
Abstract
Autologous human fibroblasts have the potential to differentiate into the osteogenic lineage under specific conditions and can be utilized for bone regeneration. However, their efficiency is currently unsatisfactory. Recently, low-intensity nanosecond pulsed electric field (nsPEF) stimulation has been demonstrated to enhance cell pluripotency by activating epigenetic regulatory pathways. In this study, human dermal fibroblasts were exposed to different intensities of nsPEF to assess whether these exposures resulted in changes in proliferation rate, calcium salt deposition, and expression of differentiation-related markers in different experimental groups. The results showed a significant increase in cell proliferation, pluripotency, bone marker expression, and osteogenic differentiation efficiency when stimulating cells with 5 kV/cm of nsPEF. However, cell proliferation and differentiation significantly decreased at 25 kV/cm. Additionally, the proliferation and efficiency of osteogenic differentiation were reduced when the nsPEF intensity was increased to 50 kV/cm. Treatment with a 5 kV/cm of nsPEF led to increased and concentrated expression of Yes-Associated Protein (YAP) in the nucleus. These observations suggest that human dermal fibroblasts possess a heightened potential to differentiate into osteogenic cells when activated with nsPEF at 5 kV/cm. Consequently, the nsPEF strengthening strategy shows promise for fibroblast-based tissue-engineered bone repair research.
Introduction
Human autologous fibroblasts are frequently used in cell-based therapies due to their extensive distribution and exceptional capacity for proliferation (Garcia et al., 2020; Sorrell and Caplan, 2004; Wan et al., 2018). In bone regeneration, fibroblasts can be utilized to construct engineered bone grafts directly, resulting in reduced donor-site morbidity (Sommar et al., 2010). Although fibroblasts were once thought to be terminally differentiated cells, they have demonstrated a strong potential for osteogenic transformation under specific conditions such as trauma, chronic inflammation, or keloid formation (Jiang et al., 2021; Li et al., 2022). This potential plasticity allows for fibroblasts to potentially be used in engineered regeneration of bone tissues, including applications in repairing bone defects.
However, standard procedures for inducing osteogenic differentiation of fibroblasts in vitro remain unsatisfactory due to inefficient conversion of dermal-to-osteogenic lineage under typical culture conditions (Zhou et al., 2008). Due to their status as terminally differentiated cells, fibroblasts exhibit low express levels of genes associated with cellular pluripotency during this differentiation process, resulting in loss of transformation ability. To overcome this issue, it is critical to augment the expression levels of essential functional factors responsible for cell reprogramming artificially. This augmentation would enable fibroblasts to regain their pluripotency. It has been shown that totipotency can be induced by directly infecting human fibroblasts with four defined factors, resulting in the generation of induced pluripotent stem cells that are morphologically, proliferationally, and genetically indistinguishable from embryonic stem cells, and can form teratomas (Takahashi et al., 2007). Nonetheless, alterations in the genome have raised significant concerns regarding the neoplasm formation and ethical controversies (Nakagawa et al., 2008).
In recent years, a new type of electrical field stimulation, low-intensity nanosecond pulsed electric field (nsPEF), has demonstrated superior performance in managing energy pulses with precision and enhancing single-cell bioconversion efficiency, while minimizing electrical harm to cells (Haberkorn et al., 2021). By applying high-voltage electrical stimulation with a short pulse width to cells, intracellular ion channels can be mobilized, leading to the activation of downstream signaling pathways (Haberkorn et al., 2021). It has been shown that nsPEF promotes differentiation efficacy of bone marrow mesenchymal stem cells (BMSCs) by mediating promoter DNA demethylation of vital pluripotency-related genes, such as OCT-4 and Nanog at a relatively low level of voltage intensity (Li et al., 2020). Analogously, low-intensity nsPEF exerted similar effects in enhancing the differentiation potential of somatic cells such as myoblasts and osteoblasts, and have shown its biosafety and cytocompatibility (Hartig et al., 2000; Zhou et al., 2018). However, the prodifferentiation effects of low-intensity nsPEF stimulation on fibroblasts stills require further investigation.
Since the expression of important pluripotency-related gene expression is directly regulated by orchestration of transcriptional factors and epigenetic status (Wang et al., 2017), it is expected that low-intensity nsPEF stimulation (generally <20 kV/cm) could enhance the reprogramming efficiency of terminally differentiated cells in vitro by modifying pluripotency gene expression through epigenetic means. The forced modification of histone or DNA through treatment with low-intensity nsPEF, such as deacetylation or demethylation, can promote somatic cell reprogramming and facilitate the conversions between different cell types or transdifferentiation (Wang et al., 2017). Given this, it was suggested that low-intensity nsPEF could enhance the transformation potential of human dermal fibroblasts by upregulating pluripotency-related gene expression. As a result, it could facilitate effective dermal-to-osteogenic lineage transformation efficacy under conventional osteogenic induction conditions.
Herein, we aimed to investigate the promotive effect of low-intensity nsPEF on the efficiency of osteogenic transformation in human dermal fibroblasts to uncover its potential in future studies of bone tissue engineering and regenerative medicine.
Materials and Methods
Isolation and culture of human dermal fibroblasts
Human dermal fibroblasts were isolated from skin tissues donated by patients undergoing orthopedic surgery at the First Affiliated Hospital, College of Medicine, Zhejiang University. The relevant information pertaining to the donors is listed in the Supplementary Table S1. All procedures were approved by the Ethics Committee of the First Affiliated Hospital, College of Medicine, Zhejiang University. The dermis was diced under aseptic conditions and subsequently digested in a 0.2% collagenase (Solarbio, Beijing, China) solution for 2 hours at 37°C on a thermostatic shaker (Innova40, New Brunswick, Eppendorf, Germany). Then, the tissue samples were collected and rinsed with Dulbecco's modified Eagle's medium (DMEM; Gibco, Thermo Fisher Scientific), and resuspended in DMEM containing 10% fetal bovine serum (FBS; Gibco). The primary cells were cultured in standard Petri dishes and passaged with 0.25% trypsin (Gibco) upon reaching full confluence. Cells from passage 1 to passage 3 were utilized in all subsequent experiments.
Application of nsPEF
To deliver nsPEF stimulation to the cells, the cell samples underwent pulsing within a capacitive pulse generator system illustrated in Figure 1. In short, a high-voltage pulse power supply (SKS series; Matsusada Precision) charged the capacitor system through an RG217 coaxial cable (M17/78-RG217; Times Microwave Systems), leading to the development of a pulsed electric field between the electrodes. Human dermal fibroblasts (1 × 106/mL) were suspended in 1 mL of DMEM culture and transferred to a colorimetric dish with electrodes on both sides. The cells underwent five repetitions of electric field stimulation at 5, 25, and 50 kV/cm, with a 1 Hz frequency and 100 ns pulse width. A 1-second time interval separated each pulse, and cells unaffected by nsPEF served as control. These cells were cultured in either standard DMEM or osteogenic medium for various assays, as described in the subsequent sections.
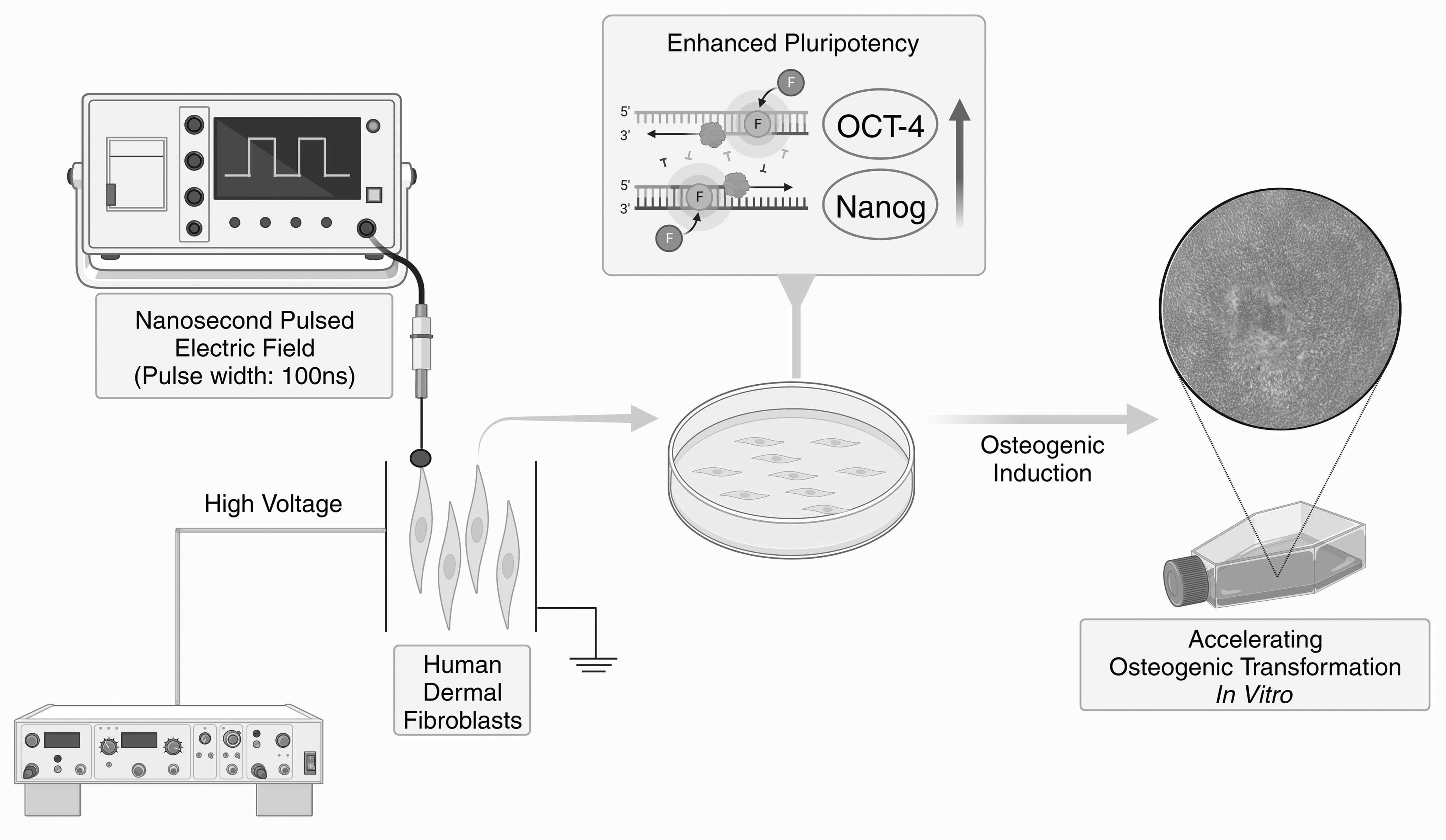
Fibroblasts were stimulated with low-intensity nsPEF through a capacitive pulse generator system. The cellular pluripotency of the fibroblasts was enhanced and their differentiation toward osteogenesis was accelerated in an osteoinductive environment. nsPEF, nanosecond pulsed electric field.
Cell proliferation analysis and migration assay
To analyze the impact of nsPEF on cell viability, cells treated with various intensities of nsPEF (0, 5, 25, and 50 kV/cm) were seeded in 96-well plates at a density of 1000 cells per well. The cells were then maintained in DMEM containing 10% FBS. On the first, third, fifth, and seventh day, cell proliferation was assessed using the Cell Counting Kit-8 reagent (CCK-8; Dojindo Molecular Technologies, Inc., Japan) according to the manufacturer's protocol. Cell proliferation curves were generated by measuring the optical densities of the supernatants in each well using a spectrophotometer (Elx800; Biotek, Germany) at 450 nm. Cells from three individual donors were involved in this assay.
Similarly, the pulsed cells and untreated cells were seeded onto six-well plates at a density of 10,000 cells/cm2 and maintained in the DMEM supplemented with 10% FBS. After reaching complete confluence in the six-well plates, the cells were starved in serum-free DMEM for 8 hours in a serum-free culture condition. Subsequently, the cell monolayer was scratched using a sterile 200-μL pipette tip and incubated in the serum-free DMEM. Images of the scratched cells were captured after scratching and at the indicated time point (24 and 48 hours) using a phase-contrast microscope (CKX53; OLYMPUS, Japan). This assay included cells from three individual donors.
Osteogenic induction differentiation experiment
Cells treated with different intensities of nsPEF (0, 5, 25, and 50 kV/cm) were seeded in six-well plates at a density of 30,000 cells/cm2 and incubated to 90% confluence in the DMEM containing 10% FBS. According to the reported induction protocol (Li et al., 2022), human dermal fibroblasts were induced into the osteogenic lineage by using osteogenic induction medium, which was prepared by adding FBS (10%),
RNA isolation and real-time quantitative PCR
The cells were subjected to various intensities of nsPEF (0, 5, 25, and 50 kV/cm) and subsequently seeded onto six-well plates at a density of 10,000 cells/cm2. Cells were maintained in the DMEM supplemented with 10% FBS. After 2 and 5 days of normal culture or 14 days of osteogenic induction, cell lysis was performed, and total RNA was extracted by using the EZ-Bioscience Quick RNA Extraction Kit (B0004DP, Roseville, MN). RNA samples with A260/A280 ratios of 1.8 to 2.0 were used for cDNA synthesis using 4 × reverse transcriptase (A0010GQ; EZ-Bioscience).
The real-time quantitative PCR was conducted with 2 × Power SYBR Green PCR Master Mix (A0012-R2; EZ-Bioscience) in a real-time thermal cycler (7500fast; Applied Biosystems; Thermo Fisher Scientific) according to a previously described protocol (Tu et al., 2020). For this study, thermocycling procedures were set as: 95°C for 10 minutes, followed by 40 cycles of 95°C for 30 second and 60°C for 30 second. The human housekeeping gene GAPDH was employed as an internal control. The gene expression levels were determined using the 2−ΔΔCt method. The primers for this assay are listed in the Supplementary Table S2. The assay included cells from four individual donors.
Western blot
Total proteins were extracted from human dermal fibroblasts treated with or without nsPEF for 5 days, respectively, using radioimmunoprecipitation assay lysis buffer containing 1% PMSF and 1% phosphatase inhibitor (Beyotime Institute of Biotechnology, Shanghai, China). Cell lysates were resolved on 10% sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE) gels (Solarbio). A mass of 40 μg of protein was loaded in each lane for electrophoresis and then transferred to PVDF membranes. After transfer, the membranes were blocked with 5% nonfat milk (Bio-Rad Laboratories, Inc., CA) at 37°C for 1 hour. Immunoblotting was performed according to previously published protocols (Kaneda-Ikeda et al., 2020). The membranes were incubated with primary antibodies followed by secondary antibodies conjugated to horseradish peroxidase. Details of the antibodies used are listed in Supplementary Table S3. Protein bands were visualized using an enhanced chemiluminescence detection system.
Immunofluorescence staining
After exposure to various intensities of nsPEF, the cells were seeded on glass coverslips at a density of 1 × 104/cm2. Subsequently, the cells were kept in DMEM for 5 days. Before conducting immunofluorescence staining, the cell samples were blocked with PBS that contained 5% serum, and primary antibodies (Supplementary Table S3) were incubated with the cells overnight at 4°C. Following three rinses with PBS, the samples were incubated with secondary antibodies (Supplementary Table S3) at room temperature for 2 hours. Cell nuclei were stained with DAPI (VECTASHIELD Antifade Mounting Medium with DAPI; VECTOR Laboratories, Burlingame, CA). The stained sections were then observed using a Leica confocal microscope (SP8; Leica, Germany).
Statistics
The article presents data as mean ± standard deviation. Statistical differences among various groups were calculated using one-way ANOVA, followed by a Least Significant Difference post hoc test through SPSS software (version 22.0; SPSS, Inc., IL). Differences were considered significant when the p-value was <0.05.
Result
Fibroblasts proliferated faster after treatment with low-intensity nsPEF
In this study, CCK-8 assay was performed to test whether nsPEF treatment can change the proliferation ability of fibroblasts (Fig. 2). Compared with the other groups, fibroblasts treated with 5 kV/cm nsPEF proliferated at the fastest rate (p < 0.05). Conversely, the cell proliferation rate decreased with increasing electric field. The proliferation rate of fibroblasts treated with 25 or 50 kV/cm nsPEF is slower than that of untreated cells (p < 0.05). The CCK-8 assay confirmed that the low-intensity nsPEF can enhance the proliferation ability of fibroblasts.
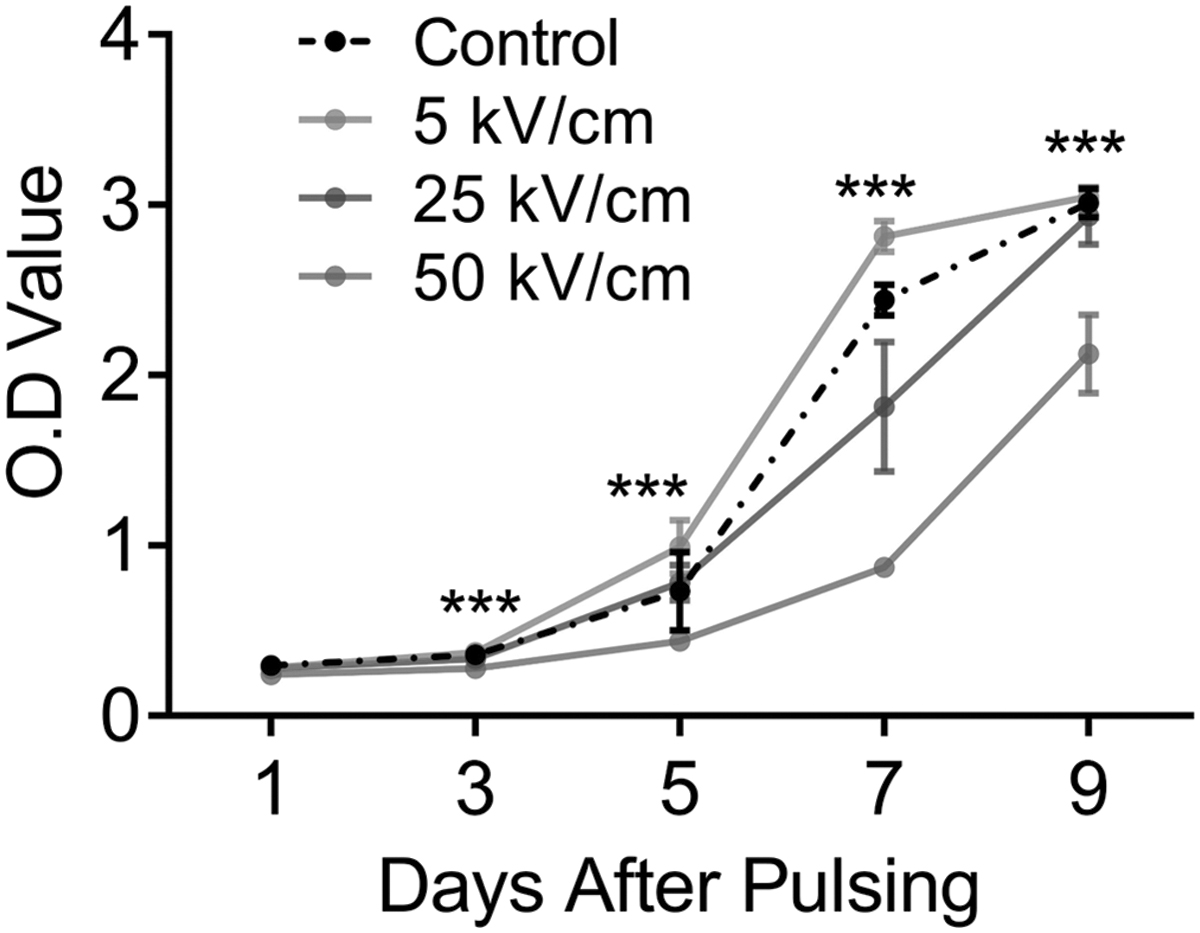
The cell proliferation was assessed on the first, third, fifth, seventh, and ninth day after nsPEF treatment. The absorbance of each group at 450 nm represents the cell proliferation level. ***p ≤ 0.001.
Low-intensity nsPEF facilitated cell adhesion and migration
The adherence ability of fibroblasts in the 5 kV/cm group was close to that of the control group (Fig. 3A). However, the number of stably adhered cells gradually decreased as the electric intensity increased from 5 to 50 kV/cm. After these cells reached 100% confluence, the scratch wound healing assay was performed as previously described (Tu et al., 2019). As illustrated in Figure 3A and 3B, fibroblasts treated with a 5 kV/cm nsPEF exhibited the highest migration speed among the four groups, whereas those treated with 25 or 50 kV/cm nsPEF migrated at a slower rate compared with the cells in the control group over the same time period (p < 0.05). These results suggest that nsPEF enhanced cell migration at the 5 kV/cm level.
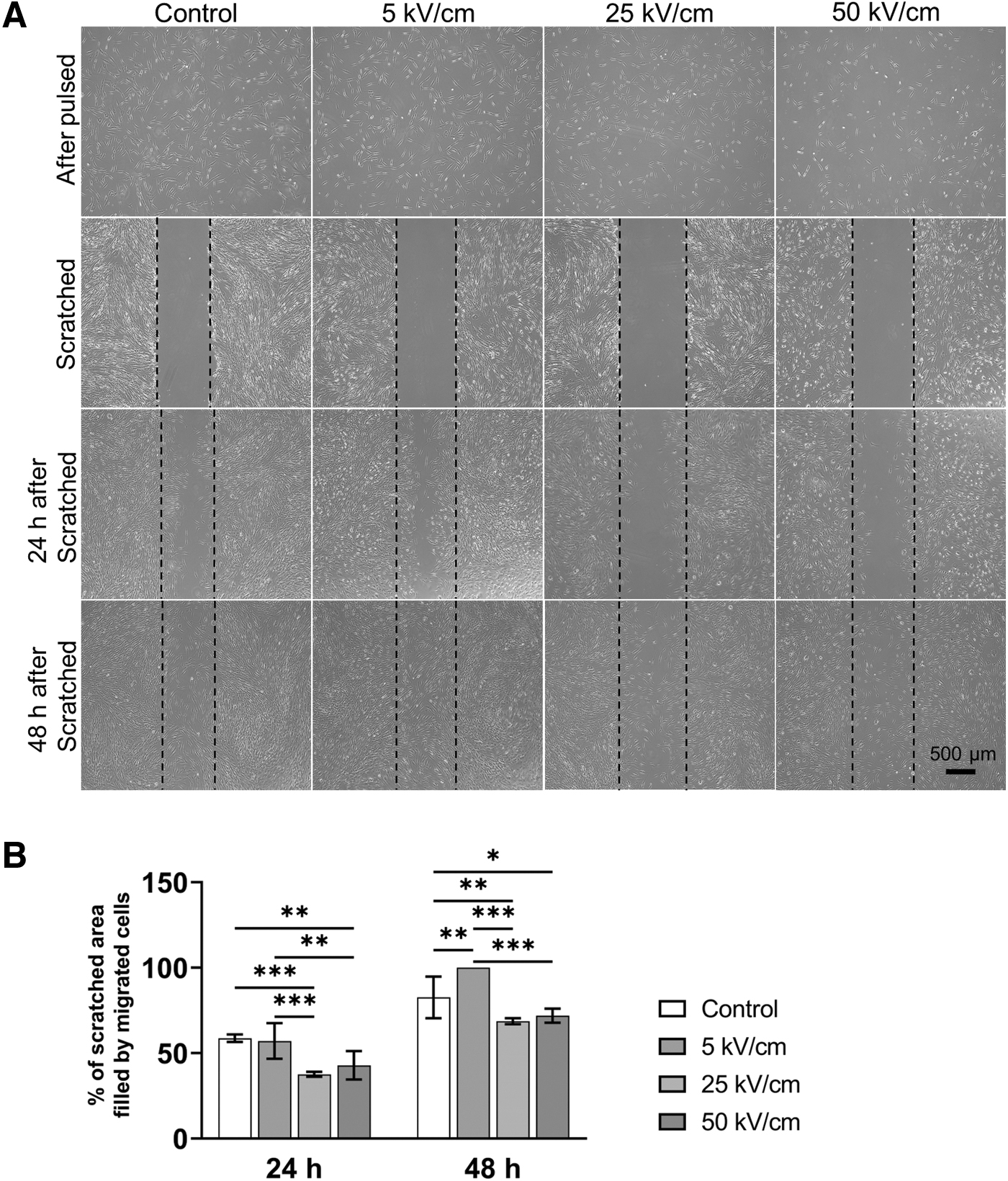
Cell migration capacity was altered after nsPEF treatment.
Low-intensity nsPEF treatment enhances the potential for cells transformation toward osteogenesis
After exposure to various intensities of nsPEF and culturing under normal conditions for 2 or 5 days, the expression levels of genes related to cell pluripotency and multiple lineage markers were analyzed by real-time quantitative PCR (Fig. 4A). Specifically, on day 2, cells stimulated with 5 kV/cm nsPEF showed relatively higher expression levels of OCT-4, Nanog, ALPL, DNMT1, COL1, and COL2 compared with the control group (p < 0.05). Conversely, the expression of RUNX2 was relatively lower (p < 0.05), while no significant differences were observed for OCN, OPG, aggrecan, and SOX-9 (p > 0.05). The expression levels of all genes showed no significant difference between the control group and cells stimulated with 25 kV/cm nsPEF (p > 0.05), except for the Nanog. In the cells stimulated with 50 kV/cm nsPEF, the expression levels of Nanog, ALPL, DNMT1, ALPL, RUNX2, and COL1 were relatively lower (p < 0.05), while there were no significant changes in OCN, OCT-4, OPG, Aggrecan, COL2, and SOX-9.
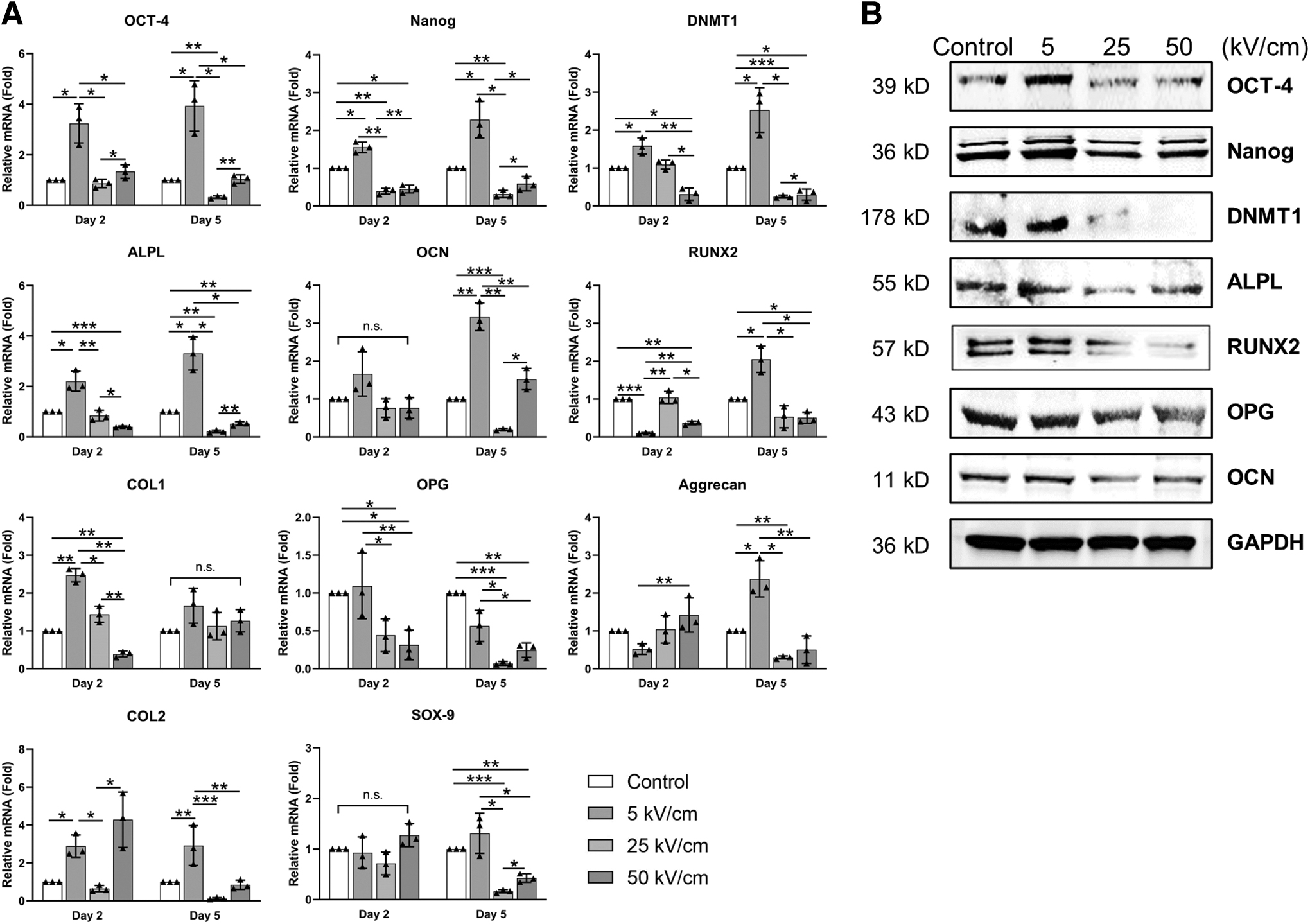
Low-intensity nsPEF treatment enhances cell totipotency and facilitates cellular osteogenesis/chondrogenesis.
On day 5, cells stimulated with 5 kV/cm nsPEF showed higher levels of OCT-4, Nanog, DNMT1, ALPL, OCN, RUNX2, aggrecan, and COL2 (p < 0.05), while there was no significant difference in COL1, OPG, and SOX-9 (p > 0.05). In contrast, the expression levels of OCT-4, Nanog, DNMT1, ALPL, OCN, OPG, aggrecan, and SOX-9 were significantly lower (p < 0.05) in cells stimulated by 25 kV/cm nsPEF, whereas there was no significant difference for RUNX2, COL1, and COL2 (p > 0.05). Cells stimulated by 50 kV/cm nsPEF showed a lower expression level of DNMT1, ALPL, RUNX2, OPG, and SOX-9 (p < 0.05), but no significant difference was observed for OCT-4, Nanog, OCN, COL1, Aggrecan, and COL2 (p > 0.05). Adipogenic differentiation, on the other hand, was largely unaffected by low-intensity nsPEF stimulation, as evidenced by the upregulation of expression of only PPAR-γ and CEB/P-α in 5 kV/cm group on day 2 (Supplementary Fig. S1, p < 0.05).
In summary, 5 kV/cm of nsPEF treatment significantly upregulated the expression of these genes, which is consistent with previous reports (Sen et al., 2010). On the contrary, fibroblasts in 25 kV/cm group and 50 kV/cm group showed relatively lower lineage conversion capacity and attenuated osteogenic phenotype.
Low-intensity nsPEF increased the production of proteins related to osteogenesis
The above changes in gene expression levels were also verified at the protein level. As shown in Figure 4B, western blot revealed that the production level of osteogenesis proteins, such as ALPL, RUNX2, and OCN, were significantly unregulated in the 5 kV/cm group, except for the OPG. However, the expression levels of these genes were significantly downregulated in cells treated with 25 and 50 kV/cm nsPEF (Fig. 4B). Meanwhile, cells stimulated by 5 kV/cm nsPEF showed a significant increase in the expression of OCT-4, Nanog, and DNMT1, which are associated with stem cell pluripotency and differentiation ability. In contrast, the production of these proteins was considerably reduced in cells exposed to 25 or 50 kV/cm nsPEF.
Low-intensity nsPEF improves the efficiency of osteogenic transformation of fibroblasts cultured in vitro
After 14 days of induction, the Alizarin Red S staining revealed the most abundant deposits of red-stained calcium salts in the 5 kV/cm group, indicating enhanced osteogenic phenotype of fibroblasts after the low-intensity nsPEF treatment (Fig. 5A). Obvious osteogenic nodule formation was observed in the 5 kV/cm group, but not in the other groups. Meanwhile, the calcium salt abundance in the 50 kV/cm group was lower than that in the control group (Fig. 5B, p < 0.05). Furthermore, after 14 days of osteogenic induction, the expression of OCT-4 and Nanog was significantly reduced in each nsPEF-treated group, whereas the expression levels of osteogenesis-related markers were significantly increased (Fig. 5C, p < 0.05), especially at the 5 kV/cm level where the increase was most significant.
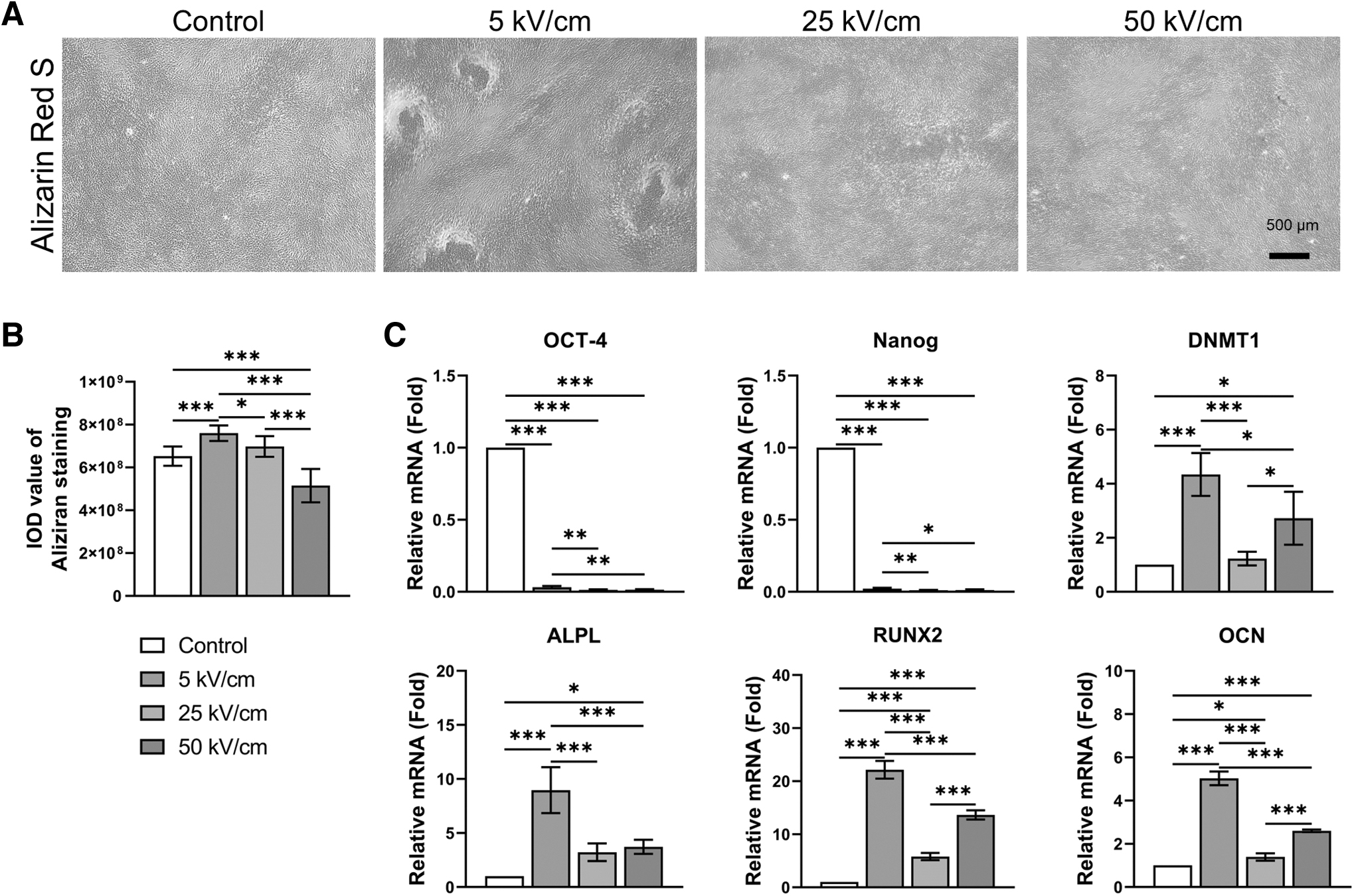
Low-intensity nsPEF enhances osteogenic induction efficiency.
The nsPEF treatment altered YAP phosphorylation levels and its subcellular distribution
Although the expression of total YAP was not affected by nsPEF stimulation, the distribution of YAP within the cell changed significantly after treatment with differing nsPEF intensities (Fig. 6). Compared with the control group, nsPEF treatment at 5 kV/cm resulted in the distribution of YAP closer to the nucleus, whereas nsPEF treatment at 25 kV/cm resulted in a uniformly dispersed YAP within intracellular plasma. With further elevation of the electric field strength to 50 kV/cm, this tendency was partly reversed, meaning that the YAP moved closer to the nucleus once again. However, the formation of the YAP-TAZ complex was significantly disrupted at 50 kV/cm level.
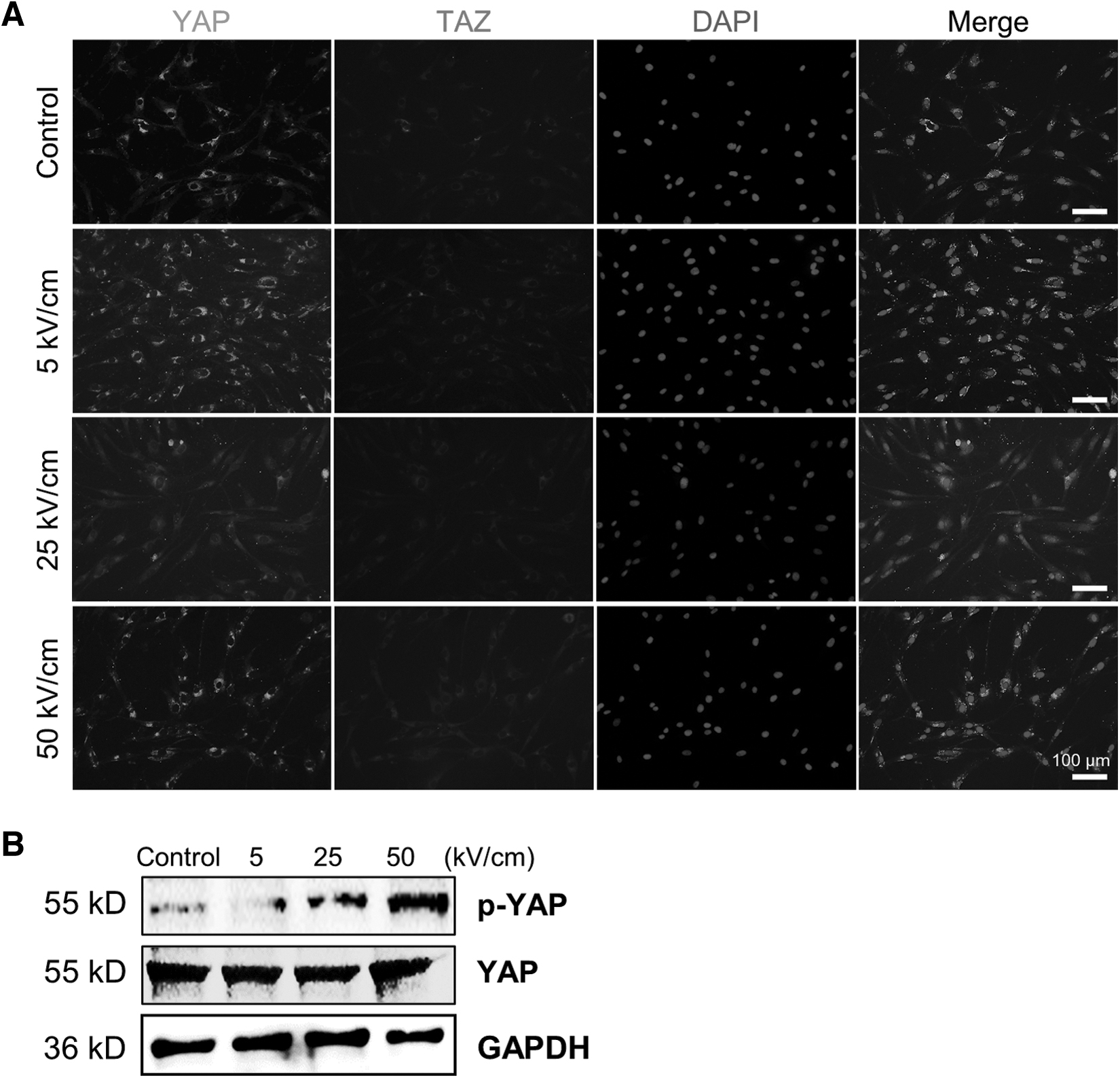
Low-intensity nsPEF alters YAP phosphorylation levels and its subcellular distribution.
Western blot suggested that YAP undergoes dephosphorylation upon low-intensity nsPEF treatment, and then its phosphorylation is activated by nsPEF in a dose-dependent manner as the electric field intensity is elevated (Fig. 6B).
Discussion
Reprogramming cellular phenotypes by exploiting adult cells' intrinsic transdifferentiation potential is currently a trending topic in regenerative medicine (Takahashi et al., 2007). Human dermal fibroblasts are considered a superior option for bone regeneration due to their low acquisition cost and obvious osteogenic tendency in specific pathological environments (Chen et al., 2019), but so far this phenomenon of transdifferentiation is difficult to replicate in vitro. There is still a need for a safe and reproducible method to increase the totipotency of cultured fibroblasts in vitro to enhance transformation efficiency. In recent years, research has shown that the application of low-intensity nsPEF can increase cell viability and enhance cell pluripotency in vitro (Vadlamani et al., 2019). However, even though nsPEF presents a promising opportunity to enhance the efficiency of in vitro osteogenesis induction in fibroblasts, there is a lack of relevant literature evidence, and suitable electric field parameters need to be investigated.
In previous cytobiological studies, the electric field strength of nsPEF in practical application scenarios is typically selected to be between 10 and 20 kV/cm (Zhou et al., 2023). Nevertheless, lower electric field strengths may be more beneficial for cell differentiation. The low-intensity nsPEF has no inhibitory effect on mammalian cells and are cytocompatible in most cases (Beebe, 2013; Das et al., 2020; Zhou et al., 2018) and is also widely used in the field of bioengineering synthesis, which can improve the biological conversion efficiency of single-celled organisms such as microalgae (Buchmann et al., 2019; Haberkorn et al., 2021; Haberkorn et al., 2019). Previous research has demonstrated that applying nsPEF to myoblasts or osteoblasts at an electric field intensity of 5 kV/cm can induce proliferation with consequent myotube maturation or the formation of osteogenic nodules. However, higher intensities of nsPEF (25 kV/cm) may hinder myoblast differentiation (Vadlamani et al., 2019). To elucidate the biological effects resulting from various intensity levels of nsPEF treatment, fibroblasts were subjected to low (5 kV/cm), high (25 kV/cm), and very high (50 kV/cm) intensities of nsPEF.
Following proliferation and migration assays, it was observed that low-intensity nsPEF treatment stimulated cell activity as previously reported. Cellular totipotency and osteogenic transformation efficiency were also significantly enhanced at the 5 kV/cm level. These results validate the previously proposed hypothesis that low-intensity nsPEF improves the osteogenic differentiation efficiency of fibroblasts by enhancing their transdifferentiation potential.
The biological effects of nsPEF are mechanistically linked to its alteration of phosphatidylinositol-4,5-bisphosphate (PIP2) enrichment on the cell membrane (Tolstykh et al., 2017b). As previously reported, treatment with nsPEF results in a dose-dependent deficiency of PIP2 on the surface of the cell plasma membrane (Tolstykh et al., 2017a), and consequently cannot activate Hippo pathway through the enrichment of the NF2 protein (Hong et al., 2020). The inactivation of the Hippo pathway will facilitate the nuclear translocation of nonphosphorylated YAP protein, thereby amplifying downstream transcriptional activity (Ma et al., 2019). However, extremely high electric field strength has a notable negative effect on cellular function; organelle damage triggers apoptosis and impaired protein synthesis, which is disadvantageous for clinical applications. Therefore, it is crucial to identify appropriate electric field parameters to improve the effectiveness of cell biotransformation.
In this study, nsPEF stimulation at 5 kV/cm significantly reduced YAP phosphorylation levels and led to a redistribution nearer to the cell nucleus, indicating that electric field stimulation of this magnitude may exist within the optimal dosage range and have a beneficial regulatory function. Interestingly, as the nsPEF intensity was increased to 50 kV/cm, nuclear translocation of YAP moderately increased, corresponding to the moderately regulated expression levels of various genes related to osteogenesis. It is speculated that this phenomenon may be closely linked to the initiation of the autophagy mechanism following organelle damage induced by high voltage (Ullery et al., 2015). Since the YAP protein homeostasis is closely related to cellular autophagy (Tang and Christofori, 2020), it cannot be degraded in time and thus remains around the nucleus, resulting in partial rescue of cellular transcriptional activity.
Recently DNMT1 has been identified as a new downstream effector of YAP/TAZ complex (Wu et al., 2017) and plays an important role in the determination of cell phenotypic transformation and differentiation fate (Hu et al., 2022). As reported in the previous literature, DNMT1 drives the reprogramming process in a variety of adult cells (Jin et al., 2020; Lee et al., 2001). Therefore, subsequent in-depth molecular mechanism studies could focus on the role of DNMT1 in the reprogramming process of fibroblasts. Although nsPEF was first reported to enhance totipotency by regulating DNMT1 activity in BMSC (Li et al., 2020), the effect of nsPEF treatment on fibroblast DNMT1 dynamics is uncertain. In the present study, the expression level of DNMT1 in fibroblasts was monitored to elucidate its role in cell transdifferentiation. In contrast to BMSCs, fibroblasts exhibited a different expression pattern of DNMT1 following low-intensity nsPEF stimulation. In the current study, it was found that the expression of DNMT1 by fibroblasts did not create a negative feedback loop with OCT-4 and Nanog during normal culture conditions.
After 14 days of osteogenic induction, low-intensity nsPEF stimulation resulted in a substantial decrease in allosteric-related genes while DNMT1 expression was further elevated, corresponding to significantly higher osteogenic markers than the control and other high-energy electric field stimulation groups. This implies that the regulation mechanism of transdifferentiation potential in adult cells may vary from that of stem cells.
In this initial study, it was preliminarily validated through in vitro experiments that low-intensity nsPEF stimulation can enhance the osteogenic transformation of human dermal fibroblasts in an osteoinductive culture environment. Although this phenomenon has not been validated histologically or in the in vivo microenvironment, the physical parameters identified in the preliminary test can guide subsequent investigations. The creation of this methodological model is advantageous for researchers seeking to delve deeper into the molecular mechanisms responsible for the impact of low-intensity nsPEF on cellular function and phenotypic evolution. In future study, low-intensity nsPEF-treated fibroblasts can be directly combined with an osteoinductive scaffold to fabricate engineered bone regeneration grafts with various construction measures. This augmentation strategy might also be used for the regeneration of diverse mesenchymal tissues, such as cartilage and myocardium. These approaches have the potential to improve bone defect repair based on tissue engineering principles and may also broaden the scope of clinical translational applications.
Footnotes
Authors' Contributions
J.L.: Conceptualization, Data Curation, Formal Analysis, Methodology, Resources, Investigation, and Writing—Original Draft. Z.W.: Conceptualization, Methodology, Investigation, and Writing—Original Draft. H.Z.: Formal analysis, Data Curation, and Visualization. P.L.: Validation, and Visualization. H.L.: Funding acquisition, Resources, Supervision, and Writing—Review and Editing. T.T.: Conceptualization, Funding acquisition, Project administration, Supervision, Validation, Visualization, and Writing—Review and Editing.
Consent Statement/Ethics Approval
The protocol for handling the human adipose tissue has been approved by the Clinical Research Ethics Committee of the First Affiliated Hospital, Zhejiang University School of Medicine.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This work was sponsored by the Zhejiang Provincial Natural Science Foundation (LS21H060001) and Young Talents Training Program of the First Affiliated Hospital of Zhejiang University School of Medicine (B22150).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.