
Abstract
A combination of a MEK/ERK-signaling inhibitor and three chromatin-remodeling molecules enhances generation of platelet-producing megakaryocytes in vitro, possibly through direct reprogramming.
Direct lineage reprogramming (DLR) is a conversion of one specialized cell type to another without the need for a pluripotent intermediate. DLR can be achieved by combinations of transcription factors, cytokines, and small molecules modulating signaling pathways and/or epigenetic states, and involves reversal of metastable transcriptional states (Apostolou and Hochedlinger, 2013). The factors required for a particular DLR might give invaluable insight to the nature of the epigenetic constraints and master regulators determining lineage specification of the cell types used in the process. Factors driving a particular DLR from one somatic cell to another can in many cases also be used to enforce the same lineage specification from iPS cells, potentially allowing inexhaustible ex vivo production of a defined cell type.
A clinically relevant example is production of platelets (PLTs), the most abundantly produced cell type in our bodies with >1 × 1011 produced each day from megakaryocytes (MKs) shedding. The cytokine thrombopoietin (Tpo) is the primary regulator of platelet production, supporting the survival, proliferation, and differentiation of MKs (McCarty et al, 1995). Tpo binds to its surface receptor, c-Mpl, and acts through multiple downstream signaling pathways, including the PI-3 kinase-Akt, MAPK, and MEK-ERK1/ERK2 pathways. Erythropoietin (Epo) is also a thrombopoietic factor, especially in the absence of the Tpo (Hacein-Bey-Abina et al, 2020).
Direct reprogramming of fibroblasts to MKs has previously been accomplished using retroviral vector-mediated overexpression of defined transcription factors (Runx1, Gata2, Gata1, Lmo2, Tal1, and Myc) (Pulecio et al, 2016). Of these transcription factors Gata1 and Tal1 also support enforced MK development from iPS cells (Borst et al, 2017). Although there are potential advantages of generating MKs and PLTs platelets from iPS cells, such as a defined and scalable process using unlimited numbers of a self-renewing cell, current iPS-derived MKs are inferior compared with MKs derived from primary hematopoietic cells that are functionally similar to in vivo produced circulating platelets (Matsunaga et al, 2006). The field is, therefore, searching for novel factors enhancing MK/PLT ex vivo generation.
An intriguing study by Qin et al published in Cell Stem Cell describes a new method for generating PLT-producing MKs, not from an MK progenitor but from committed precursors of the developmentally related erythroid lineage. The state-of-the art culture method for producing functional PLTs from cord blood ex vivo involves culturing CD34+ cells to MK-progenitors with a yield of >1 × 1011 PLTs per cord blood unit (Matsunaga et al, 2006). Qin et al explore an alternative method where cord blood cells instead are cultured with Epo and other factors supporting generation of erythroid precursor cells, followed by an attempt to reprogram these cells to platelet-producing MKs using small molecule epigenetic and signaling modulators.
The screen for identifying chemical MK-reprogramming molecules was performed on CD235a+ erythroblasts (EBs) isolated from mononuclear umbilical cord blood cells cultured for 7 days in the medium supporting erythroid progenitor expansion. EBs were next recultured in medium where the erythroid factors were replaced by Tpo and combinations of candidate molecules. Importantly EBs generate 2%–3% phenotypic MKs at day 7, even without adding any of the small molecules. The article's main finding is that addition of the four molecules, histone methyltransferase inhibitor Bix01294 (B), DNA methyltransferase inhibitor RG108 (R), histone deacetylase inhibitor valproate (V), and PD0325901 (P), leads to a 10-fold increase in generation of MKs (Fig. 1).
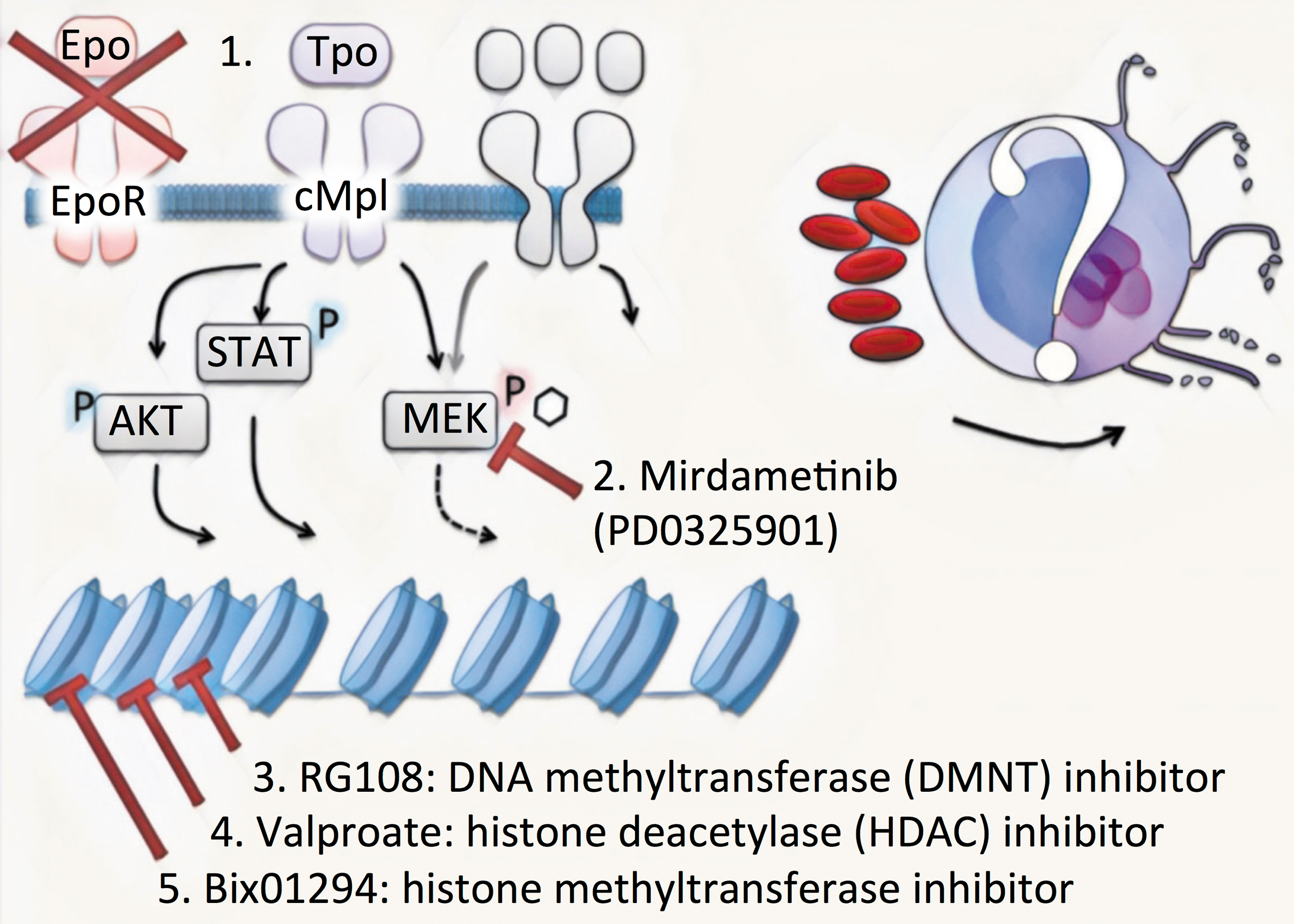
Efficient generation of PLT-producing MKs from EBs was accomplished by reculturing EBs in medium in which Epo was replaced by Tpo. Epo and Tpo signaling are highly related processes involving the same downstream mediators. The protocol uses 0.5 mM PD0325901 (P), a selective inhibitor of MEK and the downstream ERK pathway, affecting Epo, Tpo, and many other MEK-dependent cytokine signaling pathways. In addition, three epigenetic modifiers are used (BRV molecules). The MK-inducing medium contains 0.5 mM BIX-01294 (B), a highly selective histone methyltransferase inhibitor. It should be noted that a near 100% inhibition of methyl transferase activity is achieved already with 10 μM Bix01294 (Chang et al, 2009) suggesting lower doses may be sufficient. In addition, the medium contains 0.04 mM RG108 (R), a DNA methyltransferase inhibitor and 0.2 mM valproate (V), an HDAC inhibitor. Epo, erythropoietin; HDAC, histone deacetylase; MK, megakaryocyte; PLT, platelet; Tpo, thrombopoietin.
Of the four BRVP molecules only P is able to increase MK development. The article further demonstrates the MKs generated from EBs in presence of the BRVP molecules produce a large number of fully functional PLTs. The PLT yield per unit of cord blood is, however, lower than for state-of-the art methods (Matsunaga et al, 2006). To investigate the dynamic cellular changes in the culture single-cell RNA-Seq and ATAC-Seq analyses of initial EBs and of cells collected at several time points during the culture toward MKs were performed. The bioinformatic trajectory analyses show a trajectory involving early expansion of cells with an erythroid/MK bi-potent expression signature before the MKs emerge. The findings are potentially transformative in the field but raise two important questions.
First, is the observed increase in MK generation a result from true reprogramming? An important concern challenging this claim is the observation the EB population is able to generate MKs also without the BRVP factors, suggesting the EB population is contaminated by a small number of cells with MK potential. Although addition of the BRVP factors indeed increases MK generation, it does not demonstrate direct lineage conversion. Second, what is the exact mechanistic role of Tpo and the BRVP molecules? Since none of the scRNA-Seq and ATAC-Seq experiments were performed without the BRVP molecules as a negative control, we can only speculate about the mechanistic roles of BRVP factors.
In particular, the modulation of the MEK/ERK pathway using PD0325901 seems to be a new tool interesting for disturbing normal erythroid/MK development. Although both Epo and Tpo signaling involve the MEK/ERK pathway, the findings suggest erythroid maturation is more dependent on this pathway than bi-potent MK/erythroid progenitor cells. Interestingly, PD0325901 induces spontaneous erythroid differentiation of hematopoietic stem cells to erythroid progenitors in vitro a culture of human CD34+ cells containing Tpo, SCF, and Flt-3L (Zarrabi et al, 2018). Perhaps it is taking the speculations too far, but could PD0325901 with or without the three epigenetic modifying chemicals be the sought-after factor needed for maintaining MK/erythroid progenitors in culture? If so the field is an important step closer to large-scale production of both PLTs and red blood cells for transfusion.
To summarize, I believe the readers of this Journal will anticipate the experimental evidence supporting the DLR claims made by Qin et al. If they hold true, the BRVP factors may prove to be a signaling epigenetic power combo to be evaluated in the readership's other lineage reprogramming efforts.
Author Contribution
J.F. wrote the article and generated the figure.
Footnotes
Author Disclosure Statement
The author declares there are no conflicting financial interests.
Funding Information
No funding was received for this article.