
Abstract
Mesenchymal stem cells (MSCs) exist in many tissues and can differentiate into cells of multiple lineages, such as adipocytes, osteoblasts, or chondrocytes. MSC administration has demonstrated therapeutic potential in various degenerative and inflammatory diseases (e.g., graft-vs.-host disease, multiple sclerosis, Crohn's disease, organ fibrosis, and diabetes mellitus [DM]). The mechanisms involved in the therapeutic effects of MSCs are multifaceted. Generally, implanted MSCs can migrate to sites of injury, where they establish an anti-inflammatory and regenerative microenvironment in damaged tissues. In addition, MSCs can modulate innate and adaptive immune responses through immunosuppressive mechanisms that involve immune cells, inflammatory cytokines, chemokines, and immunomodulatory factors. DM has a high prevalence worldwide; it also contributes to a high rate of mortality worldwide. MSCs offer a promising therapeutic agent to prevent or repair damage from DM and diabetic complications through properties such as multilineage differentiation, homing, promotion of angiogenesis, and immunomodulation (e.g., prevention of oxidative stress, fibrosis, and cell death). In this study, we review current findings regarding the immunomodulatory and regenerative mechanisms of MSCs, as well as their therapeutic applications in DM and DM-related complications.
Introduction
Mesenchymal stem cells (MSCs) have been successfully isolated and expanded from many tissues, such as bone marrow, adipose tissue, hair follicles, umbilical cord, placenta, amniotic fluid, and tooth pulp (Keating, 2012). Cultured MSCs comprise unit fibroblasts with a spindle morphology; they express multiple cell surface markers, such as CD29, CD90, CD73, CD105, CD51, and CD44. However, they do not express CD34, CD11b, CD31, CD45, CD19, HLA-DR, or other hematopoietic lineage markers. The stemness of MSCs is defined as the capacity for self-renewal and differentiation into a large number of lineages, including chondrocytes, osteoblasts, and adipocytes (Fig. 1). MSCs can be maintained for more than 20 passages without the loss of their main properties (Horwitz et al., 2005; Keating, 2012).
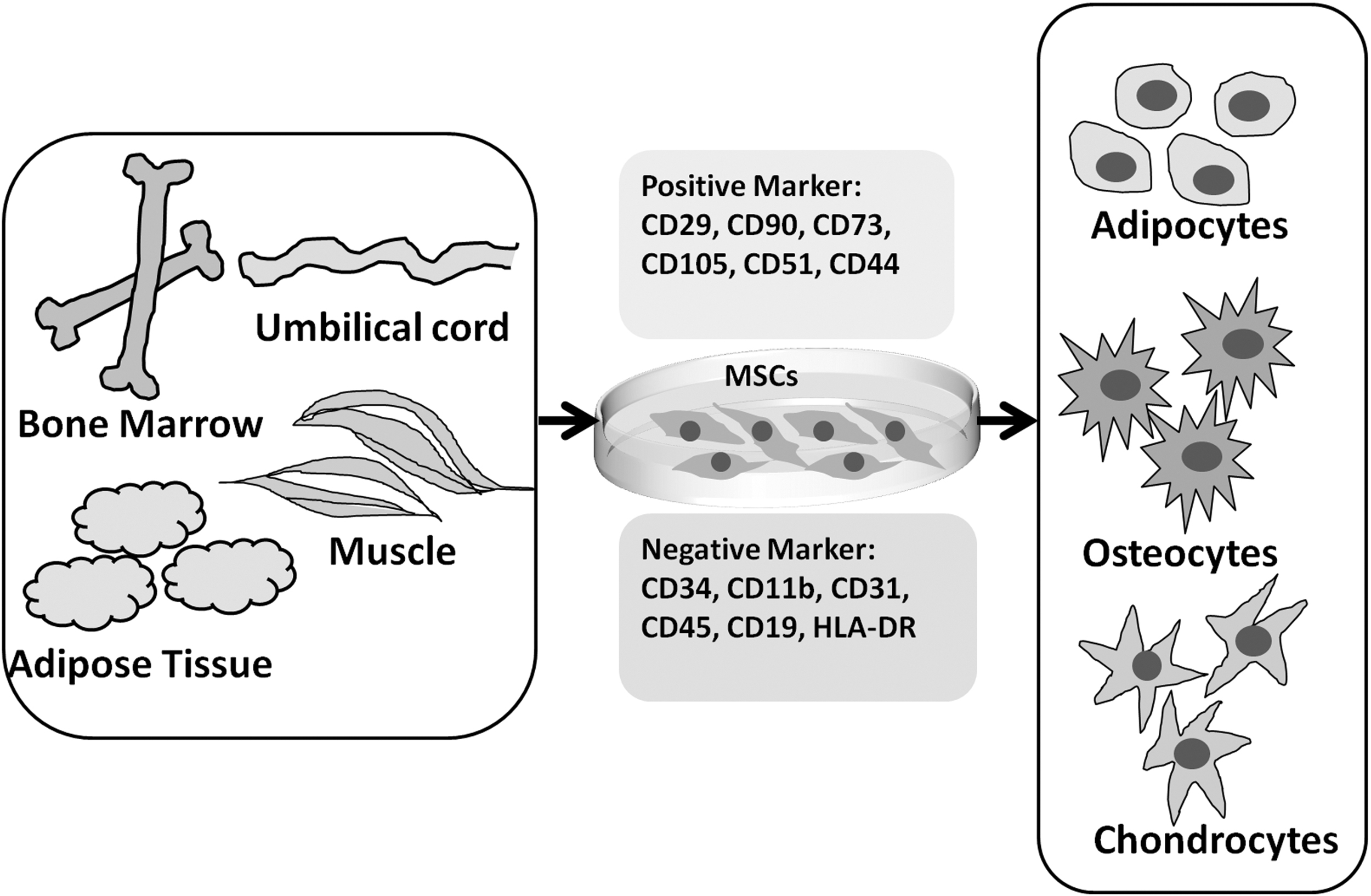
The biological properties of MSCs. MSCs can be isolated from adipose tissue, bone marrow, umbilical cord, and muscle. The expanded MSCs are characterized as unit fibroblasts with a spindle morphology and express a panel of surface markers. Moreover, MSCs possess the capability of differentiation into adipocytes, osteocytes, and chondrocytes. MSCs, mesenchymal stem cells.
MSC-based therapy has been recognized as a novel type of regenerative medicine because MSCs exhibit broad tissue distribution, multipotent differentiation, and well-established clinical efficacy; they have immunomodulatory and proangiogenic effects on various diseases (Shi et al., 2010).
Many animal and clinical studies have demonstrated the therapeutic efficacies of MSCs in multiple diseases (Ren et al., 2012). MSC administration can improve clinical status in patients with graft-versus-host disease (GvHD), cardiovascular diseases, and autoimmune diseases (Hare et al., 2012; Le Blanc et al., 2008; Sun et al., 2010; Tyndall and van Laar, 2010). The inflammatory tissue microenvironment varies among diseases treated with MSC-based therapy; the influence on MSCs also differs, leading to diverse efficacy outcomes (Krampera, 2011; Ren et al., 2008). Inflammatory status has also been shown to influence the clinical efficacy of MSC-based therapy.
Diabetes mellitus (DM) is a severe disease with high global prevalence and a high rate of mortality worldwide. Impaired insulin secretion and insulin resistance cause the accumulation of advanced glycation end products, which can enhance pathological vascular changes; such changes lead to various macrovascular complications (e.g., atherosclerosis, myocardial infarction, coronary artery disease, and peripheral arterial disease) and microvascular complications (e.g., retinopathy, neuropathy, and nephropathy) through increased oxidative and nitrosative stresses (Bernardi et al., 2012; Khamaisi and Balanson, 2017; Papatheodorou et al., 2016; Qiu et al., 2021).
These severe complications may lead to life-threatening organ and tissue dysfunction. Traditional therapies only focus on lifestyle modification (e.g., exercise and diet) and glucose management; they ignore the effects of diabetic complications (Peng et al., 2018; Wu and Mahato, 2014).
Pancreatic transplantation can improve difficulties; however, the use of this approach is limited by the small number of donors (Peng et al., 2018). Despite the donor shortage, pancreatic transplantation is associated with reductions in DM-related complications (Jenssen et al., 2017). MSC-based therapy is a promising treatment for DM and various complications because of its functional properties, such as multipotent differentiation and immune-suppressive capacity (Peng et al., 2018; Wu and Mahato, 2014). In this review, we discuss the mechanisms involved in MSC-based therapeutic efficacy and describe the current situation regarding therapeutic applications of MSCs in DM and DM-related complications.
Therapeutic Potential of MSCs
Large numbers of MSCs can be produced from various tissues (e.g., bone marrow, umbilical cord, and adipose tissues); such MSCs have clinical potential because of their extensive proliferative capacity (Horwitz et al., 2005). MSC-based therapy has been investigated in animal models of diseases such as GvHD, inflammatory bowel disease, DM, systemic lupus erythematosus, autoimmune encephalomyelitis, and collagen-induced arthritis (Fiorina et al., 2009; Polchert et al., 2008; Rafei et al., 2009; Sun et al., 2009). Importantly, the therapeutic efficacy of MSC administration has varied among animal disease models. Some studies have shown significant improvement after MSC treatment, whereas other studies have shown no improvement; in some instances, worse outcomes have been observed (Djouad et al., 2009; Sensebé and Bourin, 2009).
For example, MSC utilization in a collagen-induced arthritis mouse model did not provide any benefits after intravenous administration; in arthritis-induced tissues, however, damage was significantly improved after intraperitoneal injection of MSCs (Augello et al., 2007; Djouad et al., 2005). Diverse results were also observed in a mouse model of GvHD (Németh et al., 2009; Sudres et al., 2006). These discrepant findings might arise from many factors, such as MSC dose, administration route, and administration schedule, as well as recipient inflammation status.
MSCs are presumed to migrate into sites of tissue injury and participate in tissue repair (Shi et al., 2012). This process is associated with inflammatory cell activation and the release of various trophic factors, chemokines, and leukotrienes; these factors are precisely regulated to maintain tissue homeostasis in combination with the activities of fibroblasts and endothelial cells (Shi et al., 2012). There remains minimal information regarding the detailed mechanisms that are involved in migration, differentiation, and survival within damaged areas.
Furthermore, MSCs can suppress uncontrolled immune responses in sites of tissue injury and engage in negative regulation to ensure inflammatory homeostasis. Inflammatory factors produced in the microenvironments at sites of injury can stimulate the immunosuppressive capacity of MSCs (Ren et al., 2008, 2009). Therefore, inflammatory status should be considered when developing MSC-based therapeutic strategies in specific diseases. Later in this review, we will discuss the modulatory capacity of MSCs involved in therapeutic efficacy (Fig. 2).
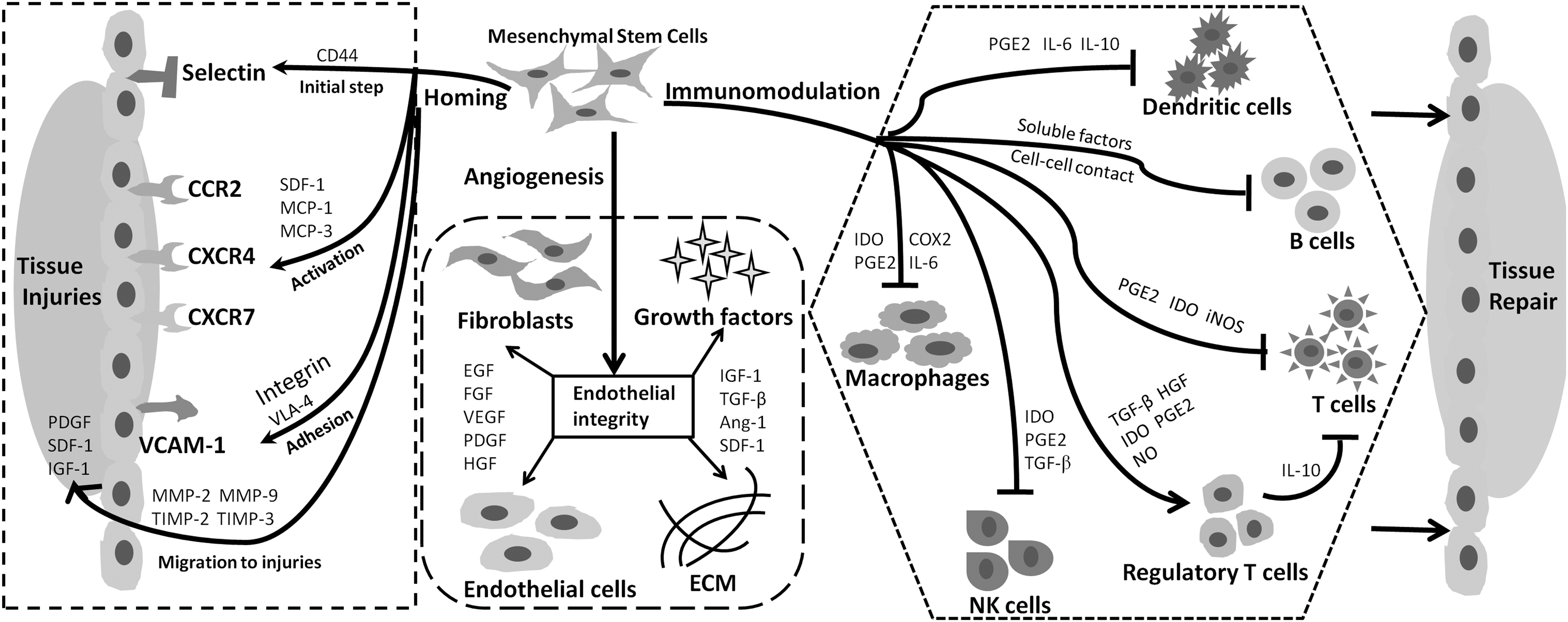
The mechanisms involved in the therapeutic potential of MSCs: homing property, angiogenic capacity, and immunosuppressive modulation. Ang-1, angiopoietin-1; CCR2, CC-chemokine receptor 2; CD44, cluster of differentiation 44; CXCR4, CXC-chemokine receptor 4; CXCR7, CXC-chemokine receptor 7; Cox2, cyclooxygenase-2; ECM, extracellular matrix; EGF, epidermal growth factor; FGF, fibroblast growth factor; HGF, hepatocyte growth factor; IDO, indoleamine 2,3-dioxygenase; IGF-1, insulin-like growth factor-1; IL-6, interleukin-6; IL-10, interleukin-10; iNOS, inducible nitric oxide synthase; MCP-1, monocyte chemoattractant protein-1; MCP-3, monocyte chemoattractant protein-3; MMP-2, matrix metalloproteinase-2; MMP-9, matrix metalloproteinase-9; NK cells, natural killer cells; NO, nitric oxide; PDGF, platelet-derived growth factor; PGE2, prostaglandin E2; SDF-1, stromal cell-derived factor-1; TGF-β, transforming growth factor-β; TIMP-2, tissue inhibitor metalloproteinase 2; TIMP-3, tissue inhibitor metalloproteinase 3; VCAM-1, vascular cell adhesion molecule-1; VEGF, vascular endothelial growth factor; VLA-4, very late antigen 4.
MSC homing to sites of injury
Fundamental properties of MSCs that influence their clinical efficacy include migration and homing to injured tissues with inflammation (De Becker and Riet, 2016). MSC transplantation can enhance interactions among immune cells, stromal cells, endothelial cells, and tissue fibroblasts, thus improving angiogenesis and tissue repair at sites of injury (Ren et al., 2008). Unlike leukocyte homing, the mechanisms that underlie MSC homing require further investigation.
When MSCs are locally transplanted to damaged tissue, they are directed to sites of injury via chemokine receptors; when MSCs are systemically administered, they undergo complex migration to target sites and ensure effective repair (Ullah et al., 2019). The initial steps of migration are regulated by selectins in endothelial cells and CD44 expression in MSCs. Interactions among these factors can promote MSC migration throughout the vasculature (Karp and Leng Teo, 2009; Sackstein et al., 2008). The subsequent activation step is mediated by G protein-coupled chemokine receptors, which are guided by inflammatory signals. The CXC-chemokine receptor 4 (CXCR4)–stromal cell-derived factor (SDF-1) axis has an essential role in this process; however, CXCR4 exhibits inconsistent effects on MSC migration in response to SDF-1 (Ip et al., 2007; Lau and Wang, 2011).
Other critical chemokine receptors on MSCs require further investigation to determine their effects on MSC homing (Honczarenko et al., 2006). The integrin B1 subunit can bind to very late antigen 4, which interacts with vascular cell adhesion molecule-1 to initiate migration arrest. Suppression of the integrin B1 subunit can inhibit MSC homing (Chang et al., 2007; Segers et al., 2006).
As described above, MSCs can cross the basement membrane, which is located beneath endothelial cells. Matrix metalloproteinase (MMP)-2 and MMP-9 degrade components of connective tissue (e.g., collagen and gelatin) to promote MSC mobility (Steingen et al., 2008; Toth et al., 2012); mobility is then regulated by tissue inhibitor metalloproteinase-3, tissue inhibitor metalloproteinase-2, and membrane type 1 MMP (Zhao et al., 2004). Finally, MSCs migrate to sites of injury in response to signals such as platelet-derived growth factor, insulin-like growth factor, SDF-1, and chemokines (Ullah et al., 2019).
Currently, there is a need to more fully elucidate the vascular effects, immune components, and tissue regeneration mediators associated with specific diseases (Fig. 2). Clarification of the interactions between MSCs and the regenerative microenvironment will facilitate the use of MSCs in the treatment of multiple diseases (Shi et al., 2018).
Regenerative properties of MSCs in damaged tissues
At sites of injury, the endothelial monolayer usually loses its barrier function in the capillary bed, thereby inducing the release of various chemokines and leukotrienes into the blood. These inflammatory factors induce MSC recruitment and stimulate the production of various trophic factors, such as epidermal growth factor, fibroblast growth factor, vascular endothelial growth factor (VEGF), platelet-derived growth factor, hepatocyte growth factor (HGF), insulin-like growth factor-1, transforming growth factor-β (TGF-β), angiopoietin-1, and SDF-1 (Ma et al., 2013; Ren et al., 2008, 2009; Shi et al., 2012). These paracrine factors help to rebuild endothelial integrity and improve angiogenesis by modulating endothelial cell development and extracellular matrix production; they also prevent leukocyte-mediated effects on endothelial cells (Lee et al., 2011; Ma et al., 2013, 2014).
Engrafted MSCs can support the functional recovery of damaged tissues and organs by differentiating into tissue-specific progenitors or promoting tissue reestablishment (Lee et al., 2011; Shi et al., 2012; Volarevic et al., 2011) For example, MSC-secreted HGF effectively regulates the remyelination of endogenous neural cells, thus enhancing MSC-based therapeutic efficacy in models of autoimmune encephalomyelitis and spinal cord demyelination (Bai et al., 2012). Overall, these results suggest a complex tissue repair mechanism, in which MSCs and secreted trophic factors improve the function of tissue-resident progenitor cells and promote repair (Klimczak and Kozlowska, 2016; Yu et al., 2020).
Immunomodulatory effects of MSCs on tissue repair
MSCs exhibit immunoregulatory effects through interactions with inflammatory and immune components (Le Blanc and Mougiakakos, 2012). Macrophages are important mediators in the initiation and management of inflammation. The proinflammatory cytokines produced by M1 macrophages or T cells can stimulate MSCs to secret mediators that direct monocyte differentiation into anti-inflammatory M2 macrophages (Le Blanc and Mougiakakos, 2012). M2 macrophages can produce anti-inflammatory cytokines (e.g., interleukin [IL]-10 and TGF-β1) that help to improve tissue repair after inflammation (Mantovani et al., 2013). Interactions between MSCs and macrophages enhance the MSC capacity to trigger the transition from proinflammatory to anti-inflammatory activities (Bernardo and Fibbe, 2013).
MSCs reportedly exhibit immunomodulatory potential at sites of tissue injury (Le Blanc and Mougiakakos, 2012). For example, MSCs can suppress the expression of MHC class II, CD1a, CD40, CD80, and CD86, thereby producing immature dendritic cells (Jiang et al., 2005). In addition, MSCs can suppress T cell proliferation activated by certain stimuli, such as prostaglandin E2 (PGE2), TGF-β, HGF, and heme oxygenase (Stagg and Galipeau, 2013). Regulatory T cells (Tregs) can suppress T cell-mediated immune responses. Several studies have shown that MSCs can promote the differentiation of CD4+ T cells into Tregs. This polarization of T cells toward a Treg phenotype is regulated by direct MSC contact with helper T cells, as well as the secretion of PGE2 and TGF-β (Burr et al., 2013; English et al., 2009).
Notably, the MSC-mediated inhibition of T lymphocyte proliferation and Treg production is monocyte-dependent (Cutler et al., 2010; François et al., 2012). Monocyte depletion impedes T cell differentiation toward a regulatory phenotype, while monocyte supplementation restores this process. Overall, MSCs can improve the balance between proinflammatory T cells and anti-inflammatory Tregs through a process associated with monocyte polarization toward M2 macrophages, which possess anti-inflammatory characteristics (Akiyama et al., 2012; François et al., 2012; Melief et al., 2013).
The results of clinical studies have suggested that proinflammatory cytokines induce immunosuppressive functions in MSCs, while the absence of these cytokines can lead to immune enhancement after MSC administration. Therefore, the maintenance of sufficient proinflammatory cytokine levels before MSC administration can promote immunosuppressive efficacy (Wang et al., 2014). For example, cytokine pretreatment can improve the therapeutic efficacy of MSCs in an animal model of acute myocardial ischemia (Luo et al., 2012). Moreover, the administration of MSCs pretreated with interferon-γ can significantly improve survival in mice with GvHD (Polchert et al., 2008).
Similarly, MSCs conditioned with interferon-γ and tumor necrosis factor-α (TNF-α) demonstrate better efficacy in a model of concanavalin-A-induced hepatitis. Inflammation can enhance the immunomodulatory activity of MSCs at sites of tissue injury; recipient inflammatory status should be considered because the immunomodulatory profile of the administered MSCs might change during the use of immunosuppressants (Shi et al., 2018). Some studies have shown that immunosuppressants (e.g., dexamethasone) reduce the expression levels of inducible nitric oxide synthase and indoleamine 2,3-dioxygenase, which can lead to the failure of MSC-based therapy (Chen et al., 2014). Specific disease-related inflammatory statuses can influence the immunosuppressive properties of MSCs.
The spread of COVID-19 has become a global public health crisis; the disease can induce a hyperinflammatory state or cytokine storm, leading to acute respiratory distress syndrome and multiorgan damage. Kassem and Kamal (2020) summarized the efficacy of MSCs in the treatment of COVID-19. Because of their immunomodulatory properties, the administration of MSCs can ameliorate the cytokine storm that occurs in COVID-19 patients. Moreover, MSCs exert regenerative and antifibrotic effects in infected lungs, where they promote tissue repair and regeneration (Kassem and Kamal, 2020). MSCs produce PGE2 and IL-10, which can reduce inflammatory cytokine release, inhibit M1 macrophage activity, and block neutrophil recruitment to sites of lung injury (Devaney et al., 2015; Taghavi-Farahabadi et al., 2020).
In a phase I clinical trial involving seven COVID-19 patients, MSC treatment improved disease symptoms and pulmonary function without adverse effects. Natural killer cells disappeared and TNF-α release was reduced, while the release of IL-10 (a well-known inhibitory cytokine) was increased after MSC administration; these findings suggest that MSCs can produce anti-inflammatory effects in COVID-19 patients (Leng et al., 2020).
Thus, the immunosuppressive functions of MSCs can be influenced by many aspects, such as the source of MSCs, the number of passages before MSC injection, the dose of MSCs, and the disease-specific inflammatory status of the recipient. Mechanisms related to interpretation of the immunomodulatory properties of MSCs continue to receive substantial attention from many primary and clinical researchers.
As stated above, the combination of immunomodulatory and tissue regenerative potential of MSCs remains a scientific and clinical highlight, and MSC-based therapy is recognized as a promising treatment for DM (Fig. 3).
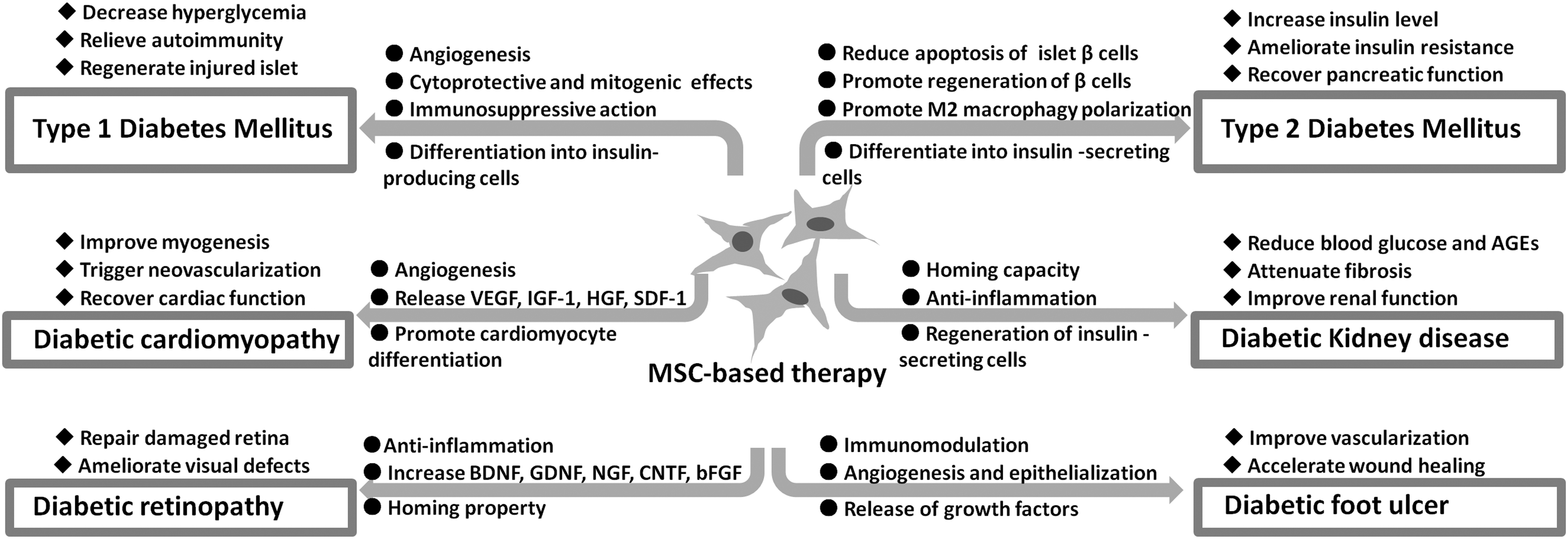
A schematic model demonstrating the biological properties of MSCs and potential therapeutic mechanisms involved in cell therapies for diabetes and associated complications. BDNF, brain-derived neurotrophic factor; bFGF, basic fibroblast growth factor; CNTF, ciliary-derived neurotrophic factor; GDNF, glial cell-derived neurotrophic factor; NGF, nerve growth factor.
Therapeutic Potential of MSCs in Type 1 DM
Type 1 DM (T1DM) is a chronic autoimmune disease with a high prevalence worldwide (Monaghan et al., 2015). Pathological changes in T1DM primarily comprise dysfunctional activity involving insulin-secreting beta cells, which leads to autoimmune destruction and hyperglycemia (Burrack et al., 2017). Traditional primary therapy to control glucose levels through insulin injection usually becomes ineffective during long-term treatment. Islet transplantation has limited applicability because of problems such as insufficient pancreatic cell availability, risk of immune rejection, and the use of immunosuppressive drugs (Li and Ikehara, 2014; Nair et al., 2020). MSC-based therapies may overcome these difficulties because they have low immunogenicity and exhibit immunosuppressive properties.
MSCs are potential candidates for T1DM treatment because of their anti-inflammatory potential and multipotent differentiation capacity (Peng et al., 2018). The immunomodulatory properties of MSCs can improve the balance between destruction and regeneration of insulin-secreting beta cells. In addition, MSCs differentiate into insulin-producing cells and improve the repair of damaged pancreatic tissue; these abilities can be applied to the treatment of T1DM (Päth et al., 2019).
Bone marrow mesenchymal stem cells (BMSCs) can promote graft transplantation and reduce immunorejection; moreover, transplanted BMSCs travel to sites of injury in the pancreas, where they induce endogenous beta cell proliferation and islet regeneration to improve islet function and vascular regeneration (Bell et al., 2012; Chhabra and Brayman, 2013; Ren et al., 2012). Adipose-derived mesenchymal stem cells (ADSCs) have also received clinical research attention because they can enhance tissue regeneration and inhibit inflammation. ADSCs have been successfully used to treat T1DM and prevent graft rejection (Chandra et al., 2011). In a seminal study, MSCs were able to restore the balance of Th1/Th2 cytokines, increase the proliferation of CD4+CD25+Foxp3+ Tregs, and induce the infiltration of apoptotic leukocytes in pancreatic islets (Zhao et al., 2009).
However, the DM microenvironment (including hyperglycemia, chronic inflammation, and oxidative stress) has various effects on the biological properties and functions of MSCs. Davies et al. (2016) investigated the effects of T1DM on MSC biology and functionality; they found comparable potency in autologous MSCs derived from T1DM donors and healthy controls, in terms of growth kinetics and colony-forming capacity. Despite differences in gene expression between the two types of MSCs, significant differences were not observed in terms of immunosuppressive potential, migratory capacity, or hemocompatibility. Moreover, transplantation of autologous MSCs into DM patients led to improvements in C-peptide and HbA1c levels, indicating that MSC-based therapy is effective for the treatment of T1DM (Davies et al., 2016).
El-Badawy and El-Badri (2016) assessed the safety and therapeutic efficiency of various MSC types based on data from several clinical trials. They found that the transplantation of umbilical cord-derived MSCs led to better outcomes in T1DM patients, compared with transplantation of bone marrow-derived MSCs. Moreover, the administration of MSCs at an early stage of T1DM was more effective than intervention at later stages (El-Badawy and El-Badri, 2016).
Thus far, MSCs have demonstrated robust efficacy in both preclinical and clinical studies of T1DM treatment. However, further investigations are needed to explore the interactions between T1DM disease characteristics and therapeutic properties of MSCs. Such findings will help to characterize the efficacy of autologous MSC-based therapy for T1DM.
MSCs-Based Cell Therapy for T2DM
T2DM is caused by reduced secretion of insulin and the onset of insulin resistance. Conventional therapeutic approaches include insulin injection and oral administration of antidiabetic drugs (Zang et al., 2017). However, long-term disease leads to progressive loss of islet function. Because of its immunomodulatory properties and proangiogenic capacity, MSC-based therapy is potentially useful for the treatment of T2DM.
MSCs (e.g., BMSCs and ADSCs) differentiate into insulin-secreting cells, which increase insulin levels in patients after successful implantation (Bani Hamad et al., 2021; Chen et al., 2020; Pagliuca et al., 2014; Zhang et al., 2020). Under hyperbaric oxygen conditions, intra-arterially injected autologous BMSCs can regulate glycemic status and promote insulin secretion (Bhansali et al., 2014). In addition, MSCs can restore islet function and suppress insulin resistance in T2DM patients (Pagliuca et al., 2014). Notably, MSCs can differentiate into insulin-producing cells and aid in the regeneration of beta cells; intrapancreatic MSC transfusion leads to the attenuation of insulin resistance and recovery of pancreatic function, as well as an increase in the fasting C-peptide level, which is an essential indicator of beta islet cell function (Zang et al., 2017).
MSCs have sufficient regenerative capacity to restore pancreatic islet beta cells by homing to sites of injury and secreting certain cytokines (e.g., IL-6, SDF-1, granulocyte colony-stimulating factor, and granulocyte-macrophage colony-stimulating factor) (Caplan and Dennis, 2006). The coadministration of umbilical cord MSCs with liraglutide leads to protective effects on beta cells in terms of reduced apoptosis (Wang et al., 2020). Moreover, MSCs can enhance insulin secretion by promoting beta cell regeneration (Guney and Gannon, 2009). M2 macrophages can improve insulin resistance by reducing the levels of proinflammatory cytokines. MSCs have been shown to promote M2 macrophage polarization and directly reduce insulin resistance (Fujisaka et al., 2009).
In addition, some studies have focused on the effects of T2DM on MSC properties. ADSCs derived from T2DM donors are more likely to exhibit impaired cell replication, increased senescence and apoptosis, and abnormal levels of adipokines (e.g., adiponectin and resistin), compared with ADSCs derived from nondiabetic donors (Cramer et al., 2010). ADSCs isolated from ischemic limbs of DM patients are less robust in terms of phenotype and function, compared with ADSCs isolated from nondiabetic individuals who lacked signs of limb ischemia (Kočí et al., 2014).
In a clinical setting, Skyler et al. (2015) assessed the safety and efficacy of allogeneic MSCs in the treatment of T2DM, using a randomized placebo-controlled trial design. They found that the HbA1c level was reduced in patients who underwent administration of MSCs, which indicated that MSCs could aid in the treatment of T2DM. Packham et al. (2016) assessed the safety, tolerability, and efficacy of allogeneic bone marrow-derived MSCs in diabetic kidney disease (DKD). Compared to placebo, the administration of bone marrow-derived MSCs tended to stabilize or improve estimated glomerular filtration rate (eGFR). Moreover, no acute adverse events were observed in either of these two clinical trials, indicating that MSC treatment is safe.
This section has summarized potential applications of MSCs in the treatment of T2DM; robust efficacy has been observed in both preclinical and clinical studies. However, the mechanism that underlies effective MSC-based therapy for T2DM.
Applications of MSCs in Pancreatic Islet Transplantation
Human islet transplantation is a promising treatment for DM; however, grafted islets are affected by blood-mediated inflammation, acute rejection, ischemia associated with poor vascularity, and islet toxicity. Recent studies have shown that MSCs can improve pancreatic islet transplantation outcomes and overcome important challenges associated with islet transplantation. Upon cotransplantation with pancreatic islets, MSCs exhibit immunomodulatory and proangiogenic effects.
The immunosuppressive capacity of MSCs is sufficient to mitigate inflammation-related damage to transplanted islets. For example, in a mouse model of islet cell and BMSC coadministration, MSCs were able to reduce autoimmunity by blocking T cell proliferation and inhibiting both the differentiation and maturation of dendritic cells; these effects led to decreased blood glucose levels and increased serum insulin levels (Katuchova et al., 2015).
Notably, MSCs can secrete important trophic factors that aid in the recovery of islet vascularization and graft function in transplanted islets (Figliuzzi et al., 2014). Moreover, in a mouse model of islet cell and MSC coadministration, greater numbers of new peri-islet vessels and higher VEGF expression levels were observed, compared with mice that had received islet cells alone (Sakata et al., 2010). These results suggest that the coadministration of BMSCs with islets can improve islet vascularization and functional recovery after implantation because of the immunosuppressive and proangiogenic properties exhibited by BMSCs.
MSC-Based Therapy for Diabetic Complications
Diabetic cardiomyopathy
Diabetic cardiomyopathy is a severe complication of DM, which is characterized by cardiomyocyte hypertrophy and apoptosis, as well as increased collagen deposition caused by hyperglycemia-induced ventricular dysfunction in the heart (Volarevic et al., 2011; Yoon et al., 2005). MSCs can improve angiogenesis and myogenesis by releasing growth factors such as VEGF, insulin-like growth factor-1, and HGF (Yoon et al., 2005; Zhang et al., 2008). In a rat model of diabetic cardiomyopathy, BMSCs aided in cardiac functional recovery by promoting cardiomyocyte differentiation and improving both myogenesis and angiogenesis (Cai et al., 2015; Zhang et al., 2008). Furthermore, MSCs secreted the hypoxia-related factors HOM-1, SDF-1, and VEGF under hypoxic conditions; these factors triggered neovascularization and the restoration of myocardial function (Cai et al., 2015; Kourembanas, 2015).
MSCs have been implicated in cardiomyocyte regeneration and cardiac recovery. The optimal routes and sites of MSC injection require further investigation to ensure the effectiveness of MSC-based therapy for myocardial infarction.
Diabetic kidney disease
DKD is a major cause of chronic renal disease (Liu and Tang, 2016; Nagaishi et al., 2016); it is characterized by podocyte loss, increased proteinuria, enhanced fibrosis, and eventual renal failure. Damaged podocytes impede glomerular function and cause severe proteinuria. Proteinuria can enhance fibrosis and tubulointerstitial inflammation, leading to impaired renal activity. The increased levels of advanced glycation end products, glycated albumin, and blood glucose in DM can interfere with the functions of proximal tubular epithelial cells (Peng et al., 2018).
The administration of MSCs to rats with streptozotocin-induced DM leads to the regeneration of insulin-secreting cells, thus restoring renal function (Ezquer et al., 2008, 2012). Notably, SDF-1 release improves MSC homing capacity during renal damage repair (Wu et al., 2014). BMSCs can inhibit the release of the proinflammatory factor TGF-β1, ameliorate tubular interstitial fibrosis, and reverse apoptotic damage in tubular interstitial cells; these changes indicate improvements in DKD (Nagaishi et al., 2016). MSCs also reduce fibrosis and glomerulosclerosis via podocyte restoration, thus producing regenerative and protective effects in renal tissues damaged by DM (Ezquer et al., 2015). BMSC administration helps to regulate insulin, heme oxygenase-1, advanced glycation end products, and glucose, thereby reversing renal damage (Hamza et al., 2017).
Zhou et al. (2009) showed that MSC transplantation could attenuate DKD in a rat model by reducing blood glucose, albumin/creatinine (Alb/Cr) ratio, and renal mass. Ebrahim et al. (2018) reported that MSC-derived exosomes could significantly improve renal function and reduce the expression levels of fibrotic markers in renal tissue; these findings indicated substantial amelioration of streptozotocin-induced DKD.
Hickson et al. (2021) summarized the therapeutic outcomes of MSC-based intervention in experimental DKD. Renal function was improved, as demonstrated by reductions in markers of kidney injury. Notably, renal function improvement was greater when MSCs originated from bone marrow or umbilical cord/amniotic fluid than when MSCs originated from adipose tissue or other tissues. In addition, the renal outcomes of MSC-based therapy were influenced by tissue source, donor health, MSC dose, and administration route (Hickson et al., 2021).
Multiple studies have assessed the clinical outcomes of MSC transplantation as treatment for DKD. Some investigations revealed positive outcomes in animal models of DKD. However, the efficacies of MSC-based therapies for DKD require further exploration, particularly in terms of the appropriate dose and timing of MSC injection, as well as quantifiable indicators of therapeutic response.
Diabetic retinopathy
Diabetic retinopathy (DR) is a common complication of DM characterized by abnormal ocular vascularity and retinal lesions leading to blindness (Cheung et al., 2010; Wong et al., 2016). Conventional therapies such as laser photocoagulation and vitrectomy do not resolve the cause of the disease (Ciulla et al., 2003). Thus, MSC-based therapies have been considered for DR.
MSCs can differentiate into ocular cells, thereby promoting regeneration and repair in damaged retina (Davey et al., 2014). MSCs have also been shown to mitigate and restore visual impairments (Inoue et al., 2007; Wang et al., 2010). Some neurotrophic factors (e.g., brain-derived neurotrophic factor, nerve growth factor, ciliary-derived neurotrophic factor, basic fibroblast growth factor, and glial-derived neurotrophic factor) are upregulated in animal models of DR after injection with MSCs; this upregulation confirms the neuroprotective activity of MSCs in DR (Scalinci et al., 2011). In a therapeutic investigation, CD140+ ADSCs ameliorated DM-related visual defects (Elshaer et al., 2018). Despite the prospective applications of MSC-based therapy for DR, there remain many barriers to its clinical usage.
Diabetic foot ulcer
Diabetic foot ulcer (DFU) is a severe complication with high prevalence among DM patients. Current therapy is hindered by the lack of effective methods to heal deep chronic wounds and microvascular obstructions. DFU management is often impeded by factors such as infection, peripheral neuromuscular disease, hypoxia, and neuropeptide conduction (Xiong et al., 2021). MSCs constitute a promising therapeutic option to repair tissue damage and improve wound healing through enhanced epithelialization and angiogenesis. MSCs secrete various factors with immunomodulatory and vascular effects (An et al., 2021). Li et al. (2020) reported that the MSC-derived exosomal lncRNA H19 improved wound healing in a DFU model through the microRNA-152-3p-mediated enhancement of PTEN expression.
In addition, ADSCs were protective against DFU in diabetic rats. Enhanced angiogenesis, cellular proliferation, and growth factor release were observed in wound beds after ADSC treatment, suggesting that ADSCs are suitable for clinical treatment of DFU (Shi et al., 2016). Furthermore, exosomes from Nrf2-overexpressing ADSCs accelerated cutaneous wound healing by improving vascularization in a DFU rat model (Li et al., 2018). MSCs have also shown efficacy in DFU treatment when cotransplanted with autologous skin fibroblasts (Vojtassák et al., 2006). Similar results have been reported when DFU treatment was performed using MSCs derived from placenta, bone marrow, and human umbilical cord (Lopes et al., 2018).
Impacts of Allogeneic- or Autologous-Derived MSCs on DM
Allogeneic- or autologous-derived MSCs have distinct effects when used in the treatment of DM. Allogeneic MSCs elicit an immune response when rejected by the recipient's immune system, despite their low immunogenicity (Griffin et al., 2013). Autologous MSC-based therapy may be safer because MSCs are extracted from a patient, then returned to the same patient; this avoids unwanted immune responses and graft rejection (Sávio-Silva et al., 2020). However, it remains unclear whether autologous or allogeneic MSCs should be used in the treatment of DM (Sávio-Silva et al., 2020).
Chronic inflammation and hyperglycemia in DM may impair the MSC microenvironment and preclude the use of autologous MSCs. Some studies of MSCs have shown that hyperglycemia can lead to senescence and apoptosis, as well as diminished proliferation and osteogenic differentiation (Mahmoud et al., 2019). Other studies have demonstrated altered function in diabetic MSCs, which may cause reduced efficacy and interfere with therapeutic outcomes (Mahmoud et al., 2019).
Notably, the type of DM may affect biological properties of MSCs. MSCs derived from T1DM patients showed comparable properties in terms of viability, proliferation, multipotency, immunomodulation, and migration. In contrast, MSCs from T2DM patients exhibited severe senescence and apoptosis, along with lower proliferative potential and proangiogenic capacity (Cramer et al., 2010; Karina et al., 2019). BMSCs from T1DM patients have similar morphology, multilineage differentiation, and immunosuppressive capacity, compared with MSCs from healthy individuals (Yaochite et al., 2016). ADSCs from T2DM patients exhibit reduced proliferation, enhanced senescence, and altered CD105 expression; these manifestations suggest that angiogenesis is impaired (Karina et al., 2019; Marycz et al., 2016; Shin and Peterson, 2012).
Thus, it is important to assess intrinsic MSC abnormalities to avoid adverse effects on the efficacy of MSC-based therapy for DM (Sávio-Silva et al., 2020).
Clinical trials have demonstrated that both autologous and allogeneic MSCs are safe and effective in the treatment of DM (Sávio-Silva et al., 2020). However, some critical aspects of preclinical and clinical approaches require further attention, such as increased monitoring and follow-up of immune responses, reduction of alloantigen expression, selection of optimal tissue sources, and identification of appropriate administration routes (Lohan et al., 2017). Such information will help to assess the efficacy of MSC-based therapy and develop more precise criteria for DM patients who should receive this treatment.
Effects of MSC-Derived Exosomes on DM
Exosomes are small extracellular vesicles with membranes that can mediate signal transduction through the delivery of noncoding RNA, protein, and DNA. MSC-derived exosomes have become a focus of tissue repair research in the context of diabetic complications because of their cell-free properties, particularly in terms of regeneration and immunosuppression (Jiang et al., 2016; Marrazzo et al., 2014). Human MSC-derived exosomes can alleviate DM by reducing blood glucose levels, reversing insulin resistance, and relieving beta cell destruction (Sharma et al., 2021; Sun et al., 2018). MSC-derived exosomes ameliorated peripheral neuropathy in a mouse model of DM by increasing blood vessel density and the number of intraepidermal nerve fibers (Fan et al., 2020).
Human umbilical cord MSC-derived exosomes helped to reduce T2DM severity by improving hepatic glycolysis, glycogen storage, and lipid metabolism through activation of the autophagy pathway (He et al., 2020). In another study, MSC-derived exosomes improved renal function in a model of DKD through the induction of autophagy (Ebrahim et al., 2018). Overall, the findings in previous studies suggest that MSC-derived exosomes can be used in the treatment of DM and diabetic complications (Cantaluppi et al., 2012; Zhang et al., 2018).
The dose and quantity of exosomes should be assessed in preclinical and clinical studies to facilitate clinical application (Pachler et al., 2017; Watson et al., 2016); the route of exosome administration should also be clarified. Other remaining challenges include extraction protocol simplification, purity enhancement, and cost reduction (Castaño et al., 2019). In addition, because diverse substances are contained in MSC-derived exosomes, the therapeutic mechanisms and signaling pathways of such exosomes require further investigation.
Clinical Applications of MSC-Based Therapy for DM
Because MSCs have shown substantial potential for the treatment of DM, many clinical trials have been conducted. In May 2022, the Clinical Trials database (ClinicalTrials.gov) contained information regarding 38 clinical trials of MSC-based therapy for DM (12 T1DM-focused trials, 10 T2DM-focused trials, and 16 diabetic complication-focused trials). Both autologous and allogeneic MSCs have been used in the registered trials; the types of MSCs have included BMSCs, ADSCs, umbilical cord-derived MSCs, and placenta-derived MSCs. In one clinical trial, the administration of autologous BMSCs reduced the levels of antipancreatic islet antibodies, blood glucose, and HbA1c, while increasing the level of C-peptide (Mesples et al., 2013).
In a trial of newly diagnosed T1DM patients, the administration of autologous BMSCs led to increased C-peptide levels (Carlsson et al., 2015). Multiple trials have suggested that MSC-based therapy can improve beta cell function in T1DM (Gan et al., 2018). Several trials of patients with T2DM have suggested that MSC-based therapy can increase both stimulated and fasting levels of C-peptide (Bhansali et al., 2009, 2017; Hwang et al., 2019; Jiang et al., 2011).
In addition, concerning MSC administration routes and doses, Kabat et al. (2020) analyzed data from 914 clinical trials. They found that intravenous injection was the most common and reproducible method; importantly, robust efficacy was achieved with 100–150 million MSCs per patient. Lalu et al. (2012) assessed the safety profile of MSC-based therapy through a systematic review of adverse events that occurred during clinical trials. While they did not detect associations between MSC-based therapy and severe adverse events (e.g., acute toxicity, infection, organ complications, death, and malignancy), they found a significant association between MSC-based therapy and transient fever (Lalu et al., 2012).
Multiple promising preclinical studies have led to phase I/II clinical trials of MSC-based therapy for DM. However, such therapy cannot be widely used by clinicians because various concerns remain. For example, undifferentiated MSCs are suspected to undergo mutation and cell transformation during long-term culture (Tolar et al., 2007). Another concern regarding MSC-based therapy for DM is the lack of an effective administration route. Approximately 70% of intravenously injected MSCs do not migrate beyond the lungs; thus, few MSCs reach the heart, liver, and kidney (Zhang et al., 2015). Overall, clinical trials provide important insights concerning the overall understanding of MSC-based therapy.
Limitations of MSC-Based Therapy
In many studies, MSCs have demonstrated low immunogenicity and good tolerability; thus, MSC-based therapy is promising and has received intense research interest. However, several concerns remain regarding the use of MSCs. Contamination should be avoided during large-scale manufacturing of MSCs; MSC heterogeneity is another important challenge, such that stem cell properties must be maintained without promoting tumorigenicity or immunogenicity.
The transplantation of short-term cultured MSCs has led to malignant tumor formation in experimental models of myocardial infarction and diabetic neuropathy (Jeong et al., 2011); chromosomal analysis of those MSCs revealed multiple chromosomal aberrations (Jeong et al., 2011). In some instances, migrating MSCs were incorporated into the tumor microenvironment, where they exerted protumorigenic effects by reducing apoptosis and enhancing angiogenesis (Casiraghi et al., 2013). Importantly, allogenic MSCs (whether derived from different sources or administered through different routes for various clinical conditions) did not contribute to tumor progression during long-term follow-up (Casiraghi et al., 2013).
Moreover, MSCs have not undergone malignant transformation during clinical trials (Abdi et al., 2008; Trounson and McDonald, 2015). Notably, several reports highlighted the antitumor effects of MSCs, which involved inhibiting tumor cell proliferation and inducing tumor cell apoptosis (Kostadinova and Mourdjeva, 2020; Yang et al., 2019). With in the context of MSC-based clinical trials, there is an urgent need for protocols that enable large-scale manufacturing; moreover, stringent quality control is needed to avoid inconsistencies regarding the therapeutic agent. Therefore, standards and consensus assays should be established for MSC production and release processes (Robb et al., 2019). Other concerns to address are the establishment of markers to detect MSCs in recipients, as well as indicators for immune response monitoring in recipients.
Finally, the cost of MSC-based therapy is an important limitation for many patients. The above concerns must be addressed to ensure optimal short- and long-term therapeutic benefits.
In conclusion, despite considerable progress in MSC-based therapy, many aspects require careful consideration, such as MSC production, appropriate dosing and timing, frequency, optimal administration route, and transplantation protocol (Fig. 4). These challenges must be addressed in preclinical experiments before MSC-based therapy can be successfully translated into clinical applications.

Overview of the limitation of MSC-based clinical application.
Conclusion
DM is usually accompanied by progressive chronic complications. Conventional therapies include insulin injection and oral administration of antidiabetic drugs, both of which have limited clinical efficacy. MSC-based treatments provide an alternative strategy for controlling DM through immunomodulation and tissue regeneration. Hundreds of clinical trials regarding MSC-based therapy have been registered on clinicaltrials.gov; most are not yet completed. The available data are inconclusive; additional trials are needed for comprehensive statistical analysis. Furthermore, detailed investigations are needed regarding the mechanisms of MSC persistence, homing, and immunomodulation. Successful navigation of these challenges will guide future applications of MSC-based therapy.
Footnotes
Authors' Contributions
L.Z. and S.W. were responsible for conceptualization, methodology, data curation, and writing-original draft preparation; J.S.Q., Z.G.H., C.X.K., and N.N.H. were responsible for data curation and investigation; X.D.S. was responsible for supervision, writing-reviewing and editing, and funding acquisition.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This work was supported by the National Natural Science Foundation of China (Grant Nos. 82170865 and 81870593).