
Abstract

Lay Summary
Introduction
H
This multifunctional nature of HBx implies its role in altering several cellular mechanisms by hijacking the host cell homeostasis in favor of cancer initiation and evolution (Sivasudhan et al., 2022). HBx is known to activate the regulator transcription factor MYC (Terradillos et al., 1997) and prolongs its cellular levels by inhibiting its ubiquitination and proteasomal degradation (Kalra and Kumar, 2006). HBx is also involved in the regulation of gene expression through epigenetic mechanisms.
For example, HBx relieves chromatin-mediated transcriptional repression of cccDNA of hepatitis B virus involving SETDB1 histone methyltransferase (Andrisani, 2021; Rivière et al., 2015). Wang et al. reported in 2017 that the HBx protein promotes the “stem-like” properties of OV6+ cancer cells in hepatocellular carcinoma.
It is now well established that pluripotency can be induced in mature/well-differentiated cells using Yamanaka's factors (YFs)—a cocktail of transcription factors OCT4, SOX2, KLF4, and c-MYC. Pluripotent stem cells generated using YFs are known as “induced Pluripotent Stem Cells” (iPSCs) (Anonymous, 2016; Yamanaka, 2012).
The use of MYC, a well-known proto-oncogene, is often seen as a concern. In fact, each one of them (MYC, KLF4, SOX2, and OCT4) has been implicated in various cancers (Kim et al., 2015; Miller et al., 2012; van Schaijik et al., 2018; Wei et al., 2006) and all are essential for normal growth and development. The process of generating iPSCs with YFs is not very efficient despite its relative simplicity (Brumbaugh and Hochedlinger, 2013; Winzi et al., 2014; Zhang et al., 2012).
This is especially true for clinical samples obtained in small quantities where the number and quality of the cells could be poor (for example from a bio bank). The efficiency of the iPSC derivation decreases with increasing cell passage number and other parameters such as the age of the donor (Trokovic et al., 2015). Similarly, fibroblast samples from some individuals are very difficult to reprogram. Epigenetic modulators are known to increase the efficiency of iPSC derivation (Kang et al., 2014).
Considering the role of HBx in cell proliferation, epigenetic pathways, protein stability, MYC activation, and, more importantly, the fact that HBx imparts “cancer stem cell” properties to transformed liver cells, we tested our hypothesis that the addition of HBx in the Yamanaka's reprogramming cocktail could enhance the efficiency of iPSC generation. We report here that HBx can not only augment the iPSC generation, but also could substitute MYC in the cocktail.
Materials and Methods
Cloning of Hbx
A synthetic sequence corresponding to 2A peptide from Thoseaasigna virus capsid protein and BspTI (AflII) restriction site was first cloned to pCXLE-EGFP (Plasmid No. 27082; Addgene, www.addgene.org) (Okita et al., 2011). The clones were verified by sequencing and subsequently digested with AflII (Cat. No. ER0831; Thermo Fisher Scientific, Waltham, MA) to prepare the vector.
Hbx insert was polymerase chain reaction (PCR) amplified from pcDNA3-HBV (Gao et al., 2004) with Phusion DNA polymerase (Cat. No. M0530S; New England Biolabs) according to the manufacturer's protocol, gel purified using the GeneJet gel extraction kit (Cat. No. K0691; Thermo Fisher Scientific), and digested with AflII. The vector and insert (1:2) were ligated using T4 ligase following the manufacturer's instructions (Cat. No. EL0011; Thermo Fisher Scientific). The cloned plasmid (hereafter denoted as pHBx, a mammalian expression in which HBx gene sequence was cloned), was verified by sequencing, by transfecting HEK293 cells, and by verifying the protein expression (Supplementary Data S1a–c).
Cell culture
HEK 293 cells were cultured in 10% fetal bovine serum (FBS), Dulbecco′s modified Eagle's medium (DMEM) from CellClone, Genetix Biotech Asia, New Delhi, Cat. No. CCS-500-SA-U, supplemented with 1 × penicillin (100 U/mL) and streptomycin (100 μg/mL) (Cat. No. A018-5X100ML; Hi-Media, Mumbai).
At around 80% confluence, to validate the gene expression the plasmid (vide-supra) was transfected into HEK293 cells using PEI (Cat. No. 49553-93-7; Sigma-Aldrich/Merck) following an optimized version of the standard protocol (Rajendra, 2018) with a DNA:PEI ratio of 1:3. The transfected cells were harvested by scrapping out on the third day for downstream applications.
A variety of fibroblast lines available in our lab that were grown following the established protocols from patient samples as well as healthy volunteers (approved by the institutional ethics committee whenever applicable or obtained from commercial sources such as biobanks) were used for the experiments (Rittié and Fisher, 2005). The derivation of fibroblasts from certain clinical samples resulted in poor-quality fibroblasts, either a result of volume or quality of the specimen, because of age of the donors or other reasons (some of these fibroblasts showed the signs of senescence).
Immunofluorescence/immunohistochemistry/staining
Cells grown on the chamber slides were fixed with ice-cold methanol (Cat. No. 34860; Sigma-Aldrich/Merck) for 5 minutes at room temperature and then gently washed with phosphate buffered saline (PBS). Cells were incubated overnight in blocking buffer that contained the primary antibodies: OCT3/4, SSEA4, TRA-1-80, cytokeratin-18, HNF-4α, ASGPR1 (1:200; SC-5279, SC-21704, SC-21706, SC-32329, SC-374229, and SC-393849; Santa Cruz Biotechnology, Inc., Santa Cruz, CA), GATA4 (mAb No. 36966; Cell Signaling Technologies, MA) antibody against human vimentin (pre-diluted, Cat. No. PM104; PathnSitu Biotechnologies, CA), secondary goat anti-mouse Alexa Fluor® 568 (Cat. No. ab175473; Abcam), and anti-mouse Alexa Fluor-488 (Cat. No. 115-545-062; Jax Immune Research, ME).
More details are provided in the CTAT Supplementary Data S2. After blocking with 5% FBS in TBST for 2 hours, cells were rinsed with TBST and incubated with appropriate secondary antibodies for 4 hours at room temperature in the dark. The nuclei were counterstained with DAPI. Images were acquired using fluorescence microscopes from Nikon Eclipse Ti-S or Evos M5000 (inbuilt gray scale camera and therefore all images are pseudocolored).
Proprietary software associated with microscopes were used to acquire images. Inkscape 1.12 (inkscape.org), ImageJ (NIH, DC), Paint (Microsoft, CA), and Irfanview 4.6 (www.irfanview.com) were used to view, analyze, adjust/improve, label, and layout the images and figures. Flow cytometry data were acquired by FACS Aria III, and FACS data were analyzed by Flowjo 10.4 (FlowJo LLC, Oregon).
Generation of iPSC
iPSCs were generated following established protocols (Anonymous, n.d.; Muthusamy et al., 2016; Ohnuki et al., 2009; Vaidyanath et al., 2020). Fibroblasts (high passage—over 15 passages and low passage—under 5 passages) were cultured on gelatin-coated cell culture plates.
On the fourth day, they were electroporated with Yamanaka's plasmids (plasmid nos. 27078; [human SOX2 and KLF4], 27080 (human L-MYC, LIN28), 27077 (human OCT3/4, shRNA against TP53) from Addgene, www.addgene.org) (Okita et al., 2011) plus or minus pHBx on Amaxa Nucleofector (Lonza, Cologne, France) using the manufacturer's protocol for human dermal fibroblasts.
We have also transfected the two groups of fibroblasts with pHBx alone. The electroporated cells were seeded on to ES qualified Matrigel™ (Product No. 354277; Corning, NY) coated six-well plates (∼105 cells/cm2) with 10% FBS containing DMEM. The media was changed after 2 days to Essential 6 (Cat. No. A1516401; Invitrogen) supplemented with FGF2 20 ng/mL (Cat. No. 100-18B; Peprotech), and the media was changed every 2 days till iPSC colonies started to appear.
Picking of iPSC colonies and their maintenance
When colonies appeared (range: day 18 to 30, from citing of the earliest “nidus” appeared to the appearance of the last colony on any of the culture dishes), they were allowed to grow for 2 days to allow them to a size convenient for manual picking (when they reached about 200–300 μm size) and the picked colonies were allowed to grow larger and they were “passaged” by mechanical splitting into a new Matrigel coated plate containing mTeSR™1 media with its supplement (Anonymous, n.d.) and maintained as suggested by the manufacturer (Cat. No. 85850; StemCell Technologies, Canada). Subsequent passages were done by enzymatic dissociation (Accutase®, Cat. No. A11105-01; ThermoFisher Scientific, MA) when the colonies reached 50%–60% confluence.
Western blot
Whole-cell protein extracts were prepared from scraped out HEK 293, fibroblasts, or iPSC culture plates and extracted using RIPA lysis and extraction buffer. Extracts were run on 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene difluoride membrane using a transfer apparatus following the standard protocols (Bio-Rad, Hercules, CA).
After incubation with 5% nonfat milk in TBST (10 mM Tris, pH 8.0, 150 mM NaCl, 0.5% Tween 20) for 60 minutes, the membrane was washed once with TBST and incubated with rabbit antibodies against human OCT-3/4 antibody (Cat. No. SC-5279; Santa Cruz Biotechnology, Inc.), 1:1000 dilution; mouse anti-HBx antibody (Cat. No. MA1–081; Thermo Fisher Scientific), 1:1000, overnight incubation at 4°C.
All blots were standardized with GAPDH as the internal reference protein. Mouse anti-GAPDH antibody (Cat. No. 60004-1-Ig; Proteintech) was used at dilution 1:1000 at 4°C overnight. The membrane was washed with TBST buffer and incubated with a 1:5000 dilution of horseradish peroxidase-conjugated anti-rabbit (Cat. No. SC-2004; Santa Cruz Biotechnology, Inc.)/anti-mouse antibodies (Cat. No. SC-2005; Santa Cruz Biotechnology, Inc.) for 2 hours at room temperature.
Blots were washed with TBST four times and developed with the ECL system (Cat. No. 170–5060; Bio-Rad), according to the manufacturer's protocols. Chemiluminescence imaging was done using iBright 1500 (Thermo Fisher Scientific).
Real-time polymerase chain reaction
Total RNA was isolated using NucleoZOL (Cat. No. 740404.200; Takara, Shiga, Japan) following the manufacturer's instruction. cDNA was prepared from (deoxyribonuclease treated) total RNA using RevertAid Reverse Transcriptase (Cat. No. EP0441; Thermo Fisher Scientific) following the manufacturer's instructions. Real-time quantitative polymerase chain reaction (RT qPCR) was done using specific oligonucleotide primers targeting the genes of interest (Supplementary Table S1), denaturing temperature 95°C, annealing temperature 60°C, extension temperature 72°C in triplicates and two repeats, using GoTaq® qPCR Master Mix (Cat. No. A6001; Promega) following the manufacturer's instructions on a Veriti Thermo Cycler (Applied Biosystems, MA).
The data were acquired using the software associated with the same machine (ViiA7 V1.2), and relative quantification with respect to the housekeeping genes was calculated using the by 2 (–ΔΔCt) method.
Screening of iPSC colonies
To identify and estimate the efficiency of iPSC colony formation from fibroblasts, the six-well plates were stained for the activity of alkaline phosphatase as iPSC colonies are known to produce alkaline phosphatase (Vaidyanath et al., 2020). We used BCIP/NBT tablets (Cat. No. B5655; SigmaFast Sigma-Aldrich/Merck) as a substrate for alkaline phosphatase that produces a deep purple-black precipitate when alkaline phosphatase is present.
The plates were imaged after 3 weeks of transfection. To estimate the efficiency of iPSC colony formation from fibroblasts, purple-black spots larger than 50 μm were counted in four fields (4 × objective).
Characterization of iPSC colonies
The picked colonies were passaged over 15 times to check whether the colony morphologies retain the acceptable morphology under light microscopic examination. The iPSC colonies were tested for the expression of alkaline phosphatase (Cat. No. A14353; Invitrogen), OCT3/4, SSEA4, and TRA 1-81. Differentiation of iPSC: iPSC cells were aggregated to form embryoid bodies (EBs) that were differentiated to ectodermal, endodermal, and mesodermal lineages following the established protocols.
The germ layers were evaluated using specific markers by immunofluorescence microscopy (Ohnuki et al., 2009; Muthusamy et al., 2014; Baghbaderani et al., 2016). The iPSCs were subsequently differentiated to “hepatocyte like cells” following a modified version of the protocol described by Basma et al. (2009).
Genomic DNA was isolated from iPSC using DNEasy blood and tissue kit (Cat. No. 69504; Qiagen, Germany). DNA PCR: to verify persistence/integration of HBx DNA in iPSC lines: A PCR was done to investigate the persistence (as an episome) or integration (genomic) of the transfected HBx DNA in the iPSC derived from the fibroblasts transfected.
The same forward and reverse primers (Ta = 60C) were used for the PCR (Supplementary Table S1). Taq DNA polymerase was used for the PCR following the manufacturer's protocol. The PCR products were resolved on a 1% agarose gel in TAE buffer (Voytas, 2001).
Karyotyping
Briefly when the plates (fibroblasts/iPSC) reached 80% confluence, cells were treated with colcemid for 30 minutes at a final concentration of 0.1 μg/mL with media. The cells were harvested by trypsinization, washed in PBS, and pelleted down by centrifugation at 200 g. The cells were resuspended in 0.075 M potassium chloride and incubated at 37°C for 20 minutes in a volume of 2 mL.
Cells were pelleted down. Eight drops of fresh ice-cold fixative (3:1, methanol: acetic acid) were added to suspend the cells. The volume was brought up to 0.5 mL with the fixative solution and was incubated for 15 minutes at room temperature. This was repeated twice. Following a last spin, the cells were resuspended in 200–300 μL of fresh fixative, dropped onto clean slides, and air dried and a proportion of metaphase spreads were imaged and assembled (Anonymous, n.d.).
Cell cycle, proliferation, and apoptosis analysis
We performed cell cycle analysis and apoptosis assay using flow cytometry with propidium iodide (Cat. No. 11195; SRL, Mumbai, India), a standard nuclear stain (Crowley et al., 2016; Riccardi and Nicoletti, 2006). The cell proliferation assay was done using the standard MTT assay (Cat. No. 11465007001; Sigma-Aldrich/Merck). MTT assay was performed on each well following the manufacturer's protocol.
Briefly, equal numbers of cells were seeded on six-well plates. The test wells were transfected with pHBx, and the control was transfected with GFP-expressing plasmid (Plasmid No. 27082; Addgene). After 3 days post-transfection, transfected cells were treated with low-dose puromycin (0.5 μg/mL) for 6 hours. Untransfected cells with and without puromycin treatment, transfected cells with and without puromycin treatment were subjected to flowcytometry with propidium iodide to estimate apoptosis in each experimental group. pHBx transfected and control cells were also used for cell cycle analysis using propidium iodine.
Statistics
Data are reported as means ± standard deviation whenever applicable from at least three independent experiments. Student's t-test was performed to compare the mean between two groups and one-way analysis of variance when more than two groups were compared. Numerical data were saved and processed using Microsoft Excel (2013) Washington, United States and Graphpad Prism 7.04, Graphstats Technologies.
Results
HBx improves the generation of iPSC
To verify whether the HBx expression vector (pHBx) is competent to express the viral protein HBx in mammalian cells, we transiently transfected HEK293 cells and fibroblasts, and total cell lysate was probed by Western Blot using an anti-HBx specific antibody as detailed. Figure 1A demonstrates that the 17 kD specific band in cell extracts from pHBx transfected cells is absent in mock transfected cells.

Human dermal fibroblasts passage 3 (HDF-P3) were transfected with YFs containing plasmids in the absence (YF) or presence of pHBx (YF+HBX) as described in Okita et al. (2011). Colonies started appearing by day 20 and we screened the plates for the expression of alkaline phosphatase, a common marker used for the screening of iPSC colonies, using NBT, an alkaline phosphatase substrate that gives a dark-colored precipitate. Figure 1B shows that the addition of HBx resulted in an increased number of alkaline phosphatase positive cell clusters (compare 1A and B), suggesting an improvement in iPSC derivation from fibroblasts.
HBx could replace c-Myc in Yamanaka's cocktail
MYC has been reported to be an oncogene, therefore developing a system lacking MYC would be advantageous. Considering the capacity of HBx to induce stemness in liver cancer stem cells, its ability to potentiate and increase the stability of endogenous MYC, we then investigated whether HBx could substitute for MYC in the Yamanaka cocktail. HDF-P3 cells were transfected with YFs without MYC but supplemented with HBx expression vector.
Figure 1B, panel “a” and “c,” shows that fibroblasts transfected with the Yamanaka cocktail or the Yamanaka cocktail in which c-MYC was substituted with HBx produced iPSC colonies with similar efficiency (Fig. 1C). Thus, our data show that HBx can substitute for c-MYC in YFs.
It is well known that samples from high passage fibroblasts (fibroblasts maintained in culture for a long time and undergone multiple rounds of cell divisions) are difficult to reprogram (Trokovic et al., 2015). To test whether the introduction of HBx in Yamanaka's cocktail can improve reprogramming, we transfected high passage fibroblasts (HDF-Px) that failed to give iPSC colonies under standard conditions (Fig. 1B-d) in our previous experiments with YFs in the presence or absence of pHBx.
We observed that these “difficult to reprogram” cells produced iPSC colonies, although the number of colonies were comparatively less (Fig. 1B-e, f). The results are summarized in Figure 1C. In short, although the addition of HBx gave more iPSC colonies, the results were not statistically significant (p = 0.56). However, the difference was significant in high passage fibroblasts (p = 0.01).
We picked these colonies for further propagation and these colonies appeared to have the characteristic human iPSC/ES morphology under a light microscope (Fig. 1D a-c and e that corresponds to a-b and e in Fig. 1B) and iPSC colonies were positive for OCT3/4, a key marker of pluripotent stem cells (Western Blot Fig. 1D-g).
HBx sequence does not integrate to iPSC lines
The integration of HBx sequence into the iPSC genome or the persistence of HBx plasmid is a concern, because HBx is associated with hepatocellular carcinoma. Genomic integration is a theoretical possibility associated with any transfection that involves DNA. We performed a Western Blot to see whether the iPSC generated out of pHBx transfection continues to express HBx protein in the event of plasmid integration or persistence.
Total cell lysate from iPSCs at the fifth passage was analyzed by Western Blot. Our results suggest that the plasmid gets lost as HBx expression was no longer detectable in cells that are reprogrammed to iPSC (Fig. 1Ea).
Next, we checked whether any integrated HBx sequence can be detected by PCR in the iPSC. Using HBx sequence specific primers and genomic DNA isolated from iPSC lines as the template, after 34 cycles of PCR we could not detect any amplicon of the expected size on agarose gel electrophoresis compared with the positive control (pHBx plasmid) (Fig. 1Eb).
The iPSC derived using p-HBx and the fibroblasts used to derive the iPSC were karyotyped and compared to exclude any abnormalities incorporated during the reprogramming process. Figure 1F shows that the two sets are indistinguishable. We have also performed an RNASeq analysis of the iPSC derived using HBx, and the preliminary analysis showed no significant difference with reference to the control (data not shown).
Characterization of iPSC lines
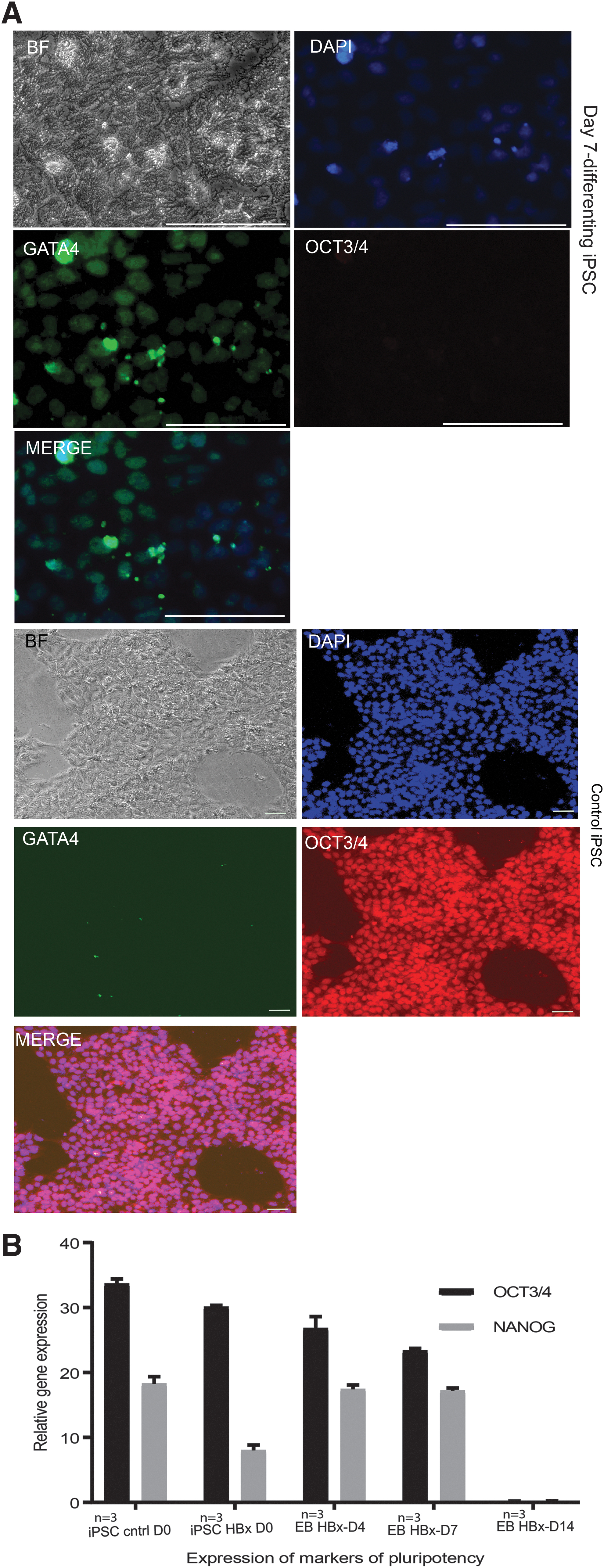
We next characterized the colonies for the expression of the markers of pluripotency, OCT3/4, SSEA4, and TRA 1-81, by immunofluorescence microscopy (Fig. 1G a-c). The iPSCs maintained their typical morphology under a light microscope after 15 passages; they showed characteristic center differentiation (Fig. 2A, right column), formed EBs (middle column), and started differentiation when FGF-2 was withdrawn from the culture medium (left column).

The iPSCs also differentiated into all the three germ layers and accordingly expressed markers specific for each distinct germ layer (Fig. 2B). We analyzed the kinetics of expression of SOX-2, nestin, GATA-4, and brachyury by RT qPCR (Fig. 2C). The expression of GATA4, nestin, and brachury increased from day 0 to 7 and decreased again by day 14.
We further differentiated the iPSCs to hepatocyte like cells (i-Heps). These cells were positive for markers characteristic of hepatocytes-nuclear factors (FOXA2 and HNF4A), cytoplasmic factors (albumin, α-fetoprotein, and coagulation factor-9), cytoskeletal protein (CK18), and surface protein ASGPR1 (Fig. 3a–g).

BF and immuno-fluorescence imaging of hepatocyte-like cells showing lineage-specific markers. The pHBx-derived iPSCs were differentiated to hepatocyte lineage to demonstrate their terminal differentiation potential. These terminally differentiated cells were positive for hepatocyte-specific markers such as FOAX2
Markers of stemness are lost on differentiation
An important determinant of iPSC quality is the ability to undergo complete differentiation. Therefore, one potential concern could be the persistent expression of stem cell markers after the induction of differentiation or incomplete differentiation. Incomplete differentiation raises the possibility of genetic instability and cancer if injected into animals (precluding clinical applications).
Our immunofluorescence imaging showed loss of Oct4 with an increase in GATA4 expression (Fig. 4A). RT qPCR analysis of mRNA from standard iPSCs, iPSCs generated using HBx, EBs, and iHeps (generated from HBx iPSC) showed that no differentiated cells, EBs, or iHeps expressed pluripotent stem cell markers OCT3/4 or NANOG (Fig. 4B).
HBx increased the expression of MYC, OCT3/4 and Ki67
We investigated whether the property of pHBx to substitute for MYC in the Yamanaks's cocktail could depend on its ability to induce MYC or to prolong its half-life. To check whether HBx increases the cellular levels of MYC, we transfected HEK293 cells with pHBx and we found that there is an increase in the levels of MYC on Western Blot.
Considering the promiscuous nature of HBx as a transactivator, we decided to check whether HBx could also increase OCT3/4. Our Western Blots showed an increased level of OCT3/4 in the transfected cells. We further checked whether there is a concomitant increase in the levels of Ki67, a nuclear protein that is associated with cellular proliferation, and as expected we found an increase in the Ki67 levels in pHBx transfected cells compared with the control (Fig. 5A).

These results are summarized in the densitometric plots of Western Blots (Fig. 5B). We used HEK cell line for these experiments, because they are much easier to transfect compared with human dermal fibroblasts (but we confirmed the expression of HBx in both fibroblasts and HEK in the beginning of the reprogramming experiment; refer Figure 1A). We assumed that the mechanism could be the same for fibroblasts (which may not be the case). This is a major drawback of this study.
Cell death at various stages adversely affects the efficiency of iPSC generation. It is well known that during the reprogramming process lots of cells undergo apoptosis and decrease in cell death would have a positive effect in reprogramming efficiency. Cell death ranges from apoptosis of fibroblasts at the time of transfection to apoptotic events during the reprogramming process.
To check this, we did a flow cytometric analysis of pHBx transfected cells, which revealed that in cells transfected with pHBx, cell death decreased significantly (about 12% in control versus ∼6% in transfected cells, p < 0.05). This effect was especially pronounced when cells were exposed to stress (low concentration puromycin treatment for 24 hours).
About 47% cells died in puromycin-treated plates compared with ∼35% in pHBx transfected cells treated with puromycin (p < 0.05) (Fig. 5C). More cells moved toward proliferation: The cell in G2/G1 was more in pHBx transfected cells (G2:13%) compared with control (G2:8.7%; Fig. 5D). An MTT assay was performed (third day post-transfection) to assess the metabolic activity, but the differences between pHBx transfected and control cells were not statistically significant (Fig. 5E).
Discussion
HBx is a multifunctional protein coded by the hepatitis B virus that is critical for the survival of the virus within the human body. It is possible that HBx helps the virus to “reprogram” the epigenetic landscape of hepatocytes to facilitate its replication, and in this process, cells mutate to form hepatocellular carcinoma. On induction of the HBx gene, resting cells begin to enter the S-phase of the cell cycle (Koike, 1995).
Promotion of cell division is important for viral survival and infectivity. HBx is also known to inhibit the ubiquitination and proteasomal degradation of c-Myc (Kumar and Sarkar, 2004). The HBx also induces growth factors, activates MAP and Src kinases, and inhibits apoptosis by multiple mechanisms, including the inhibition of p53 (Kumar and Sarkar, 2004).
HBx is known to be a “broad spectrum trans activator” with the ability to regulate all three classes of promoters (Kumar and Sarkar, 2004). Further, HBx was reported to induce “cancer stem cells” in hepatocellular carcinoma10. Considering these facts, we presumed that the addition of pHBx could increase iPSC generation in cells that are difficult to reprogram such as cells isolated from old patients, primary cells that are poorly characterized, “cell lines” derived from a very small quantity or poor-quality tissue samples (for example, from rare diseases). Therefore, our findings could be useful in iPSC generation facilities such as biobanks or iPSC repositories.
Our studies do not resolve the mechanism by which HBx improves the outcome of iPSC generation. One possibility is that HBx increases the amount of endogenous MYC in the transfected fibroblasts by prolonging the half-life of intracellular MYC. Our experiments demonstrated increased levels of MYC on HBx expression. We have also shown that HBx increases the amount of endogenous OCT3/4, which might improve the process of iPSC generation. It is also possible that HBx affects the cell cycle, especially at later stages of the reprogramming process, increasing the efficiency of iPSC generation.
Another possibility is that HBx decreases the apoptosis at various stages of the reprogramming. Significant cell death follows the transfection (“Nucleofection”) of fibroblasts. HBx protein can drive cells division in stressed cells (Kapoor et al., 2013). Reprogramming transcription factors should get access to their specific DNA sequences on the host chromatin.
In somatic cells, these regions (for example, OCT3/4 binding elements) are highly condensed and getting access to the sites is a limiting step in reprogramming. HBx plays an important role in modulating the epigenetic landscape (Kalra and Kumar, 2006; Rivière et al., 2015). It is possible that HBx could improve the efficiency of reprogramming by facilitating the interaction between the reprogramming transcription factors and their binding elements in DNA by loosening up the heterochromatin.
HBx is a broad-spectrum activator of transcription; therefore, it might enhance the complex interactions between transcriptional networks, decreasing the threshold to establish and sustain the transcriptional networks for the state of pluripotency. HBx is a pleotropic gene, which makes it very difficult to fully elucidate the mechanism of action with reference to reprogramming.
There are concerns regarding the use of MYC (a proto-oncogene) in the generation of iPSC. HBx, as alluded to earlier, is associated with hepatocellular cancer. However, in the iPSC generated using HBx, we did not find the expression of HBx protein, nor could we detect HBx integration by PCR. Further, it is possible that HBx may not be as malignant as originally perceived.
The HBx protein is known to confer resistance against nucleolar stress (Kapoor et al., 2013; Kim et al., 2008). Niu et al. (2017) reported that “transactivation of FXR by HBx may represent a protective mechanism to inhibit inflammation and the subsequent carcinogenesis.” It may be noted that HBV viral infection per se is rarely oncogenic in hepatocarcinogenesis (Niu et al., 2017). In either case, since the iPSC generated does not express HBx, there is little concern regarding the oncogenic potential of HBx in the iPSC generated.
HBx exerts a clearer effect in high passage fibroblasts and this could be because of a few reasons. With higher passage, their cell replication capacity decreases. Further, these aged cells are more prone to apoptosis when subjected to the stress of transfection and reprogramming. HBx as shown in Figure 5C and D improves cell survival on stress and cell division. This might help reprogramming. It is known that when cells age their chromosomes get more condensed. As discussed earlier, HBx relieves chromatin-mediated transcriptional repression, although we do not know whether that is the case here.
Deriving enough cells from small amounts of clinical samples such as fine needle aspiration biopsy (or poor-quality samples) is challenging. Since the starting number of cells are small, they need to undergo more rounds of cell divisions to attain enough numbers for certain experiments. The same is true for cells from higher passages.
During continuous culture, cells acquire chromosomal abnormalities, deletions, mutations, epigenetic changes, and senescence. Here, we found high passage (senescent) fibroblast samples producing iPSC colonies when YFs were supplemented with HBx. The iPSC colonies thus derived were similar (in all aspects we studied) to those that were derived from early passage fibroblasts. This shows that HBx together with YFs can probably reverse an aging (senescent) phenotype, although more experiments are required to prove this point.
We have for the first time demonstrated in vitro that Hbx can increase the cellular levels of OCT3/4. The increased levels of OCT3/4 and MYC are possibly enhancing the generation of iPSC. We have also shown that the iPSC generated using Hbx can be differentiated to all the three germ layers, and terminally to hepatocyte-like cells.
Our findings also have some importance in understanding the pathogenesis of hepatocellular carcinoma. HBx might change the epigenetic milieu favoring dedifferentiation and increase in cell division. Both favor the development of cancer, as dividing cells are more prone to accumulate mutations over time especially when apoptosis is inhibited.
Conclusion
HBx protein increases the efficiency of reprogramming of fibroblasts to iPSC. HBx can substitute for MYC protein in the YFs (OCT4, SOX2, KLF4, MYC). HBx can also improve the generation of iPSC from difficult to reprogram/senescent cell samples using YFs. This may be helpful for biobanks interested in creating iPSC repositories. Our in vitro experiments suggest that HBx induces OCT3/4 (and MYC) expression and aids reprogramming.
These findings also have implications in understanding the mechanisms of pathogenesis of hepatocellular carcinoma caused by HBV. Further, our findings suggest that HBx together with YFs can probably “reverse” senescence under certain conditions.
Footnotes
Acknowledgments
The authors acknowledge Dr. Saurabh Bhattacharya and Dr. Leena Rawai, Lal Path Labs, New Delhi for their help in karyotyping and Dr. Shajo Kunnth, Dr. Anna Velcich, and Mr. Alan Alfieri for their critical comments.
Authors' Contributions
S.M.G. conceived the idea, performed experiments, analyzed and interpreted the results, wrote the manuscript, and obtained funds; S.G. performed experiments on cell culture; R.S. performed all Western Blots; N.V. assisted S.M.G. in cloning pHBx; and S.K.S. reviewed the manuscript.
Availability of Data and Materials
Ethics Approval and Consent to Participate
Authors have taken the required approval from the Institutional Ethics Committee/Institutional Review Board for the human samples used.
Consent for Publication
All the authors have agreed on the content of the manuscript and the corresponding author on the behalf of all authors consent for publication of the manuscript.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
S.M.G. thanks the Department of Biotechnology, Ministry of Science and Technology, India (grant no. BT/PR15116/MED/31/334/2016), Government of India and Science and Engineering Research Board (grant no. ECR/2015/000275).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.