
Abstract
Oocytes contain reprogramming machinery that can transform somatic cells into totipotent cells. In this study, we aimed to isolate and characterize nanovesicles from mature porcine oocytes and described them for the first time as “intra-ooplasmic vesicles (IOVs)”. Isolated IOVs had an average diameter of 186.3 ± 10.8 nm. Proteomic analysis revealed 467 peptide reads, with the top 20 proteins related to reprogramming, antioxidative defense, cytoskeleton, heat shock proteins, and metabolism. Protein–protein interaction and gene ontology analysis indicated that these proteins were involved in various biological pathways, including protein folding, metabolism, and cellular responses to stress. Supplementing cultured fibroblasts with IOVs resulted in the expression of the pluripotency marker OCT4 and the early trophoblastic marker CDX2 and increased expression of the corresponding mRNAs together with increasing KLF4 and SALL4 expression. IOV treatment of fibroblasts for 14 consecutive days resulted in changes in cell morphology, with increased expression of ZEB2 and YBX3 as markers for epithelial-to-mesenchymal transition (EMT). These results provide a rationale for further characterization of IOVs, investigation of potential reprogramming capabilities for EMT, and the generation of induced pluripotent or oligopotent stem cells.
Introduction
Oocytes possess unique capabilities as they contain specific reprogramming machinery, comprising proteins, RNAs, lipids, and small molecules with activities of transcription, epigenetic modification, signaling, and metabolism, that enables the reprogramming of transplanted somatic nuclei to a totipotent or pluripotent state (Pfeiffer et al., 2011; Rossant, 2009). Reprogramming induces the expression of certain genes and suppresses that of others to convert differentiated cells into pluripotent stem cells or other cell types (Cherry and Daley, 2012; Goya et al., 2018).
Oocytes have the remarkable ability to reprogram the genetic material of transplanted somatic nuclei in various ways (Saadeldin, 2015). First, the enucleated oocyte can reprogram somatic nuclei upon being transferred into it in a process called somatic cell nuclear transfer or cloning (Wilmut et al., 1997). Second, they can influence somatic cell nuclei to avoid obligatory cell cycle checkpoints and undergo premature condensation (Szollosi et al., 1986). Third, they can induce somatic cells to enter meiosis through reduced division, as observed in the process of haploidization (Kubelka and Moor, 2008; Saadeldin et al., 2016; Tesarik, 2002). The meiotic machinery of the ooplasm can be used to generate artificial gametes by injecting male or female nuclei into enucleated mature or immature oocytes (Tesarik et al., 2021; Tesarik et al., 2001; Trounson, 2001; Tsai, 2000).
Previous reports have revealed the significant contribution of oocyte extracts to nuclear reprogramming (Byrne et al., 2003; El-Gammal et al., 2020; El-Gammal et al., 2018; Ganier et al., 2011; Hansis et al., 2004); however, these methods of retrieving the oocyte extract, such as crushing oocytes with ultrasonic waves (El-Gammal et al., 2020) or ultracentrifugation (Hansis et al., 2004) can yield both nuclear and ooplasmic components.
In this study, we proposed the term “intra-ooplasmic vesicles (IOVs)” to illustrate the nanovesicles inside the ooplasm and provided ultramicroscopic images and contents of cargo proteins. Furthermore, we aimed to examine the potential reprogramming effects of IOVs on the cultured adult somatic cells.
Materials and Methods
Chemicals and reagents
Unless otherwise specified, all chemicals and reagents were purchased from Sigma-Aldrich (St. Louis, MO).
Oocyte retrieval and in vitro maturation
Experiments were conducted in accordance with the guidelines for the management of laboratory animals at Chungnam National University (Approval No. 202103-CNU-079). Porcine ovaries were obtained from a slaughterhouse and transported to the laboratory in a physiological saline solution (NaCl 0.9%) at 34°C–36°C within 4 hours (Bang et al., 2023). Cumulus–oocyte complexes (COCs) were retrieved through aspiration, and oocytes surrounded by compact cumulus cell layers were selected and washed three times with HEPES buffered Tyrode's medium containing 0.05% polyvinyl alcohol. The COCs were matured through incubation for 22 hours in four-well dishes (Nunc; Thermo Fisher Scientific, Roskilde, Denmark) with maturation medium comprising TCM-199 (Gibco, Waltham, MA), porcine follicular fluid (10%), L-cysteine (0.57 mM), sodium pyruvate (0.91 mM), epidermal growth factor (10 ng/mL), kanamycin (75 μg/mL), insulin (1 μg/mL), and 10 IU/mL of both human and equine chorionic gonadotrophin. Subsequently, the COCs were cultured under the same conditions without hormones for 22 hours (Fang et al., 2023; Fang et al., 2021).
Mature COCs were harvested, and cumulus cells were removed with hyaluronidase, washed twice, and placed in microdrops of PZM-5 medium (IFP Co. Ltd., Yamagata, Japan) covered with mineral oil. Finally, they were incubated at 38.5°C in a humidified atmosphere with 5% CO2.
Isolation and characterization of IOVs
Cumulus-free oocytes (n = 1200) were placed in 10 μL microdrops (10 oocytes per microdrop) of protein-free PZM-5 medium. The microdrops were overlaid with mineral oil in the micromanipulation dishes. Oocytes were held by a holding micropipette and ooplasms were aspirated through a 16 μm needle using a micromanipulator (Narishige, Japan) with avoiding aspiration of the first polar body and the oocyte nuclei. The aspirated ooplasm was then released into the PZM-5 medium (Fig. 1A). PZM-5 medium containing the ooplasms was aspirated, pooled, diluted once with phosphate-buffered saline (PBS), mixed by gentle pipetting, and centrifuged at 16,000 g for 30 minutes. Clear supernatants were used to isolate IOVs through targeted protein binding and nanofiltration using the PureExo Exosome Isolation Kits (101 Bio, Palo Alto, CA), according to the manufacturer's instructions (Saadeldin et al., 2022). IOV pellets were eluted in 25 μL of PBS solution and IOVs were characterized using nanoparticle tracking analysis (NTA, NanoSight NS300; Malvern Instruments, Worcestershire, United Kingdom) (Mehdiani et al., 2015).

Isolation and characterization of IOVs.
Moreover, IOVs were imaged using an OMEGA-energy filtering transmission electron microscope (ZEISS LEO 912; Carl Zeiss, Jena, Germany) after negative staining with 2% uranyl acetate (Lee et al., 2021; Saadeldin et al., 2014).
Preparation of protein fraction of mature oocytes and IOVs and proteomic analysis
Protein isolation and proteomics were conducted according to previously reported methods (Lee et al., 2021) and our recent report (Saadeldin et al., 2022). In vitro matured oocytes and the eluted IOV protein contents were estimated using the bicinchoninic acid method. Protein fractionation was performed through sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Gel was treated with ammonium bicarbonate and acetonitrile for Coomassie Brilliant Blue staining (Lee et al., 2020), rinsed twice with distilled water and acetonitrile (100%), and dried and treated with ammonium bicarbonate and dithiothreitol at 56°C. Following these steps, the gel was further treated with 100 nM iodoacetamide to reduce alkylate S–S bridges and then trypsinized. The resulting peptides were purified and concentrated using MGU30–C18 trapping columns (LC Package). The proteins were subjected to MS and MS/MS using an LTQ-Velos ESI ion trap mass spectrometer (Thermo Scientific, Waltham, MA). The data-dependent mode was used for the fragmentation of the most abundant peaks from the full MS scan, with 27% normalized collision energy. The eluted peptides were analyzed using a mass spectrometer at an electrospray voltage of 2.3 kV.
The maximum ion injection times were 100 ms each for the MS and MS/MS scans. The automatic gain control target settings were 5.0 × 104 and 1.0 × 105 for the MS and MS/MS scan modes, respectively. The dynamic exclusion duration was set to 180 seconds and the exclusion mass width was 0.5 Da. The mass range for acquiring the MS spectra was 150–2000 m/z. Protein identification was performed using MASCOT 2.4 with a false discovery rate of 1% as the cutoff value. The relative quantification of proteins was performed using the exponentially modified protein abundance index and expressed as protein contents in molar (mol %) (Ishihama et al., 2005).
Bioinformatics
The Database for Annotation Visualization and Integrated Database (DAVID) (https://david.ncifcrf.gov/tools.jsp) was used for functional analysis of the complete set of genes. UniProt accession IDs were used as inputs for the species name Sus scrofa. Gene ontology (GO), including biological process (BP), cellular component (CC), molecular function (MF), and the canonical pathway involved was identified for the group of genes using Kyoto Encyclopedia of Genes and Genomes (KEGG) (Saadeldin et al., 2020; Sherman et al., 2022). The R programming language (www.r-project.org/) was used to construct a bubble plot using the ggplots2 package for GO terms. KEGG pathway annotation was performed using a bar plot (www.bioinformatics.com.cn/plot_basic_pathway_enrichment_second_class_summary_bar_plot_206). A protein–protein interaction (PPI) network was generated using the identified genes, among which the string plugin in Cytoscape recognized the interactions with a confidence score of 0.4. Nodes represent the genes, and edges represent the network interconnecting the nodes.
ClueGO v2.5.9 plugin was used for GO analysis with a kappa score of 11, a p-value of 0.05, and a GO term score of 0.4. CytoHubba was used for hub gene identification, and MCODE was used for module construction with a degree cutoff of 2, a node score cutoff of 0.2, and a K-core of 2. Furthermore, we analyzed the interactions and predicted the genes interacting with the IOV genes using GeneMANIA (https://genemania.org/) (Mostafavi et al., 2008).
Effects of IOVs on somatic cells
Porcine kidney fibroblasts (passage #2) were seeded in a four-well plate (1 × 105 cells/cm2) and cultured in Dulbecco's modified Eagle's medium (DMEM) with 10% (v/v) fetal bovine serum and 75 mg/mL kanamycin until 90% confluency. Cells were incubated in DMEM with 2% fetal bovine serum, supplemented with or without IOVs (1 × 108/mL) for 14 days.
Immunofluorescence
Immunofluorescence staining for Oct4, Cdx2, and vimentin was performed as described previously (Saadeldin et al., 2022) with slight modifications. Cells were fixed in 4% paraformaldehyde, washed with PBS, and then permeabilized with 0.1% Triton X-100. Cells were blocked using 1% goat serum for 30 minutes at 23°C. The primary antibodies against Oct4 (mouse monoclonal IgG2b, sc-5279; Santa Cruz Biotech, Inc., Santa Cruz, CA), Cdx2 (rabbit monoclonal IgG, ab76541; Abcam, Seoul, Korea), and anti-vimentin (rabbit monoclonal [EPR3776] recombinant anti-vimentin antibody, ab92547; Abcam) were diluted (1:100 in PBS). Cells were incubated with the diluted primary antibodies for 1 hour at 38.5°C, washed in PBS three times, then incubated with the secondary antibodies (Alexa Fluor 488 goat anti-mouse IgG, A11001, and Alexa Fluor 568 goat anti-rabbit IgG, A11011; Invitrogen, Life Technologies Corp., Eugene, OR), and stored in PBS for 1 hour at 38.5°C. Cells were finally washed three times with PBS, and the nuclei were counterstained with VECTASHIELD Antifade Mounting Medium containing 4′,6-diamidino-2-phenylindole (DAPI; Vector Laboratories, Burlingame, CA) for 5 minutes.
Imaging was performed using a fluorescence microscope at an excitation wavelength of 488 nm for Oct4 and 568 nm for Cdx2 and vimentin. The fluorescence intensity and pixel analysis were analyzed using ImageJ 1.53k (National Institutes of Health, Bethesda, MD).
Total RNA extraction, reverse transcription, and real-time PCR
Total RNA was extracted from IOV-treated and control cells using the RNeasy Micro Kit (Qiagen GmbH, Hilden, Germany) (four replicates). The concentration and quality of RNA were determined using a NanoDrop 2000 (Thermo Scientific). Reverse transcription was performed using the 2 × RT Pre-Mix QuantiNova Reverse Transcription Kit (Qiagen) with 100 ng of total RNA. Real-time polymerase chain reaction (PCR) was performed using a CFX Connect Real-Time PCR System (Bio-Rad) and SYBR 2 × Real-Time PCR Pre-Mix (BioFACT, Daejeon, Korea). Details of the primers used to examine genes and amplicon sizes are shown (Supplementary Table S1). Target-gene expression was compared with housekeeping gene (GAPDH) expression using the ΔΔCt method (Livak and Schmittgen, 2001; Saadeldin et al., 2023).
Statistical analysis
Values were presented as mean ± standard error of the mean (SEM) and analyzed using an unpaired Student's t-test. Differences were considered significant at p < 0.05.
Results
Isolation and characteristics of IOVs
We successfully isolated IOVs using a micromanipulation procedure (see methods and Fig. 1A). IOVs showed typical spherical morphology and the diameters of IOVs ranged from 63 up to 624 nm with an average diameter of 186.3 ± 10.8 nm (Fig. 1B). We further determined the average concentration of pooled IOVs through NTA for further treatment experiments as shown below. When IOV pellets were diluted 20 times, the average concentrations were 8.6 × 108 particles/mL. Transmission electron microscopy indicated that each IOV is enclosed by a lipid bilayer membrane (Fig. 1C).
Protein cargo contents of IOVs
Mass spectrometry detected 467 and 1591 protein reads in IOVs and whole oocytes, respectively, using their respective UniProt accession numbers (Supplementary Tables S2 and S3). After excluding the redundant reads covering the same protein, the whole oocyte contained 1498 proteins while IOV proteome includes 411 proteins, all of which were observed in the oocyte proteome list (Fig. 2). The top 20 IOV proteins are listed in Table 1.
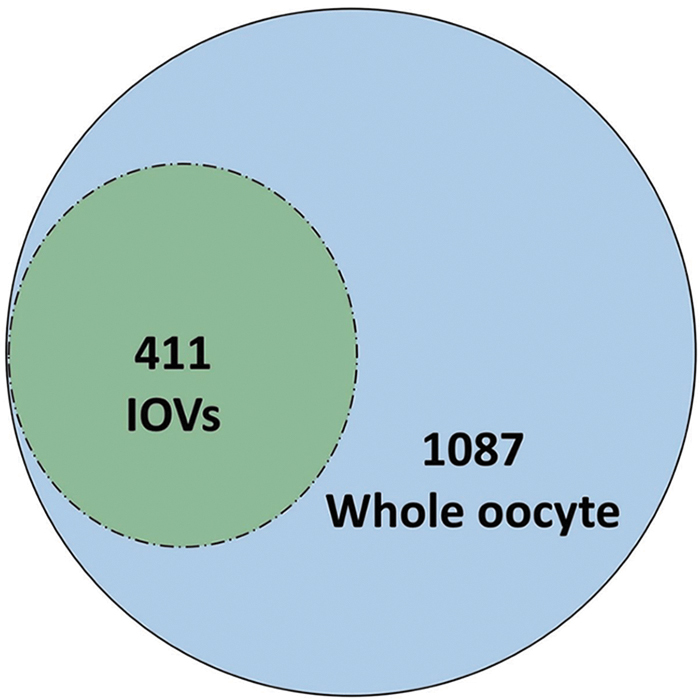
Proteins detected in IOVs and whole oocytes.
Top 20 Proteins Detected in the Intra-Ooplasmic Vesicles
Bioinformatic analysis and PPI
Functional analysis revealed that the 411 genes detected in IOVs were highly enriched in various BP, including single-organism catabolic processes (GO:0044712), carboxylic acid metabolic processes (GO:0019752), and protein folding (GO:0006457). Highly enriched CC included cytosol (GO:0005829), proteasome complex (GO:0000502), and endopeptidase complex (GO:1905369). MF-enriched genes were involved in identical protein binding (GO:0042802), unfolded protein binding (GO:0051082), and isomerase activity (GO:0016853) (Fig. 3A, B). This set of genes was further viewed as information regarding the network using PPI, as shown in Figure 4. The network was further processed for downstream analysis. The ClueGO plugin for GO analysis revealed that our network was highly enriched in the negative regulation of fertilization and proteasome complexes and positive regulation of translation (Fig. 5A). KEGG analysis of ClueGO revealed that the genes were highly enriched in proteasome-related and RNA degradation pathways (Fig. 5B).

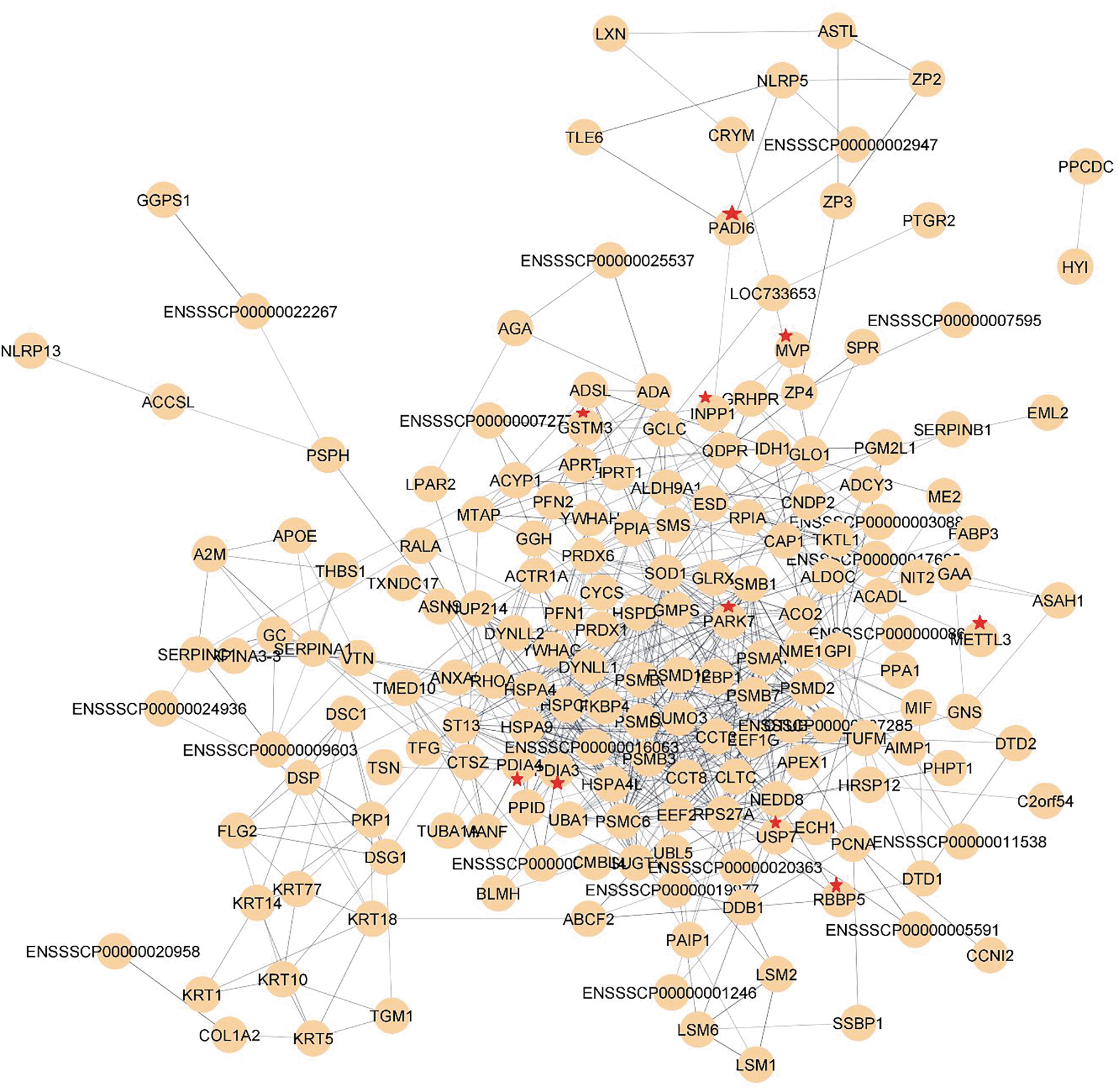
Complete gene network based on PPI generated using the total number of 411 recognized proteins. The PPI network was constructed using a string plugin from Cytoscape. Nodes are connected to one another using ages. The red star-labeled proteins are also referred to (Table 2) as a part of the reprogramming factors. PPI, protein–protein interaction.

Hub gene analysis indicated that SOD1 and HSPA9 were the two top hub genes based on the degree of connectivity, followed by RP227 and HSPCB (Fig. 6A). Two important modules were identified using the MCODE plugin (Fig. 6B, C, highlighting their relationships within the network). Furthermore, GeneMania results showed ∼46% of physical interaction and 25% coexpression between the IOV proteins and the known transcriptional factors of reprogramming, Oct4, Sox2, Klf4, Myc, and Lin28 (Supplementary Fig. S1).

Effect of IOVs on the cultured somatic cells
Extended culture and IOV supplementation resulted in the formation of cell aggregates (Fig. 7) and reduced cell diameters as evidenced by vimentin staining (Fig. 8). Cells treated with IOVs expressed OCT4 and CDX2 (pluripotency and trophoblast markers, respectively) (Fig. 9), which was approximately eight-fold higher than that in the control nontreated cells after 7 days of IOV supplementation. Real-time PCR analysis revealed increased expressions of embryonic stem cell markers OCT4, SOX2, KLF4, SALL4, and the trophectoderm marker CDX2, in addition to increased expression of YBX3, and ZEB2 (Fig. 10) (p < 0.05).

Morphological appearance of extended cultured fibroblasts supplemented with IOVs for 14 days. Images were taken on days 1, 7, and 14. Aggregated cells were observed on day 14 (white arrow). Scale bar = 200 μm.
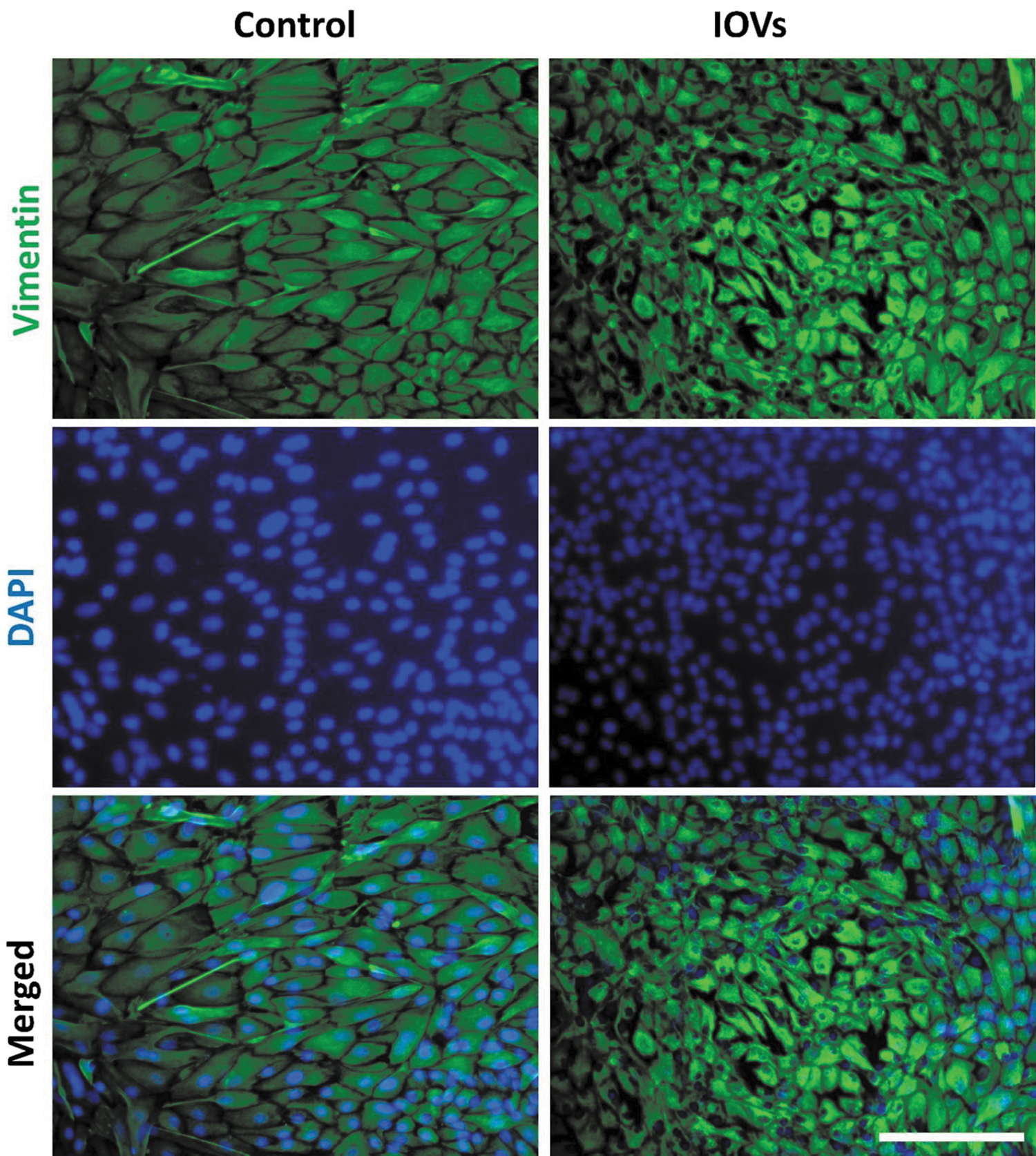
Vimentin expression in extended cultured fibroblasts supplemented with IOVs for 14 days. Cells were incubated with primary rabbit antibodies against vimentin, and secondary goat anti-rabbit antibodies labeled with Alexa Fluor 568. Nuclei were counterstained with DAPI. Scale bar = 200 μm. DAPI, 4′,6-diamidino-2-phenylindole.

The effects of supplementing IOVs on cultured somatic cells.
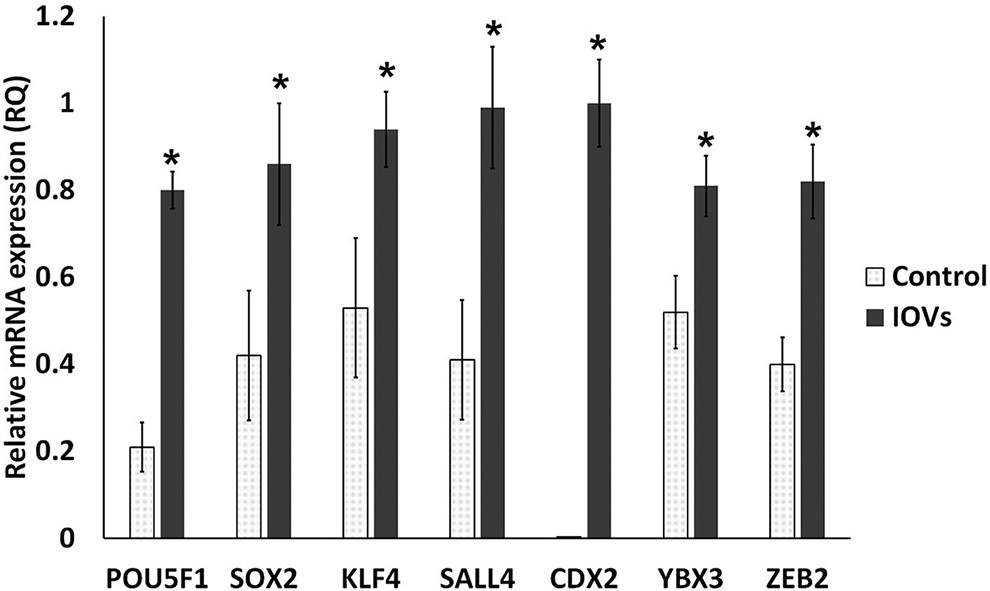
Real-time quantitative PCR comparison of mRNA relative expression (fold change). Comparison between the control and IOV-treated cells after 5 days for culture in three biological and three technical replicates. Asterisk (*) indicates significant difference when p < 0.05.
Discussion
We successfully isolated and characterized nanovesicles from mature porcine oocytes and described them for the first time as “intra-ooplasmic vesicles (IOVs)” and showed detailed proteomics analysis of the isolated IOVs.
Oocyte vesicles have been observed in numerous mammalian species, including pigs (Cran et al., 1980; de Paz et al., 2001; Ducibella et al., 1988; Falconnier and Kress, 1992; Hyttel et al., 1986a; Petr et al., 2001; Sá et al., 2011). All these studies were based on a morphological assessment of oocyte vesicles in different follicular and developmental stages using whole oocyte TEM preparation techniques. The observed vesicles manifested as sizable, heterogeneous, translucent cellular structures, and the diameter widely ranged from 30 nm to 3 μm. The size of these vesicles was proportionally related to the maturation of oocytes (Hyttel et al., 1986b; Reader et al., 2017; Reader et al., 2015). However, no information regarding the accurate isolation and functional analysis of the oocyte vesicles has been demonstrated.
In the current work, the fibroblast cellular morphology has been altered by extended IOV treatments in a mesenchymal-to-epithelial transition (MET)-like process with a relative reduction in cell size and formation of cell aggregates. It has been reported that mesenchymal cells appear to have epithelial-like morphology a few days after transduction with reprogramming factors (Takahashi and Yamanaka, 2006) in a process of MET, which frequently occurs during pluripotency and early embryogenesis (Li et al., 2010). Investigations of switching cells from MET to epithelial-to-mesenchymal transition (EMT) may yield detailed molecular insights into cell fate decisions, not only for switching between epithelial and mesenchymal cells but also for other cell types (Chen et al., 2011; Guo et al., 2014).
Induction of OCT4 was also observed in a previous study (Byrne et al., 2003), which reported that mouse nuclei could be directly reprogrammed by Xenopus oocytes to express Oct4. However, the expression CDX2 requires further investigation of the pluripotency or oligopotency of the reprogrammed cells.
Interestingly, CDX2 is expressed in the intestinal epithelium during development and regulates its patterning of the intestinal epithelium (Grainger et al., 2010) and its overexpression has been associated with cellular differentiation (Qualtrough et al., 2002). Real-time PCR results showed increased expressions of two regulators of EMT; ZEB2 is a master regulator of EMT, and ZEB2-mediated transcription is a key mechanism that transforms epithelial-like trophoblasts into cells with a mesenchymal invasive phenotype (DaSilva-Arnold et al., 2019; Vandewalle, 2005). Moreover, YBX3 induces EMT by activating the PI3K/AKT pathway (Fan et al., 2021).
IOVs may aid in acquiring pluri- or multipotency of the cells in addition to the metabolic reprogramming that is associated with genomic reprogramming. Among the top 20 proteins of IOVs are some proteins that play important roles in nuclear and metabolic reprogramming such as PARK7 (Miyamoto et al., 2011; Zhao et al., 2015), PDIA3 (Yang et al., 2014), and PADI6 (Coonrod, 2008).
One of the key players in the reprogramming process is the set of transcription factors used to induce pluripotency in somatic cells. These factors, including OCT4, SOX2, KLF4, and MYC activate pluripotency-related gene expression and repress lineage-specific gene expression (Pillai et al., 2021). Another important aspect of the reprogramming process is epigenetic regulation, which involves the modification of DNA and histone proteins that regulate gene expression. The reprogramming process involves erasing existing epigenetic marks that define the cell lineage and establishing a new epigenetic landscape characteristic of a new cell type (Xiong et al., 2019).
Understanding cellular reprogramming is important not only for the field of regenerative medicine but also for disease modeling and drug discovery. The reprogramming process can also be targeted to develop drugs that modulate cell reprogramming, providing new avenues for disease treatment (Cherry and Daley, 2012; Goya et al., 2018).
Other proteins identified in this study are involved in antioxidative defense. These proteins include GSTM3, SOD1, and PRDX2, which are indirectly involved in metabolic and pluripotency reprogramming (Dannenmann et al., 2015; Zhao et al., 2015).
Furthermore, several IOV proteins identified in this study are related to the cytoskeleton, which is a network of protein fibers that provides the cell with its shape and internal organization. These proteins included ZP3, DYNLL1, CALR, DYNLL2, KRT14, and KRT5. Heat shock proteins, which protect cells from stress, were among the top 20 proteins. These include HSPA5, HSPE1, and HSPA8. Finally, some IOV proteins were involved in metabolism, including FABP3, SMS, ALDOC, ALDH7A1, and ALB (Table 2).
Epigenetic and Pluripotency Reprogramming Proteins in the Intra-Ooplasmic Vesicles
Conclusion
We reported a detailed method for the isolation and molecular cargo content of proteins from the IOVs of porcine oocytes. The protein contents of IOVs contribute to the alteration of fibroblast morphology after extended culture into an MET-like state with the expression of early embryonic stem cell markers (OCT4, SOX2, KLF4, and SALL4), and the trophectoderm marker CDX2, as well as the EMT regulators (YBX3 and ZEB2). Overall, these results provide some insights into the functional characteristics and molecular interactions of IOV proteins and highlight their potential roles in various biological processes, including reprogramming, antioxidative defense, cytoskeleton, heat shock proteins, and metabolism. Furthermore, these findings provide a rationale for further characterization of IOVs, investigation of the potential reprogramming capabilities for EMT, and the generation of induced pluripotent or oligopotent stem cells.
Authors' Contribution
I.M.S.: conceptualization, methodology, software, data curation, writing—original draft preparation, and funding acquisition. S.B.: methodology and data curation. A.Y.M.: methodology and data curation. S.H.Y.: methodology and data curation. S.I.K.: methodology and data curation. S.L.: methodology, data curation, and investigation. J.C.: conceptualization, methodology, software, data curation, writing—original draft preparation, and funding acquisition.
Footnotes
Acknowledgments
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This work was supported by the Ministry of Science and ICT through the National Research Foundation of Korea (NRF) (grant numbers: 2021R1A2C2009294 and 2022R1I1A1A01065412) and the Brain Pool Program (grant number: 2021H1D3A2A02040098).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.