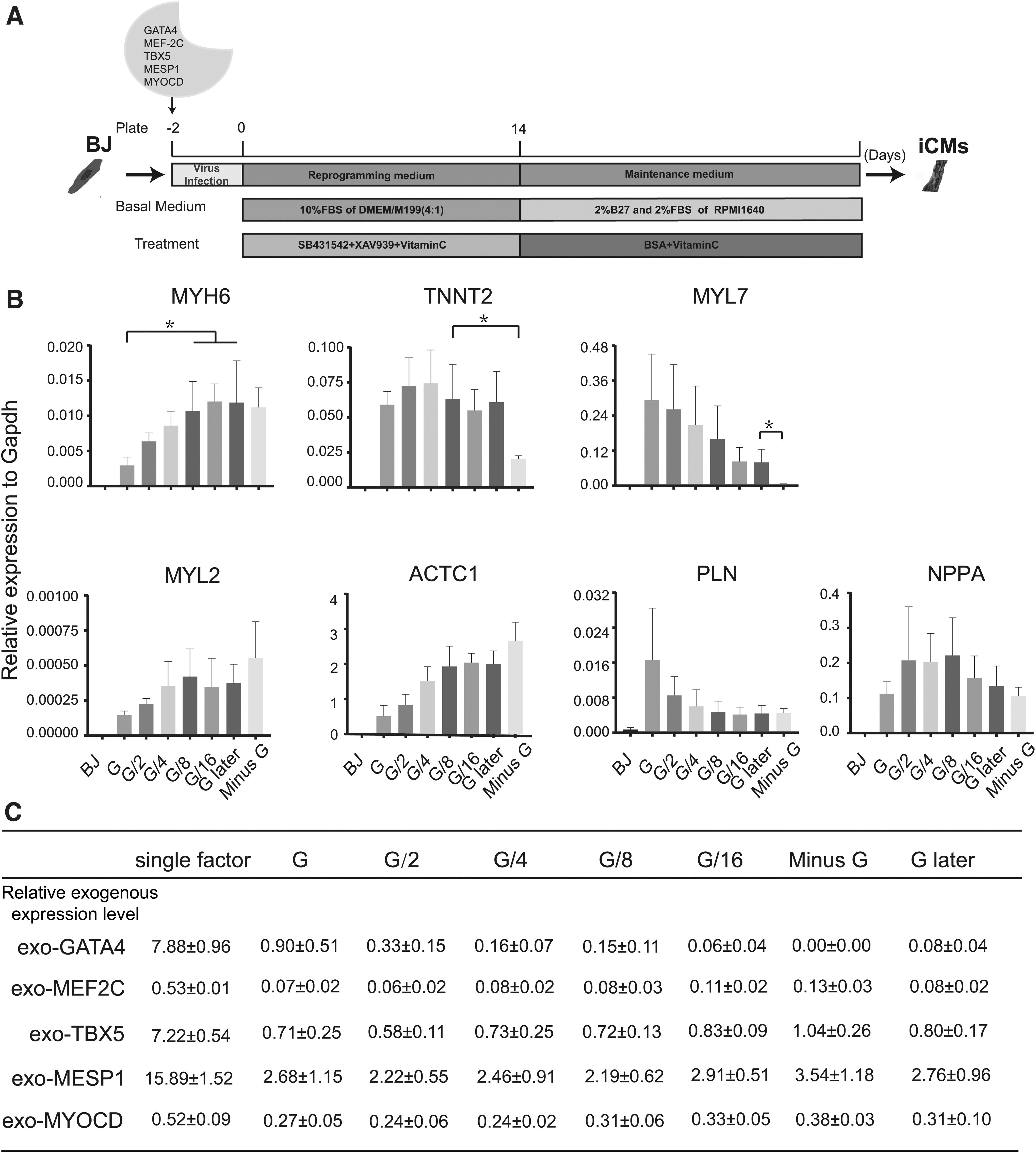
The effects of GATA4 stoichiometry on cardiac gene expression.
However, it was reported that higher MEF2C stoichiometry than GATA4 stoichiometry was associated with high efficiency in GMT-based mouse cardiac reprogramming (Wang et al., 2015). To analyze the effects of GATA4 stoichiometry on cardiac gene expression, MTMeMy plus different GATA4 dosage was used. With reduction of GATA4 dosage or later introduction of GATA4, the GATA4 stoichiometry dropped gradually (Fig. 1C) and the expression of MYH6 was upregulated dramatically in G/8, G/16, and G later groups (Fig. 1B). Meanwhile, we also found that induction of MYH6, MYL2, and ACTC1 was not depended on GATA4, but GATA4 was critical to induce TNNT2 and MYL7 (Fig. 1B).
Different GATA4 dosage resulted in varying α-MHC or cTnT-positive iCMs
During the early stage of reprogramming, iCMs in G/8 and G later groups gradually appeared thick and multinuclear and developed a rod-like shape, while such morphological changes were not obvious in G, G/2,G/4, and Minus G groups (Fig. 2A). The effects of different GATA4 dosage on reprogramming outcomes were assessed by immunostaining and fluorescence-activated cell sorting (FACS). Consistent with mRNA results, 1/16 GATA4 dosage increased reprogramming efficiency within 1 week to ∼4% α-MHC positive iCMs compared to ∼0.4% in G group (Fig. 2B). After 2 weeks of reprogramming, FACS analyses demonstrated that the percentage of cTnT-positive iCMs were most in G/2 and G/4 groups (23.1% and 23.4%), while G and G/8 groups had a similar percentage of cTnT-positive iCMs (14.9% and 15.3%).
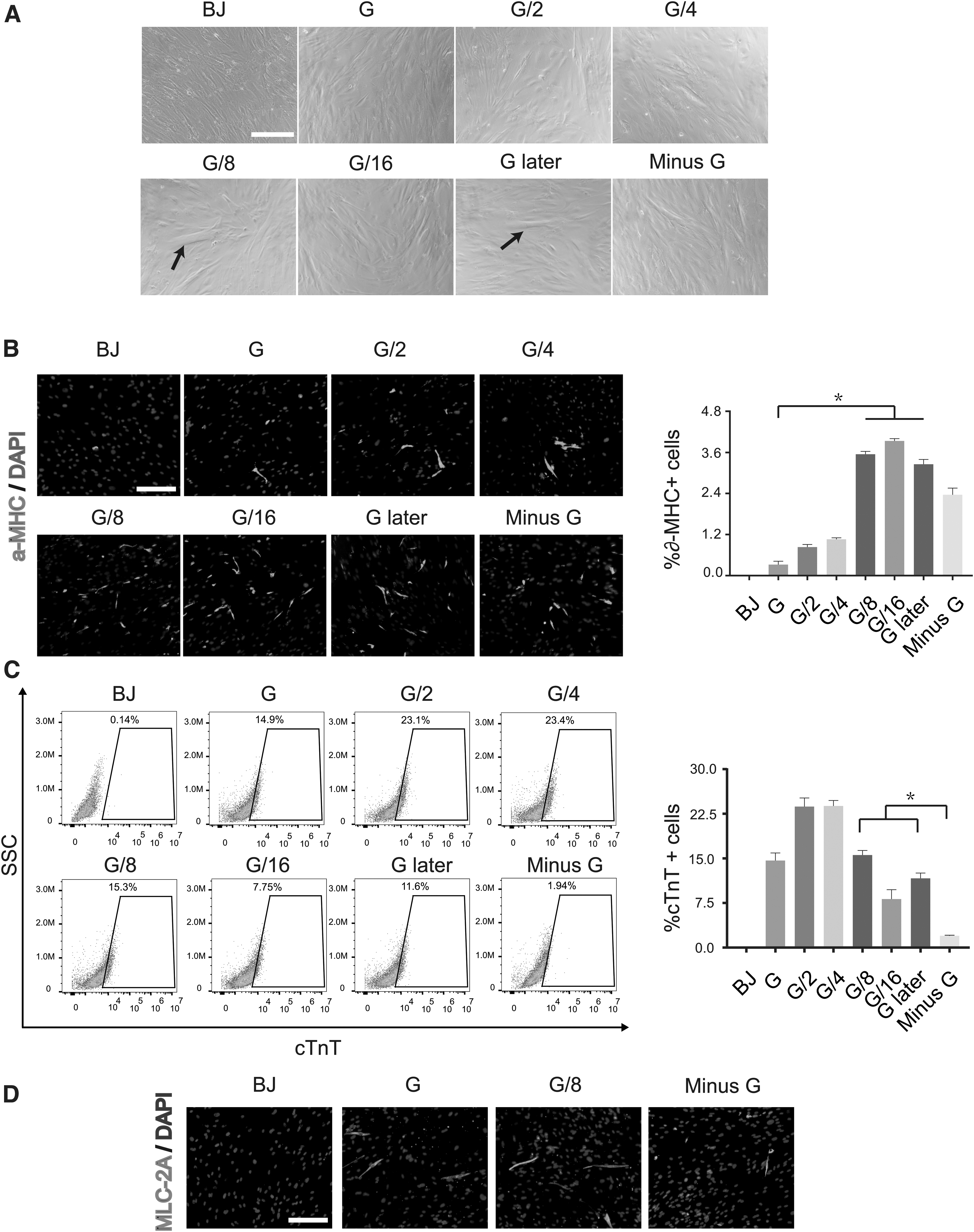
The effects of GATA4 dosage on cardiac reprogramming efficiency.
However, cTnT-positive iCMs was significantly lowered by removing GATA4 from GMTMeMy (1.94%) (Fig. 2C). Meanwhile, we also found that MLC2A-positive iCMs were also dramatically decreased after removal of GATA4 (Fig. 2D). These results showed that appropriate GATA4 dosage was important for the reprogramming efficiency.
Different GATA4 dosage results in varying numbers of functional iCMs
By cTnT and α-MHC double staining at late stage of reprogramming, We observed that there were more double positive cells in G/8 and G later groups, which were clearer and larger, and had characteristic stripes, while there were more single positive cells with smaller size and irregular shape in other groups (Fig. 3A). The induced cells did not show spontaneous contraction, although with prolonged culture. So the previous co-culture method (Zhou et al., 2019) was used to identify our iCMs, we labeled iCMs and BJ with DsRed. After 2 weeks of reprogramming, cells were replated onto neonatal mouse CMs. We found that beating colony appeared in G/4, G/8, G/16, and G later groups 1 week after co-culture and the numbers of beating loci per well in G/8 group and G later group were significantly higher than those in other groups (Fig. 3B and Supplementary Video S1).

The effects of GATA4 dosage on functional iCMs.
To further test the effects of GATA4 dosage on the functional properties of iCMs, calcium oscillation was performed by KCl stimulation 3 weeks after maintenance culture (Jayawardena et al., 2012; Nam et al., 2013). We found that compared with other groups, G/8, G/16, and G later groups had much more calcium oscillation-positive cells (Fig. 3C and Supplementary Video S2). These results demonstrated that there were more functional iCMs in G/8 and G later groups than those in other groups. For the sake of simplicity, we determine MTMeMy plus 1/8 GATA4 dosage as our reprogramming method.
iCMs were transcriptionally reprogrammed toward the CM state
To analyze time course of cardiac marker expression in our iCMs (G/8 group), qPCR was performed on day 0, 7, and 14. We uncovered that a panel of cardiac specific genes showed upregulated trend during the 2-week reprogramming period (Fig. 4). According to previous human cardiac cell atlas (Litvinukova et al., 2020), our qPCR analyses also showed that maker genes of cardiac smooth muscle cells (MYH11 and ACTA2) were obviously upregulated during reprogramming (data not shown).

Time course of cardiac gene expression profile in G/8 group. mRNA expression level of multiple cardiac genes in iCMs on day 0, 7, and 14. Gene expression was normalized to GAPDH. Data are mean ± SD (n = 3) unpaired t-test, *p < 0.05.
CMM accelerated the maturation of iCMs
Our iCMs robustly expressed plentiful CM genes at the later stage of reprogramming; however, they would not show typical calcium oscillation until 4 weeks after reprogramming, which indicated that mature iCMs need to experience a 2-week maintenance culture. To explore the method to accelerate the maturation of iCMs, CMM was prepared by CM differentiation of H9 hES cells (Fig. 5A). With the canonical protocol (Lian et al., 2012), we obtained contracting colony with strong immunostaining of cTnT and α-MHC on day 8 (Fig. 5B).
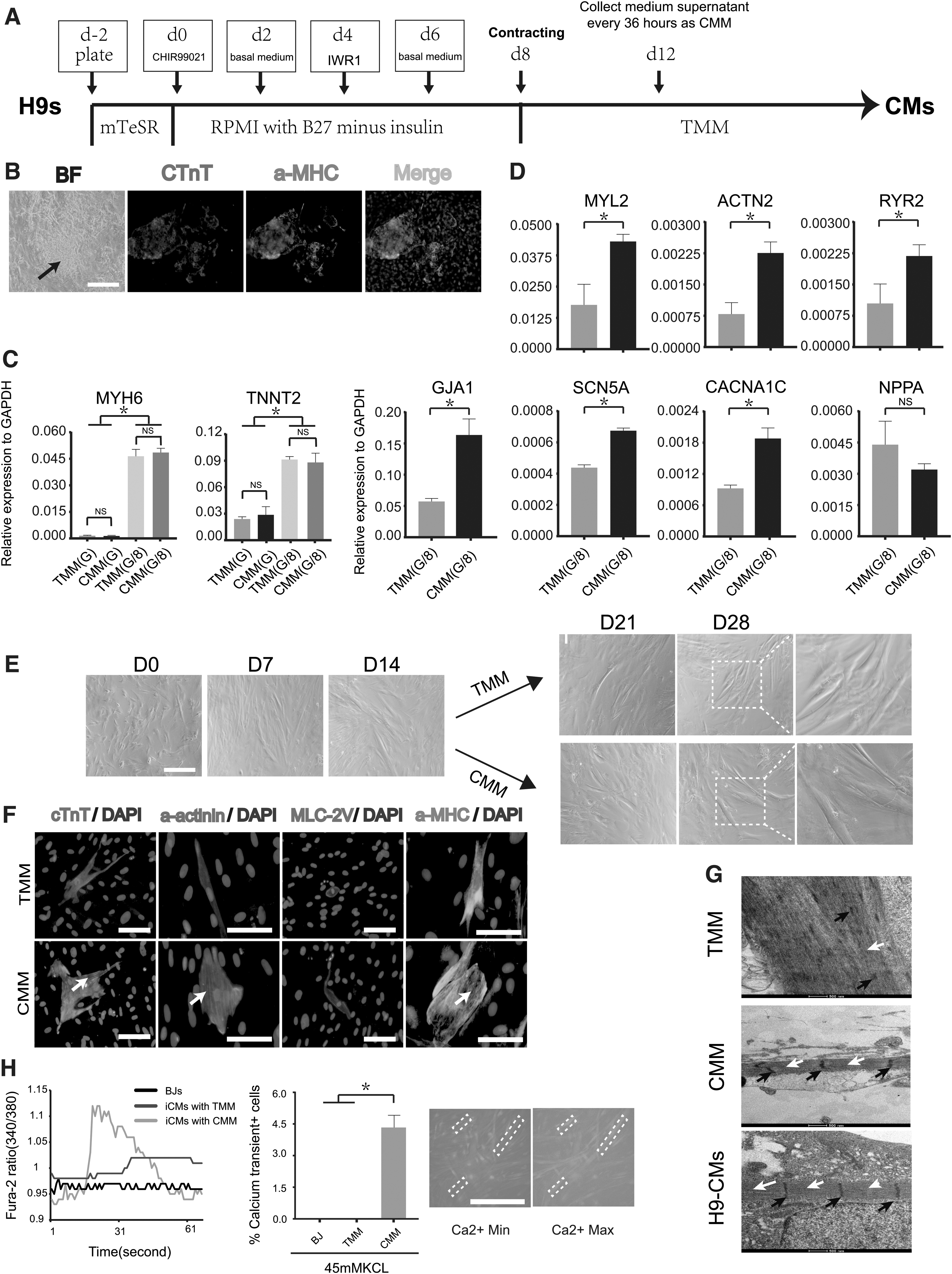
CMM accelerated maturation of iCMs.
We collected medium supernatant as CMM every 1.5 days from day 12, at which time, the whole plate showed fused beating colony. First, we analyzed the effects of CMM on expression of cardiac genes using qPCR. We found that compared with TMM treatment, CMM treatment did not affect TNNT2 and MYH6 in both G and G/8 group (Fig. 5C), but significantly increased expression level of MYL2, ACTN2, RYR2, SCN5A, CACNA1C, and SCN5A (Fig. 5D). After 2 weeks of maintenance culture, iCMs treated with CMM showed thicker morphology and clearer stripes than those treated with TMM (Fig. 5E). Immunostaining of different sarcomeric proteins revealed that compared with TMM group, CMM group showed a larger size and better-defined sarcomeric structures as early as 1 week after maintenance culture (Fig. 5F).
Transmission electron microscopy was performed to further determine whether CMM affected structural organization of iCMs. We found that after 3 weeks of maintenance culture, some iCMs in the CMM group showed myofibril bundles arranged in parallel, with relatively regular Z-line perpendicular to the myofilament and Z-lines divided myofibril bundles into several sarcomeres of the same length, which was similar to the H9-derived CMs. However, the arrangement of myofibrils and Z-line in TMM group was irregular (Fig. 5G). These results suggested that CMM facilitated structural maturation of iCMs.
To test the promotion of CMM treatment for functional property of iCMs, calcium oscillation was performed at different time after maintenance culture. We found that the CMM group showed characteristic wave of calcium oscillation only 1 week after maintenance culture but the TMM group could not (Fig. 5H), which indicated that CMM treatment accelerated functional maturation of iCMs. However, after several weeks of maintenance culture, the percentage of calcium oscillation-positive cells in both group differed a little (data were not shown), suggesting that CMM might not affect reprogramming efficiency.
Discussion
In this study, GMTMeMy were used to reprogram HDFs toward a CM-like state. We revealed the significant effects of GATA4 stoichiometry on cardiac reprogramming efficiency. We also found that CMM accelerated maturation of iCMs. Balanced factor combination and optimal culture microenvironment contributed to successful iCMs. Our finding provided new insights into early intrinsic and late extrinsic regulatory mechanisms of cardiac reprogramming.
GATA4 is the key regulator ranking at the top of transcriptional hierarchy of the cardiac gene manipulatory networks; unbalanced GATA4 dosage and inappropriate temporal expression of GATA4 result in damaged cardiac differentiation (Garg et al., 2003; Grepin et al., 1995; He et al., 2012; Molkentin et al., 1997; Tong, 2016). In studies of CM transdifferentiation, GATA4 is also an indispensible factor in both human and murine systems (Chen et al., 2012; Fu et al., 2013; Ieda et al., 2010; Nam et al., 2013; Wada et al., 2013).
Moreover, GATA4 stoichiometry in murine embryonic fibroblasts (MEFs) has been manipulated by polycistronic vectors to improve GMT-based cardiac reprogramming (Inagawa et al., 2012; Wang et al., 2015), and they found that relative lower GATA4 stoichiometry than MEF2C stoichiometry was correlated with higher reprogramming efficiency (Wang et al., 2015).
In our study, reprogramming HDF with the average pooled viruses encoding GMTMeMy resulted in a significantly higher GATA4 stoichiometry than MEF2C, MYOCD, or TBX5 stoichiometry, and the reprogramming efficiency was lower here than that described in previous studies (Wada et al., 2013), which reminded us an unreasonable exogenous stoichiometry of factors.
Precisely, we found that lowering GATA4 overexpression by reducing GATA4 dosage to 1/8 alone was sufficient to significantly promote reprogramming efficiency, although GATA4 protein level was not demonstrated by Western blot in each group in this study. Spontaneously beating in co-culture with murine neonatal CMs used to identify function of immature iCMs (Wada et al., 2013; Zhou et al., 2019). In this study, we found obvious beating loci in G/8 group, while no beating loci in G and G/2 group, which was in accordance with rare cTnT and α-MHC double immunostaining positive iCMs in G and G/2 group.
Later introduction of GATA4 could contribute to similar reprogramming outcomes with 1/8 GATA4 dosage, but we were not able to understand that the mechanism underlying the later introduction of GATA4 was only dosage dependent or time dependent just like the sequential introduction protocol (Oct4–Klf4 first, then c-Myc and finally Sox2) used to foster murine iPSC reprogramming (Liu et al., 2013). With such 1/8 GATA4 dosage method, our iCMs expressed a panel of critical CM-specific genes, although their expression level was still relatively lower compared with PSC-derived CMs. According to recent single-cell RNA-Seq analyses of the adult human heart (Litvinukova et al., 2020), maker genes of cardiac smooth muscle in our iCMs were also all upregulated, which may result from the role of MYOCD overexpression (Hoofnagle et al., 2011
In previous studies and our study, co-culture with murine neonatal CMs could make iCMs beat spontaneously without cell fusion, which suggested that animal CMs could help iCMs mature by secretion function. Moreover, cardiogenic compounds involved in CM differentiation of human PSCs were recently applied to convert partially reprogrammed iCMs into fully reprogrammed iCMs (Yamakawa et al., 2015). So we suppose that human CMs could also benefit iCMs by secreting some helpful factors during their maturation.
Because fetal CMs could not be obtained, we chose to differentiate H9 hES cells into CMs with high efficiency by manipulating WNT pathway, and prepared CMM from CM beating to complete maturation. We found that compared with TMM group, CMM group showed higher cardiac structural and functional gene expression level, better organized sarcomere, and earlier appear time of iCMs showing calcium oscillation, which demonstrated that the CMM treatment accelerated maturation of iCMs from all aspects.
However, in some batches of experiments, we also found that the function of CMM generated by H9-CMs with low differentiation efficiency was not obvious. So we think that H9-CMs introduce some active factors into the CMM, which may create a suitable microenvironment to accelerate the conversion of partially reprogrammed cells into functional iCMs. Unfortunately, in this study, we cannot point out one or some key factors working in CMM, which needs further study to dissect the difference in component between our TMM and CMM by screening for cytokines and growth factors. Meanwhile, our iCMs did not show spontaneous contraction to the end.
Compared with HCFs, HDFs are easier to obtain from healthy donors or patients to study the mechanism of cardiac reprogramming and establish an individualized cardiac disease model for pathogenesis research and drug screen. Moreover, reprogramming of HDFs can exclude the possibility of contamination with rare CPCs or CMs in HCFs and provide a more truly insight into cardiac reprogramming.
In this study, we only manipulated GATA4 stoichiometry to promote reprogramming, the phenomenon maybe only due to different susceptibility of HDFs to viruses encoding different factors. Further studies will be needed to precisely explore the most balanced stoichiometry using a polycistronic vector system and find the best introduction sequence of each factor. Stoichiometry of each factor should be appropriately determined by total protein level, in the future, to optimize this process. Meanwhile, it is critical to screen out more effective cardiogenic compounds to get functionally mature iCMs.
Footnotes
Acknowledgment
We thank Xiuzhi Bi for fund support.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
National Key Research and Development Program of China (2018YFA0108703). Also, the Research Innovation Fund of First Affiliated Hospital of Harbin Medical University, and National Natural Science Foundation of China (#81871501).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.