
Abstract
As a gene with antiaging functions, sirtuin6 (SIRT6) belonging to the sirtuin family plays a vital role in DNA repair, telomerase function, and cellular senescence, as well as maintains epigenomic stability and promotes longevity. However, its role in cell senescence in large animals, such as buffaloes, remains unknown. Fibroblasts are commonly used for somatic reprogramming, and their physiological characteristics affect the efficiency of this process. We aimed to elucidate the role of SIRT6 in cellular senescence and proliferation and analyze its effect on the biological function of buffalo fibroblasts to help improve the efficiency of buffalo somatic cell reprogramming. The expression of SIRT6 and related DNA damage was measured in buffalo fibroblasts obtained at different developmental stages (in the fetus and at 3 and 10 years of age), and the effect of SIRT6 knockdown on the senescence of buffalo fetal fibroblast was investigated. An inverse relationship was observed between SIRT6 expression and senescence in buffalo fibroblasts obtained from animals of various ages. This was accompanied by decreased cell growth, viability, and increased DNA damage. Short hairpin RNA-mediated SIRT6 knockdown accelerated the senescence of buffalo fetal fibroblasts. It blocked the cell cycle during in vitro cell culture, which further enhanced DNA damage, particularly with respect to the telomeres. Collectively, our findings suggest that SIRT6 expression was closely associated with buffalo senescence in fibroblasts. These findings serve as a foundation to better understand the cellular functions of SIRT6 and also aid in selecting donor cells for buffalo somatic cell reprogramming.
Introduction
Sirtuin6
Consequently, the sirtuin protein family has been referred to as antiaging and longevity proteins (Sinclair and Guarente, 1997). Sirtuins inhibit aging in humans through many signaling pathways, including those associated with apoptosis, mammalian target of rapamycin, AMP-activated protein kinase, and phosphatidylinositol 3 kinase (PI3K) (Wang et al., 2020).
As a vital member of the sirtuin family associated with antiaging activities, SIRT6 plays a crucial role in DNA repair, telomerase function, cellular senescence, maintenance of epigenomic stability, and promoting longevity (Liao and Kennedy, 2016). SIRT6 is localized in the nucleus, and functions as an adenosine diphosphate-ribosyltransferase (ADP-) and nicotinamide adenine dinucleotide+-dependent (NAD+-) deacylase of acetyl and long-chain fatty acyl groups, primarily in the deacetylation of the 9th lysine (H3K9) on H3 histone, the 56th lysine (H3K56), and the C-terminal binding protein-interacting protein on H3 histone (CtIP) (Wang et al., 2020). To date, few studies have been conducted on SIRT6 in large animals. SIRT6 overexpression decreases nuclear damage by dampening cytotoxicity and attenuating aging in porcine fetal fibroblasts (Xie et al., 2012). Furthermore, demethylation significantly enhances the binding of nuclear respiratory factor 1 (NRF-1) and E2F transcription factor 1 (E2F1_ to the SIRT6 core promoter in bovine adipocytes (Hong et al., 2018).
Furthermore, SIRT6 is hypothesized to directly contribute to life span extension, and enhanced SIRT6 activity reportedly increases the life span of some rodent species (Tian et al., 2019).
Owing to their convenient availability and ease of culture, fibroblasts are the main donor cells used for somatic reprogramming, and somatic cell nuclear transfer (SCNT) and induced pluripotent stem cells (iPSCs) are commonly used for their culture (Poehland et al., 2007). Fibroblasts with different degrees of senescence can affect the reprogramming results of these methods. For instance, the fusibility and in vitro developmental potential of embryos reconstructed from long-term cultured fetal fibroblasts were significantly lower than those reconstructed from short-term cultures in goats (Lan et al., 2006). Similar results were obtained for blastocyst rates, which were higher for fibroblasts at passages 10 and 20 than for cells at passage 30 in male goats (Liu et al., 2016).
These results demonstrate that the senescence of donor cells, such as fibroblasts, can influence the in vitro development of nuclear transfer embryos. Cellular senescence also acts as a barrier to classical Yamanaka factor reprogramming (Banito et al., 2009). Sharma et al. reported that SIRT6 substantially improved reprogramming efficiency in older human dermal fibroblasts (Sharma et al., 2013).
These findings suggest that the senescence of donor cells influences reprogramming efficiency. However, the factors affecting cellular senescence and their regulatory mechanisms require further investigation. We previously used fibroblasts as donor cells to produce cloned buffalo (Shi et al., 2007) and iPSCs (Deng et al., 2012). However, reprogramming is inefficient owing to donor fibroblasts derived from different individuals (Luo et al., 2022).
To overcome this limitation and in keeping with the role of SIRT6 in aging, we aimed to determine the cell senescence, proliferation ability, and the expression of SIRT6 in fibroblasts obtained from buffaloes at different developmental ages (the fetus, at 3 and 10 years of age). SIRT6 was knocked down to investigate its effect on senescence, and the characteristics of buffalo fibroblasts were investigated. We believe our findings would provide novel insights into the physiological role of SIRT6 in buffalo fibroblasts, as well as directions to improve the efficiency of buffalo-cloning protocols.
Materials and Methods
Harvest and culture of buffalo fibroblasts
Ear skin biopsies were obtained from three male buffaloes at various developmental ages (the fetus, at 3 and 10 years of age). The tissues were disinfected with 75% alcohol and phosphate-buffered saline (PBS) containing 10% of penicillin and streptomycin (Gibco), and the PBS containing 3% of penicillin and streptomycin successively for 30 seconds. They were washed thrice with PBS containing 1% of penicillin and streptomycin before the tissues were cut into small pieces. Samples (1 mm3) were resuspended and inoculated in 60-mm culture dishes with Dulbecco's modified Eagle's medium (DMEM) containing 10% of fetal bovine serum (FBS; Gibco). The cell culture conditions used in this study were as follows: CO2 incubator (Thermo Fisher Scientific) at 37°C, 100% relative humidity, and 5% CO2. After 12 hours of culture, the liquid was changed every 24 hours to prevent contamination. When the primary cells reached 70%–80% confluence, the tissue blocks were removed, and the fibroblast outgrowth was trypsinized and either passaged at a ratio of 1:3 or cryopreserved. Changes in cell morphology and proliferation were observed daily using an inverted phase-contrast microscope, and the images were recorded. All experiments regarding animals were performed in the Guangxi Key Laboratory of Animal Breeding and Diseases Control and were conducted in accordance with its guidelines for the care and use of laboratory animals.
SIRT6 knockdown in buffalo fetal fibroblasts
Short hairpin RNA (shRNA) was used to knock down SIRT6 expression in the buffalo fetal fibroblasts. The sh-SIRT6 lentiviral vector was constructed by cloning shRNA of SIRT6 into the Psi-LVRU6GP vector. Lentiviruses were produced in 293 T cells by cotransfecting 3 μg NRF, 2 μg vesicular stomatitis virus glycoprotein (VSVG), and 5 μg sh-SIRT6 lentiviral vector using a transfection reagent (9 μL; Roche, Switzerland). The transfection mixture was incubated at room temperature for 15 minutes and added dropwise to the Petri dish. The transfection media were replaced 12 hours after transfection. The viral supernatants were harvested at 60 hours with 0.45-μm membrane filtration and centrifuged at 40,000 rpm at 4°C for 2 hours.
When buffalo fetal fibroblast cells reached ∼60%–70% confluency in a 60-mm-diameter Petri dish, the medium was removed and replaced with an infection medium, including 1 mL of viral supernatants, 2 mL of culture medium (DMEM with 10% FBS), and 6 μg/mL of polybrene (Deng et al., 2012). After 48 hours of lentivirus infection, the green fluorescent protein (GFP) fluorescence was determined using a fluorescence microscope (Olympus, Japan). The SIRT6 knockdown efficiency was evaluated using quantitative real-time polymerase chain reaction (qRT-PCR) to determine its relative expression level.
Quantitative real-time polymerase chain reaction
The relative expression levels of SIRT6, p16, p21, and p53 in cultured cells were quantified using qRT-PCR, and the primers used are listed in Table 1. Total RNA was obtained from the samples using the TRIzol reagent (Invitrogen) and used to synthesize cDNA for reverse transcription. According to the manufacturer's instructions, the qRT-PCR procedure consisted of three stages: a holding stage, 95°C, 5 minutes; 40 cycles of cycling stage, 95°C, 15 seconds, 60°C, 30 seconds, 72°C, 30 seconds, and the fluorescence was collected; a melting curve stage, 95°C, 15 seconds, 60°C, 60 seconds, 95°C, 30 seconds, 60°C, 15 seconds. Each measurement was performed in triplicate. The 2−ΔΔCt method was used to evaluate the relative expression levels of the target genes. β-actin was chosen as the reference gene for assessing mRNA expression.
The Primers Used for the Quantitative Real-Time Polymerase Chain Reaction
Cell growth curve
The cells were inoculated into 24-well plates at a density of 5000 cells/mL and cultured in 5% CO2 and 95% air. The cell culture medium was changed every 2 days. At the same time every day, three holes were punctured, and the cells were counted using a living cell counting board (MC, China). The number of cells within each well was counted thrice, and the average values were recorded as the number of new cells obtained in each 24-hour period.
Cell cycle assay
Cells were inoculated into 35-mm Petri dishes, and the cell cycle steps were assessed when the cell confluence reached 90%. The cell cycle was monitored using the LeaGene reagent according to the manufacturer's instructions (LeaGene, Beijing, China). The lowest sample loading speed was selected, and flow cytometry was used to determine. The results were analyzed using ModiFit 4.0 software.
Cell apoptosis assay
The cells were trypsinized (Gibco) and were collected by centrifugation at 1, 200 rpm for 3 minutes. The cells were washed with the precooled PBS and, after further centrifugation, resuspended in 500 μL 1 × binding buffer. They were then incubated with 5 μL of Annexin V-APC (Leagene) and propidium iodide (Leagene) for 15 minutes in the dark and assessed using flow cytometry.
Measurement of β-galactosidase
The β-galactosidase activity was evaluated using a Senescence Detection Kit (Beyotime, China) according to the manufacturer's instructions. The percentage of positive cells was determined by counting the number of blue-stained cells versus the total number of cells in the same field (n = 4/group).
Immunofluorescence assay
The cells were plated on sterilized glass coverslips and placed on a 24-well plate. The cells were fixed in 4% formaldehyde for 10–15 minutes at room temperature. Permeabilization was performed using 1% Triton X-100 for 5 minutes at room temperature, followed by incubation with 1% bovine serum albumin in PBS for 45 minutes. The cells were then incubated overnight at 4°C with the indicated primary antibodies (Abcam, UK), which were prepared in the blocking buffer at dilutions ranging from 1:100 to 1:300. The cells were then incubated with species-specific Alexa Fluor-conjugated secondary antibodies (Abcam) for 1 hour at room temperature. Nuclei were stained with 10 μg/mL 4′,6-diamidino-2-phenylindole and incubated for 15 minutes at room temperature. The cells were washed twice with 1 × PBS between each step. Images were acquired using confocal microscopy (Leica Microsystems).
Western blot analysis
Total proteins from buffalo fetal fibroblasts (BFFs) were homogenized using the radioimmunoprecipitation assay (RIPA) buffer (Servicebio, G2002, Wuhan, China), contained 1% protease and phosphatase inhibitor, and incubated 30 minutes on ice. The cell lysate was obtained and centrifuged at 12,000 rpm at 4°C for 10 minutes. Protein concentrations were determined using the BCA Protein Assay Kit (Solarbio, PC0020, Wuhan, China). Proteins were separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis, transferred to a polyvinylidene fluoride membrane (Millipore, IEVH00005, Burlington, MA), and then incubated with antibodies (SIRT6, Abcam, ab191385; TRF2, Abcam, ab108997; GAPDH, Proteintech, 60004-1-Ig, Wuhan, China) overnight at 4°C and then with an HRP-conjugated secondary antibody for 1 hour at room temperature. Pictures were captured by an imaging system (UVP; United States).
Statistical analyses
Data were analyzed using the chi-square test. GraphPad InStat 3 software was used for the statistical analyses. Statistical significance was set at p < 0.05.
Results
Cell growth curves of fibroblasts derived from buffaloes at different developmental stages
Expanded fibroblasts grew around the fetal buffalo tissues within 24 hours of inoculation. In contrast, no growth was observed in 3- and 10-year-old buffalo tissues until 2–3 days (Fig. 1a). After the early passage, fibroblast morphology did not significantly differ among the three age groups (Fig. 1a). However, after passage 8, the proliferative ability of the 10-year-old buffalo fibroblasts was slower than that of the other two groups, and they appeared to cease proliferating. Conversely, fetal and 3-year-old buffalo-derived fibroblasts maintained their spindle cell morphology and continued to proliferate even after the tenth generation. Growth curves of the third generation of fibroblasts at different ages indicated that buffalo fetal fibroblasts had a higher proliferation rate and viability than those from 3- to 10-year-old buffalo fibroblasts (Fig. 1b).
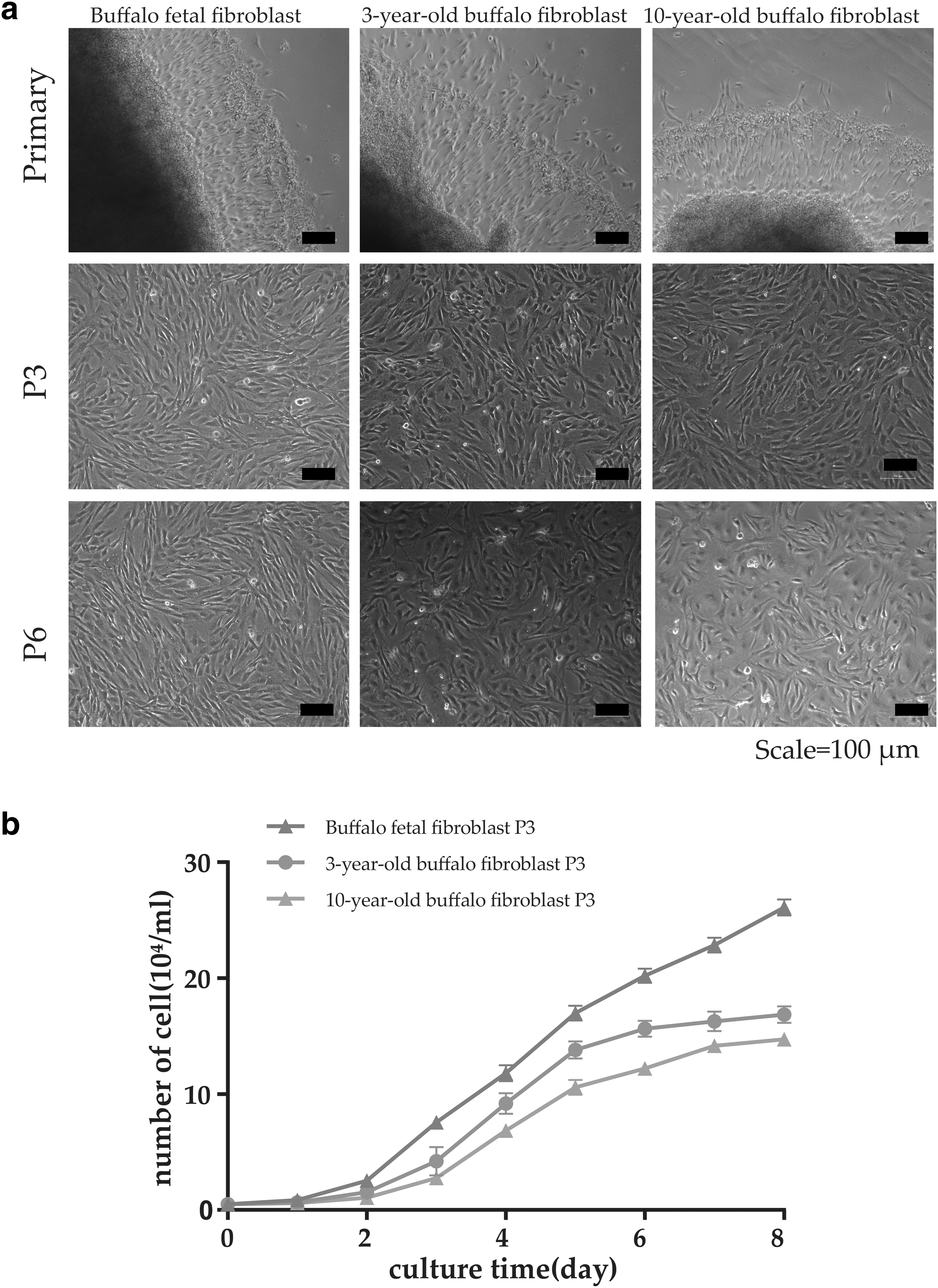
Cell morphology and growth curve of buffalo fibroblasts obtained from buffaloes at different ages.
Correlation between SIRT6 expression and buffalo fibroblast senescence at different ages
The β-galactosidase activity, which can be used as a biomarker to identify senescent cells (Dimri et al., 1995), was measured to analyze the correlation between SIRT6 expression and buffalo fibroblast senescence at different ages. We observed that the number and intensity of β-galactosidase-positive cells in 3- and 10-year-old buffalo fibroblasts were significantly higher than those in buffalo fetal fibroblasts (Fig. 2a). The β-galactosidase-positive cell rates of fetal and 3- and 10-year-old buffalo fibroblasts were 1.53 ± 0.96, 14.55 ± 4.35, and 52.92 ± 9.61, respectively (Fig. 2b). The proportion of β-galactosidase-positive cells increased with age.

Senescence features of buffalo fibroblasts were influenced by the age of the buffalo.
The relative expression levels of p16, p21, and p53, which are senescence-associated marker genes, were also measured using qRT-PCR during passage 3 of fibroblasts obtained from buffaloes of different ages. The results indicated that SIRT6 expression in buffalo fetal fibroblasts was significantly higher than that in the other two groups (Fig. 2c). The expression of the p16 gene increased significantly with age (p < 0.05). The expression of p21 and p53 genes did not significantly differ between fetal and 3-year-old buffalo fibroblasts (p > 0.05); however, the expression of p21 and p53 increased significantly in 10-year-old buffalo fibroblasts (p < 0.05) (Fig. 2d). These results suggest that SIRT6 expression is negatively correlated with the age of the buffalo.
DNA damage of buffalo fibroblasts at different ages
γ-H2AX is a marker of DNA damage (DNA double-strand breaks: DSB) (Nagai et al., 2015), and each γ-H2AX focus corresponds to one DSB. Transferrin receptor 2 (TRF2) is used as a marker of telomere localization, and the overlap of γ-H2AX and TRF2 represents the site of telomere damage (Necasova et al., 2017). Immunofluorescence observations revealed that the focus of γ-H2AX in 10-year-old buffalo fibroblasts was significantly higher than that in the other two groups. Furthermore, the focus area was larger, with more overlapping sites of γ-H2AX and TRF2 (Fig. 3). These results indicated that the number of DNA damage sites in aged buffalo fibroblasts was higher than that in buffalo fetal fibroblasts and that DNA damage was localized to the telomeres.

DNA and telomere damage sites as determined by immunostaining. TRF2 antibody, γ-H2AX antibody, and nuclei visualized with DAPI staining in buffalo fibroblasts. The merged figures are shown in the panels. Scale bars indicate 5 μm. DAPI, 4′,6-diamidino-2-phenylindole.
SIRT6 knockdown affects the proliferation and apoptosis of buffalo fetal fibroblasts
To investigate the effect of SIRT6 on the proliferation and apoptosis of buffalo fibroblasts, the cell growth curve, cell cycle, and apoptosis of passage 3 buffalo fetal fibroblasts were assessed after SIRT6 knockdown. Buffalo fetal fibroblasts were transfected with shRNA-SIRT6 lentivirus (shRNA-SIRT6 group) or blank vector lentivirus (negative control [NC] group). After 12 and 48 hours of lentivirus treatment, GFP fluorescence was observed in buffalo fetal fibroblasts (Fig. 4a), indicating a successful cell transfection. qRT-PCR results revealed that the SIRT6 mRNA expression was reduced by 93.76% following SIRT6 knockdown (Fig. 4b). The protein expression level of SIRT6 was also decreased significantly after SIRT6 knockdown (Fig. 4f). The cell growth curve of the NC group exhibited a typical “S” shape. However, the number of buffalo fetal fibroblasts in the shRNA-SIRT6 group increased on days 1–6 and decreased thereafter (Fig. 4c).

Knockdown of SIRT6 affects the proliferation and apoptosis of buffalo fetal fibroblasts.
Flow cytometry analysis revealed that the proportion of S- and G2-phase cells was 32.6% and 7.28%, respectively, and the cell proliferation index was 39.88% in the NC group. Correspondingly, the proportions of S-phase and G2-phase cells in the shRNA-SIRT6 group were 14.74% and 1.51%, respectively. The cell proliferation index was 16.25% (Fig. 4d). The number of S- and G2-phase cells and the cell proliferation index in the shRNA-SIRT6 group were significantly lower than those in the NC group (p < 0.05). The results of apoptosis detection indicated that the proportion of early and late apoptotic cells in the shRNA-SIRT6 group was 53.66% and 1.27%, respectively, which was significantly higher than that of the NC group (1.12% and 0.34%, respectively, p < 0.05) (Fig. 4e). These results indicated that SIRT6 knockdown decreased the proliferative ability and increased the apoptosis of buffalo fetal fibroblasts.
SIRT6 knockdown affects senescence and DNA damage of buffalo fetal fibroblasts
The β-galactosidase staining results revealed that most cells in the shRNA-SIRT6 group were stained dark blue (Fig. 5a), and the number of β-Gal-positive cells was significantly higher than that in the NC group (p < 0.01) (Fig. 5b). qRT-PCR indicated that the relative expression levels of p16, p21, and p53 in the shRNA-SIRT6 group were significantly higher than those in the NC group (p < 0.05) (Fig. 5c).
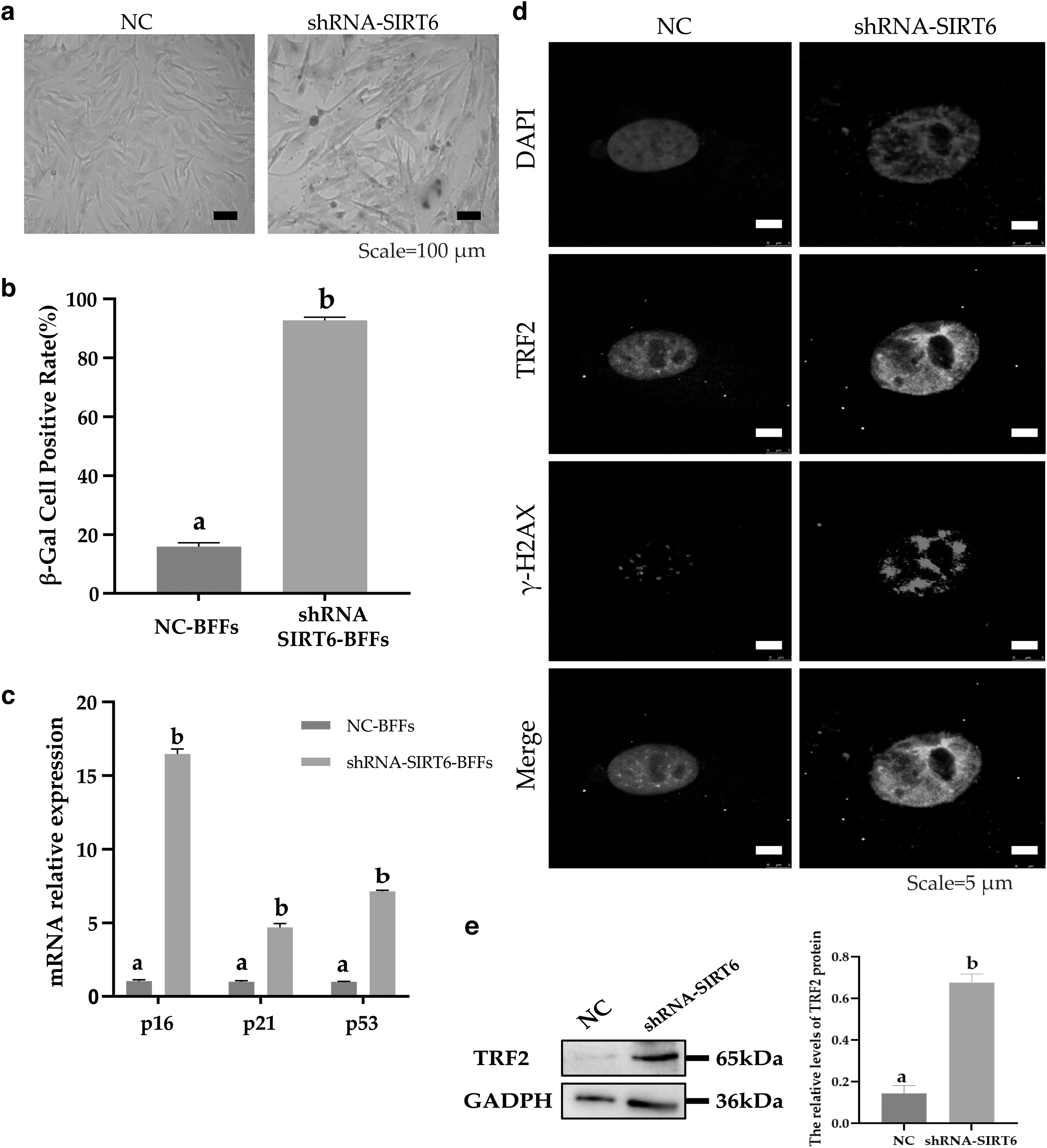
Knockdown of SIRT6 influences the senescence features of buffalo fibroblasts.
Assessment of the DNA and telomere damage sites in buffalo fetal fibroblasts after SIRT6 knockdown revealed that decreased SIRT6 expression increased the number of γ-H2AX-specific binding sites (Fig. 5d), suggesting that the DNA damage sites and area of buffalo fetal fibroblasts were increased. In addition, the protein level of TRF2 increased significantly after SIRT6 knockdown (Fig. 5e). The results also suggested that the overlap between γ-H2AX and TRF2 was increased. Hence, SIRT6 knockdown enhances DNA damage, especially in telomeres. These results also suggest that SIRT6 knockdown accelerated the senescence of buffalo fetal fibroblasts and enhanced DNA damage.
Discussion
Skin fibroblasts are the main donor cells used for SCNT and iPSC technologies owing to their availability and ease of culture (Poehland et al., 2007). However, the source of donor cells for in vitro culture can affect the cloning efficiency. Adult and fetal somatic cells can serve as donor cells for transgenic SCNT embryo production (Cho et al., 2004). Furthermore, aging adult fibroblasts can also support nuclear transfer and transcription factor-mediated reprogramming when used in pigs (Li et al., 2018). However, donor cell senescence has been postulated to affect reprogramming efficiency. In the production of cloned goats, the fusibility and in vitro developmental potential of embryos reconstructed from long-term cultured cells (passages 20–25) were significantly lower than those reconstructed from short-term cultured cells (passages 3–5) (Lan et al., 2006).
Similarly, the blastocyst rate of reconstructed embryos when using passage 30 fibroblasts from male goats was lower than that of passages 10 and 20 (Liu et al., 2016). The results from reconstructed embryos obtained from donor cells at different passages indicated that the replicative senescence of donor cells affects the efficiency of embryo production in goats. Notably, Bressan et al. observed no differences in fusion, cleavage, eight-cell embryo, or blastocyst rates between groups reconstructed with early- or late-passage donor fibroblasts in cattle (Bressan et al., 2013). However, it may have been difficult to observe significant differences between the different donor cell types in these animals owing to the relatively low efficiency of bovine cloning (Akagi et al., 2014).
Based on the pivotal regulatory role of SIRT6 in prolonging the cellular life span, Sharma et al. demonstrated that adding this gene during reprogramming substantially improved the efficiency of human dermal fibroblasts when cells from older human subjects were used (Sharma et al., 2013). However, we lack sufficient information regarding the role of SIRT6 in large animals. The effects on cattle have been reported to be related only to carcass traits (Gui et al., 2018; Gui et al., 2015; Hong et al., 2018). In this study, we focused on the role of SIRT6 in the senescence of buffalo fibroblasts successfully harvested and cultured from the fetus and from 3- to 10-year-old buffalo ear skin tissues.
Comprehensive analyses of the biological characteristics of fibroblasts from animals of different ages revealed a significant decrease in S-phase cells and the cell proliferation index with age. The expression of the senescent marker genes, p16, p21, and p53, was also increased significantly in aged buffalo fibroblasts. We demonstrated that aged buffalo fibroblasts had a much lower proliferation capacity in vitro than that in young buffalo fibroblasts. This conclusion is consistent with the findings of previous studies in humans (Zhai et al., 2016). Furthermore, we observed more β-galactosidase-positive cells in buffalo fibroblasts obtained from 10-year-old animals compared with those from the fetus. Similar results have been observed in goat skin fibroblasts (Liu et al., 2016).
SIRT6 is a critical gene that regulates longevity and has been implicated in progeria. In this study, we observed that SIRT6 expression decreased significantly with age in buffalo-derived fibroblasts, suggesting that this gene is inversely related to the degree of cellular senescence in these cells. SIRT6 knockdown in fetal fibroblasts resulted in coarse, flat, and weakly refractive cells, indicating senescence. Several cells died during the culture process, and the growth curves no longer exhibited the typical S-type. The number of S- and G2-phase cells and the cell proliferation index were significantly lower than those of nontransfected buffalo fetal fibroblasts. Moreover, p16, p21, and p53 mRNA expression levels and the number of β-galactosidase-positive cells were significantly increased.
In addition to donor cell senescence, the cell cycle of donor cells affects the development of SCNT-derived embryos in vitro. Currently, successful SCNT-derived embryos use either G0- or G1-phase donor cells as the starting materials (Akagi et al., 2014). Furthermore, one study suggested that the in vivo developmental ability of SCNT-derived embryos tended to be higher for G1-phase cells than that for G0-phase cells because the homogeneous expression derived from the former cell types likely contributed to their higher success rates (Iwamoto et al., 2012).
In this study, we observed that SIRT6 knockdown in buffalo fibroblasts affected the cell cycle, such that the number of S- and G2-phase cells in the shRNA-SIRT6 group was significantly lower than that in the control group. SIRT6 may improve the reprogramming efficiency by modulating the cell cycle of donor cells because the capacity to reprogram the somatic cell nucleus of oocytes may largely depend on the cell cycle stage of the donor cells within a few hours of activation (Akagi et al., 2014). However, this hypothesis requires further investigation.
Furthermore, we observed that p53 expression increased after SIRT6 knockdown, and inhibition of this gene can improve the efficiency of nuclear transplantation in cattle (Ma et al., 2015). In porcine fetal fibroblasts, SIRT6 overexpression decreases nuclear damage (Xie et al., 2012). This is consistent with our findings in buffalo fibroblasts after SIRT6 knockdown, where DNA damage was increased, as suggested by the increased damage to telomeres. This implies that SIRT6 prevents premature senescence in buffalo fibroblasts by controlling DNA repair and maintaining normal telomere function.
Footnotes
Acknowledgment
The authors wish to thank Dr. Dev Sooranna of Imperial College in London for editing the article.
Authors' Contributions
J.L. was involved in the formal analysis, data curation, and writing—original draft preparation. J.C. was involved in the methodology, data curation, and writing—original draft preparation. J.C. was involved in the methodology and data curation. Y.P. was involved in software. R.Z. was involved in validation. S.Y. was involved in conceptualization, writing—review and editing, supervision, project administration, and funding acquisition. L.Z. was involved in conceptualization, formal analysis, writing—review and editing, supervision, and project administration.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This work was supported by the Natural Science Foundation of Guangxi Province (grant nos. 2019GXNSFDA185002 and 2022GXNSFDA035064) and the National Natural Science Foundation of China (grant no. 31860644).