
Abstract
Handmade Cloning (HMC) is a pivotal technique for cloning pig embryos. Despite its significance, the low efficiency of this method hampers its widespread application. Although numerous factors and signaling pathways influencing embryo development have been studied, the mechanisms underlying low developmental capacity and insufficient reprogramming of cloned embryos remain elusive. In the present study, we sought to elucidate key regulatory factors involved in the development of pig HMC embryos by comparing and analyzing the gene expression profiles of HMC embryos with those of naturally fertilized (NF) embryos at the 4-cell, 8-cell, and 16-cell stages. The results showed that ZFP42 expression is markedly higher in NF embryos than in cloned counterparts. Subsequent experiments involving the injection of ZFP42 messenger RNA (mRNA) into HMC embryos showed that ZFP42 could enhance the blastocyst formation rate, upregulate pluripotent genes and metabolic pathways. This highlights the potential of ZFP42 as a critical factor in improving the development of pig HMC embryos.
Introduction
Somatic cell nuclear transfer (SCNT) is a sophisticated technique to reconstruct a new embryo by transplanting the nucleus from a somatic cell into an enucleated oocyte (Polejaeva et al., 2000). SCNT has been extensively applied in many fields including agricultural and biomedical research (Aigner et al., 2010; Lai et al., 2002; Rahbaran et al., 2021). Nevertheless, low efficiency poses a significant challenge to its further application.
The mechanisms governing the conversion of a fertilized oocyte into an embryo are heavily influenced by the nuclear remodeling and reprogramming of donor nuclei (de Macedo et al., 2022; Malpotra et al., 2022). Epigenetic alterations, such as histone acetylation and DNA methylation, are considered to be pivotal in orchestrating cell reprogramming (Jeong et al., 2021; Jullien et al., 2017; Papp and Plath, 2013).
Through the regulation of these epigenetic modifications, the expression of over 10,000 genes is modified at fertilization to promote the progress of embryonic and fetal development (Matoba and Zhang, 2018; Niemann, 2016; Ripamonte et al., 2012; Zhao et al., 2022). Transcriptional activities in SCNT embryos display variation across species, as observed in mice (Liu et al., 2016), humans (Chung et al., 2015), cattle (Liu et al., 2018), and pigs (Bai et al., 2016; Liu et al., 2020).
However, SCNT embryos of different species have the same defects that many genes did not activate as naturally fertilized (NF) embryos. It has been reported that approximately a thousand genomic regions or genes did not activate during the zygotic genome activation (ZGA) stage in mouse SCNT embryos (Matoba et al., 2014). Therefore, identifying the key factors influencing epigenetic modification of SCNT embryos is essential for elevating the efficiency of the technique.
Notably, recent studies have indicated that the H3K9me3 demethylase family, including Kdm4a, Kdm4b, Kdm4d, and Kdm4e, plays a key role in global H3K9me3 demethylation, which is a common impediment in mammalian SCNT embryo reprogramming (Liu et al., 2016; Malpotra, 2022; Matoba and Zhang, 2018; Zhao et al., 2022).
Pigs are not only a vital source of meat consumption but also an exceptional model for biomedical research due to their similar anatomical and physiological resemblance to humans (Lai et al., 2002; Nguyen et al., 2023). As an alternative to the traditional SCNT technique, Handmade Cloning (HMC) has been applied to clone pig in a few laboratories for its low cost, easy operation, and higher efficiency than the traditional technique (Du et al., 2007; Liu et al., 2020). However, its efficiency is still low due to the fact that the mechanisms underlying developmental barrier in pig cloned embryos remain inadequately understood.
In our study, we conducted RNA-sequencing (RNA-seq) to profile gene expression in pig HMC embryos and natural fertilized (NF) embryos at 4-cell, 8-cell, and 16-cell stages. Our findings indicated that ZFP42 expression was significantly elevated in NF embryos compared with their cloned counterparts. Introduction of exogenous ZFP42 messenger RNA (mRNA) could, to some extent, remove the reprogramming barriers of pig HMC embryos.
Materials and Methods
In vitro maturation of oocytes and injection of ZFP42 mRNA
Cumulus-oocyte complexes (COCs) were harvested from pig ovaries obtained from a slaughterhouse and washed thrice with pre-warmed phosphate-buffered saline (PBS) at 37°C. The COCs were then cultured in medium containing 80% tissue culture medium 199 (TCM-199), 10% pig follicular fluid, 10% fetal bovine serum (FBS), 10 IU/mL equine chorionic gonadotropin (eCG), and 5 IU/mL human chorionic gonadotropin (hCG; Suigonan Intervet, Skovlunde, Denmark).
The culture dish was maintained at 38.5°C in a humidified atmosphere with 5% CO2 for oocyte maturation. After 42–46 hours, the COCs were subjected to enzymatic digestion and washed as described previously (Liu et al., 2020). Briefly, COCs were exposed to T33 (T for Hepes-buffered TCM-199 medium; the number means percentage [v:v] of calf serum supplement, here 33%) medium containing 3.3 mg/mL pronase for 25 seconds to remove the zonae pellucidae. The liberated oocytes were washed twice in T10 and T20 media and then aligned in T10 drops containing 3 μg/mL cytochalasin B.
Pig ZFP42 and GFP complementary DNA (cDNA) synthesized by TsingKe Ltd. were cloned into the pUC57 vector separately. After amplification, the vectors were linearized with the restriction enzymes BamHI-HF and KpnI-HF to produce capped pig ZFP42 and GFP mRNA according to the manufacturer's instructions (AM1340; Invitrogen).
The mRNA molecules with 3′ ends polyadenylated (C-PAP5104H; Cellscript) were stored at −80°C, and diluted to a concentration of 100 μg/mL in PBS. After a few times test, the opposite direction of the polar body in oocytes was identified the best site for the injection of PBS containing either GFP mRNA or ZFP42 mRNA (about 20 pL aliquot), for it could avoid the loss of injected mRNA in the next step. The polar body was removed with approximately one-third of the adjacent cytoplasm by using Ultra Sharp Splitting Blades (AB Technology, Pullman, WA) and the resultant cytoplasts were collected.
Production of pig HMC embryos
Donor cell line was established and cultured as previously described (Liu et al., 2015). Primary cells were isolated from 40-day-old pig fetuses and cultured in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% FBS, 1% glutamine (Invitrogen), and 1% nonessential amino acids at 37°C in an atmosphere. The cell lines were passaged three times before being used as donor cells.
The cell fusion procedure was performed in two steps. Initially, donor cells were allocated evenly in a 20 μL T2 drop. Polar body was firstly removed with about one-third of its nearby cytoplasm using Ultra Sharp Splitting Blades (AB Technology). The putative cytoplasts were first incubated in T0 medium containing 1 mg/mL phytohemagglutinin (PHA; ICN Pharmaceuticals) for 5 seconds, then swiftly transferred to a T2 drop to adhere to a single donor cell.
The cytoplast-donor cell pairs were subjected to an electric pulse of 0.6 kV/cm and 700 kHz for 9 μseconds to induce fusion (BLS CF-150/B, Budapest, Hungary), followed by incubation in T10 drops. After 1 hour, the fused cytoplast-donor cell pairs and an additional putative cytoplast were subjected to a second electric pulse of 0.85 kV/cm for 80 μseconds. The reconstructed embryos were cultured in PZM-3 medium at 38.5°C in an atmosphere of 5% CO2, 5% O2, and 90% N2 (Vajta et al., 2000).
Evaluation of embryo development
After 2 or 6 days culture, the GFP expression of embryos was carefully examined under the SZX7 fluorescence microscope (Olympus, Tokyo, Japan). To assess the efficiency of the embryonic development process, embryos were observed under a microscope to calculate the final blastocyst rate. There were three replicates in each group. Analysis of variance analysis was carried out using SPSS12.0. A value of p < 0.05 was supposed to be statistically significant.
RNA-seq and data analysis
The pig oocytes or embryos were frozen immediately in dry ice after being taken out from incubator and sent to BGI (Shenzhen, China) for RNA-seq by DNBSEQ platform. The raw data were processed to obtain clean reads by SOAPnuke (self-developed by BGI, v1.5.6). High-quality clean reads were mapped with reference gene sequences by Bowtie2 (v2.3.4.3).
Gene expression levels were calculated by RSEM (v1.3.1). Gene function annotation and functional associations were predicted by using Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG). The p-value was calculated using R software (DESeq2). GO and KEGG analyses were performed using the R software (Yu et al., 2012).
Results
RNA-seq both NF embryos and HMC embryos in different stages
In this study, pig fetal fibroblasts served as donor cells for the construction of HMC embryos. We collected cloned embryos and NF embryos at various developmental stages, including 2-cell (absent in the NF group), 4-cell, 8-cell, 16-cell, and blastocyst stages for RNA-seq analysis (Fig. 1A). The raw data generated for NF embryos at the 4-cell, 8-cell, 16-cell, and blastocyst stages amounted to 62,824,411, 86,536,191, 82,134,172, and 62,330,617 reads, respectively.
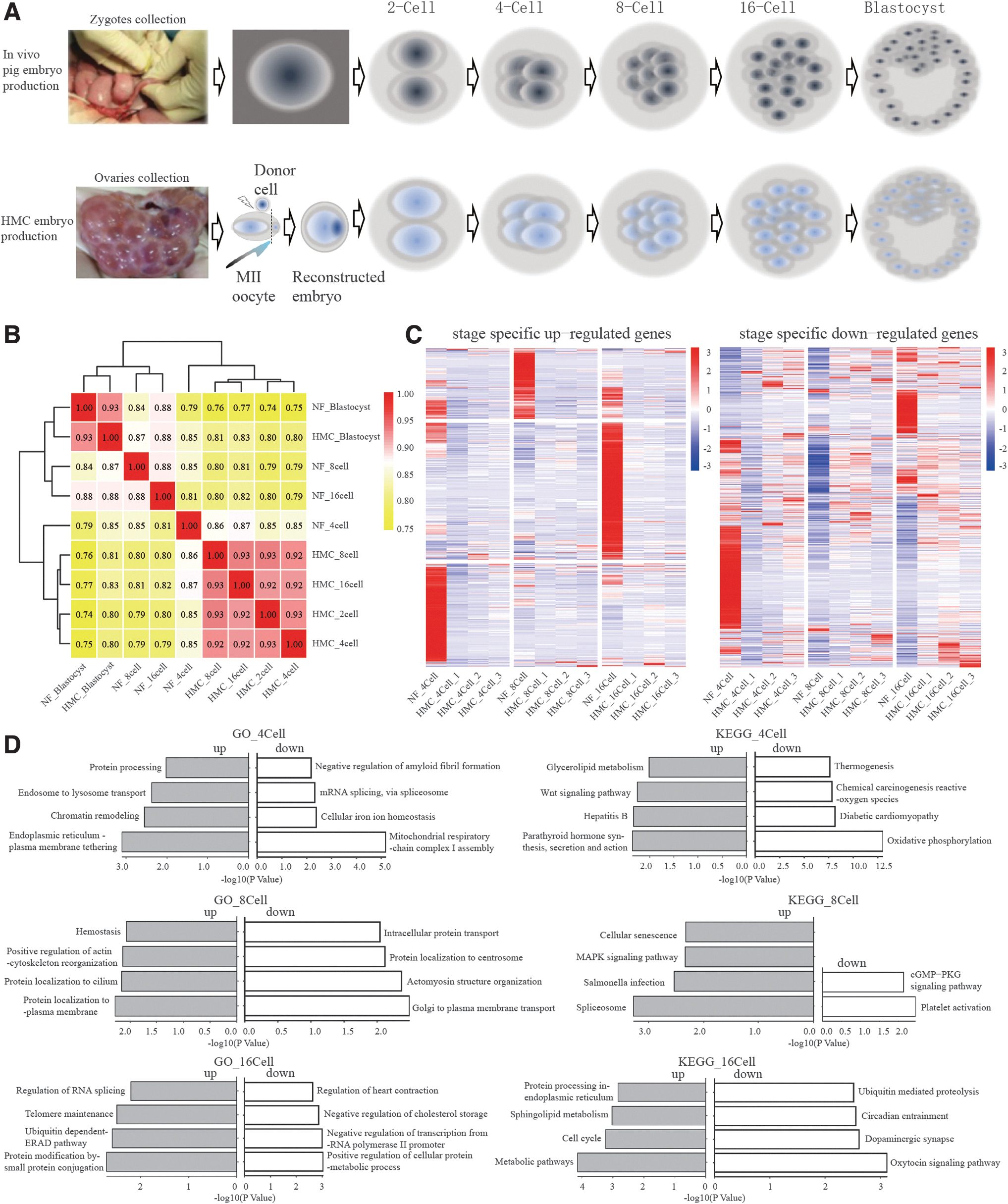
Specific genes expression pattern and pathways involved in the development of pig HMC embryos.
The clean data accounted for 99.96% of the total reads, with a mapping rate to the pig genome reference (GCF_000003025.6_Sscrofa11.1) ranging from 58.70% to 85.57%. For the HMC embryos, the raw data obtained for the 2-cell, 4-cell, 8-cell, 16-cell, and blastocyst stages were 60,522,421, 62,301,113, 58,341,497, 45,332,575, and 58,377,909, respectively (Table 1). The proportions of clean data and mapping rates were consistently higher than 99% and 75%, respectively, indicating the high quality of our data for further analysis.
RNA-Sequencing Data of Handmade Cloning and Natural Fertilized Embryos for Correlation Analysis
HMC, handmade cloning; NF, natural fertilized.
Heatmap analysis revealed that the gene expression profile of blastocysts, in both HMC and NF groups, was distinctly different from the earlier stages (4-cell, 8-cell, 16-cell). The correlation coefficients before the blastocyst stage in the NF group varied widely, ranging from 0.81 to 0.88 (Fig. 1B). Interestingly, the correlation coefficient between any two samples at different stages (2-cell, 4-cell, 8-cell, or even 16-cell) within the HMC group was consistently over 0.92.
This high correlation coefficient might suggest that certain robust genes expressed in the early development of NF embryos are absent in cloned embryos, potentially contributing to the lower blastocyst formation rate observed in cloned embryos compared to NF embryos.
To identify candidate genes that may help HMC embryos overcome the reprogramming barriers, we further collected two more replicated samples of HMC embryos and conducted comparative transcriptome analyses of the sample of NF embryos with all three replicated samples of HMC embryos from the 4-cell to 16-cell stages (Fig. 1C and Supplementary Table S1).
GO analysis of the differentially expressed genes (DEGs) indicated that upregulated genes in NF embryos were specifically enriched in protein processing and chromatin remodeling at the 4-cell stage, protein localization at the 8-cell stage, and protein modification and regulation of RNA splicing at the 16-cell stage. However, the downregulated genes were enriched in oxidation, protein metabolic process, and others. It should be noted that upregulated genes were associated with protein localization to cilium or plasma membrane, whereas the downregulated genes were associated with protein localization to the centrosome (Fig. 1D and Supplementary Table S1).
Dynamic gene regulation analysis in the development of pig HMC embryos
The early development of the embryo is involved in the consuming of maternal mRNA and transcription of zygotic genome after activation (Fig. 2A). The process is regulated by genes related to epigenetic modification, including methylation and deacetylation. To identify the potential candidates, we further analyzed the expression pattern of those genes related to epigenetic modification in both HMC embryos and NF embryos at different stages, including 2-cell, 4-cell, 8-cell, 16-cell, and blastocyst (Donor cell was calculating as a control).
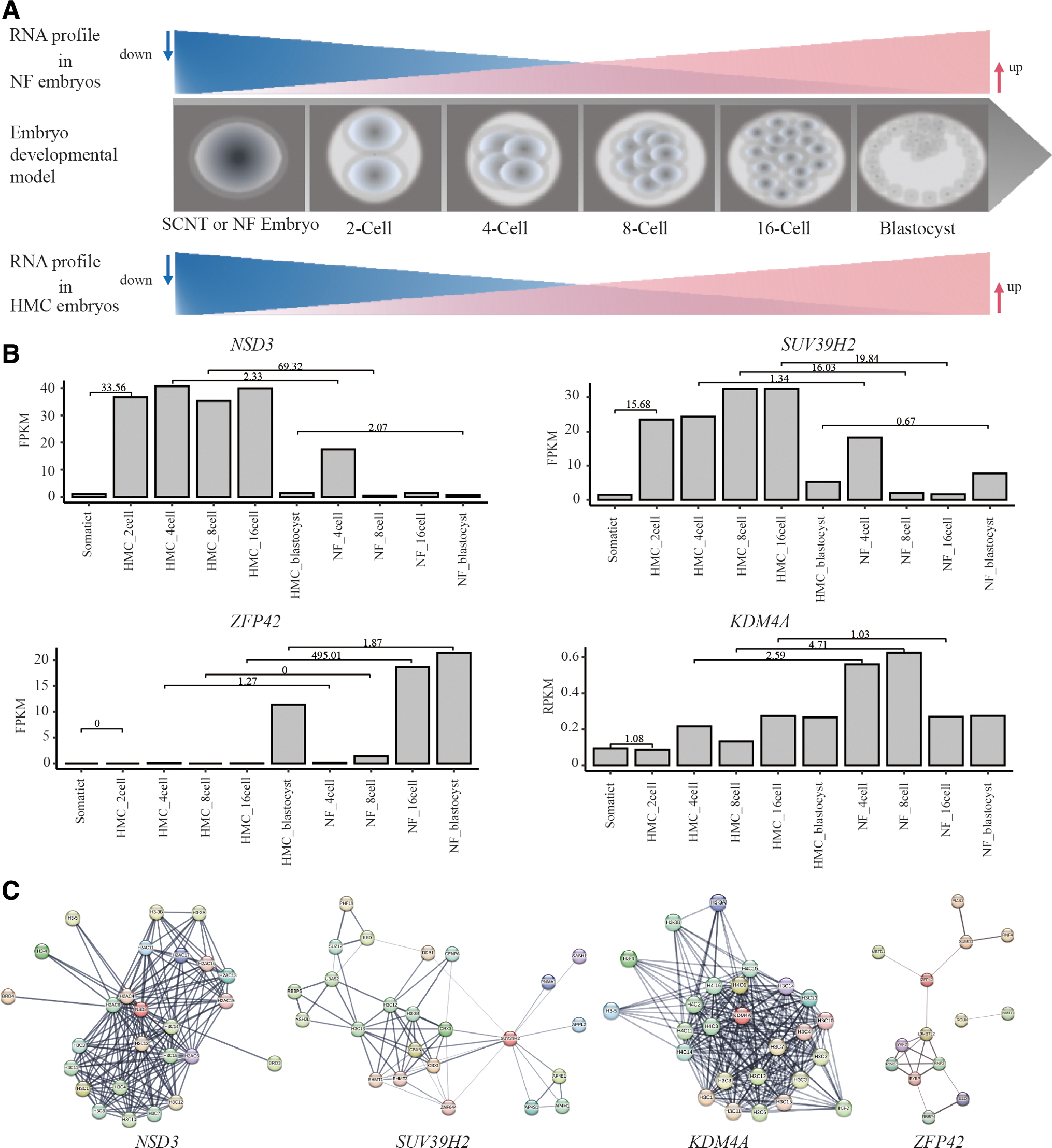
Prediction of targeted genes and their function network.
The results showed that four genes, NSD3, SUV39H2, ZFP42, and KDM4A have interesting expression patterns. NSD3 and SUV39H2 exhibited parallel expression trajectories, increasing to high expression in both 4-cell stage HMC and NF groups (Fig. 2B). However, the expression of these two genes swiftly declined in 8-cell stage NF embryos, whereas it was maintained high in HMC embryos until the blastocyst stage.
Conversely, ZFP42 displayed an inverse expression pattern compared with NSD3 and SUV39H2, with high expression in both HMC blastocysts and NF blastocysts. However, there was a delayed upregulation of ZFP42 in NF embryos until the blastocyst stage, compared with HMC embryos upregulated from the 16-cell stage (Fig. 2B). KDM4A maintained a consistently low expression level from the 2-cell stage to the blastocyst stage in HMC embryos (Fig. 2B).
In contrast, in NF embryos, KDM4A expression was elevated at the 4-cell and 8-cell stages but diminished at the 16-cell and blastocyst stages. This pattern suggested that KDM4A might play a transient regulatory role in early embryo development.
Further bioinformatics analysis of these four genes indicated their extensive involvement in the epigenetic modification signaling pathways associated with cellular developmental pluripotency. For instance, ZFP42 is predicted to directly interact with DNMT3L, which is known to stimulate de novo DNA methylation, and with DPPA3, which is implicated in the facilitation of embryonic stem cell self-renewal (Fig. 2C).
These interactions and pathways may be crucial for understanding the molecular mechanisms governing early embryonic development and the reprogramming barriers encountered in HMC embryos.
Investigating the impact of ZFP42 overexpression on early development of pig HMC embryos
Overexpressing gene as a methodological approach has proven to be a powerful tool for studying the biological function of candidate genes in targeted cell lines. For instance, the overexpression of ZFP57 in bovine embryos has been demonstrated to enhance the development of SCNT blastocysts (Yu et al., 2023). Based on the report and the result described earlier, we sought to determine whether overexpressing ZFP42 could have a similar effect on the development of pig HMC embryos.
To validate the functionality of our mRNA overexpression approach within the HMC framework, we established an mRNA overexpression system during HMC procedure (Fig. 3A). First, the system was optimized and validated by microinjecting GFP mRNA. The green fluorescence observed in both 2-cell stage and blastocyst stage HMC embryos confirmed that exogenous mRNA could be consistently translated into protein (Fig. 3B, C). We then cloned ZFP42 cDNA and constructed a gene overexpression system using the pUC57 plasmid. Subsequently, ZFP42 mRNA was successfully produced with a kit (Fig. 4A, B).
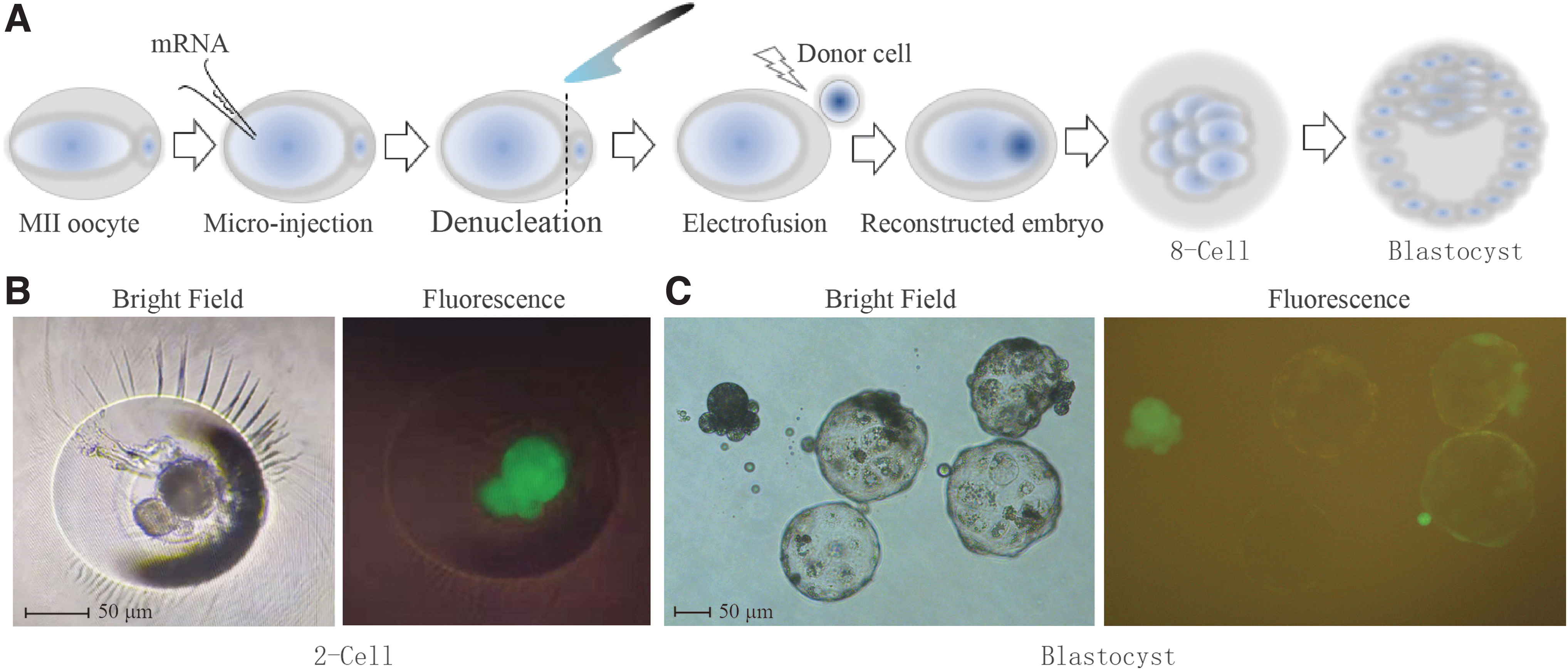
Optimization of microinjection system.
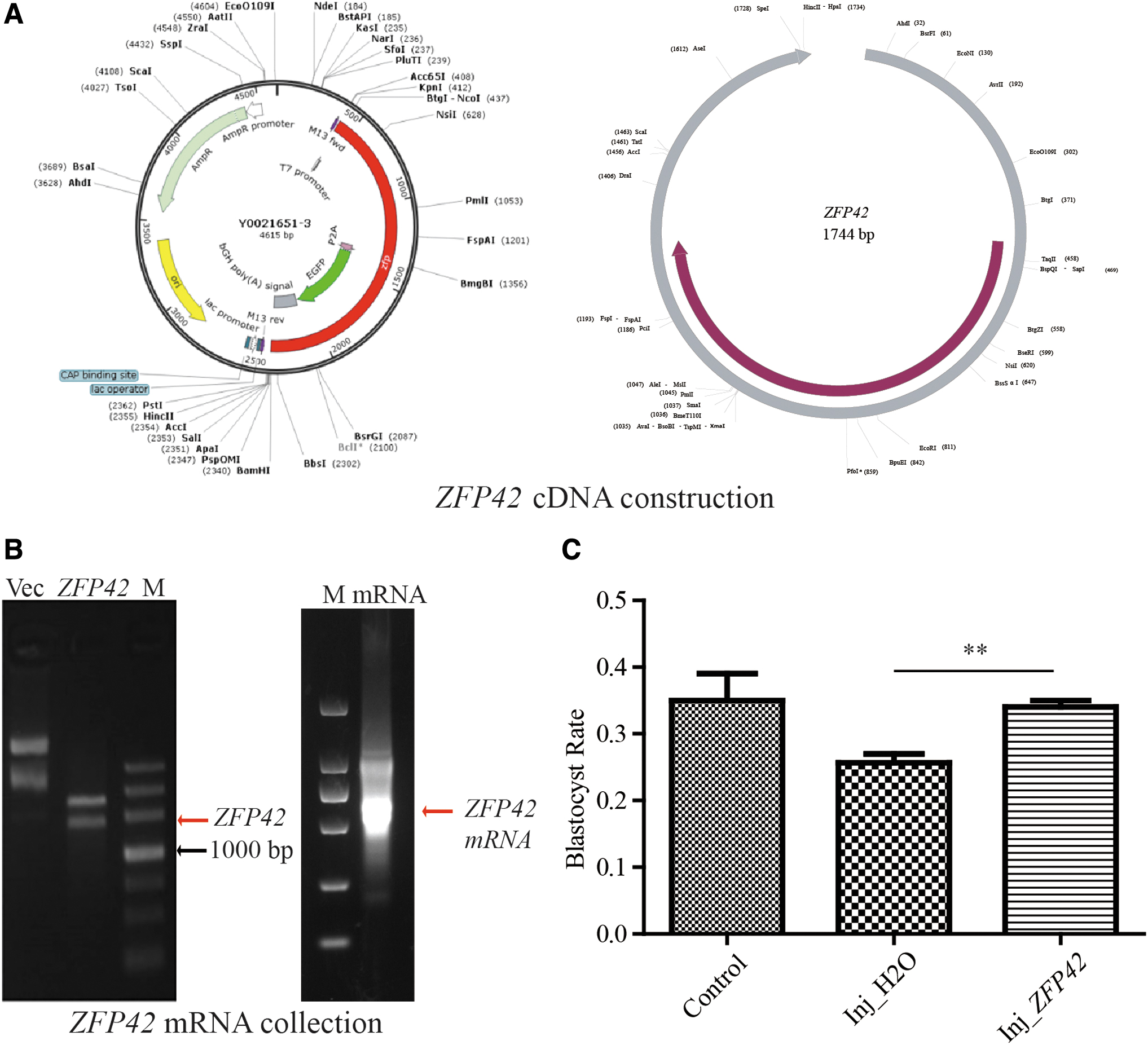
ZFP42 overexpression promotes pig HMC embryonic development.
After that, we produced HMC embryos with three of different procedures as follows: injecting ZFP42 mRNA (Inj_ZFP42 group) or H2O (Inj_H2O group) into oocytes before enucleation, standard treatment (Control) (Table 2). Seven days post-injection, we assessed the developmental status of the embryos under a microscope, and the blastocyst rate was calculated (Table 2 and Supplementary Fig. S1) to evaluate the functional contribution of ZFP42.
Embryos Used in Investigating Effect of ZFP42 on Handmade Cloning Embryo Development
Inj_ZFP42: embryos injected with ZFP42 mRNA; Inj_H2O: embryos injected with H2O; Control: HMC embryos produced with standard process.
mRNA, messenger RNA.
The blank control group exhibited a mean blastocyst rate of 34.78%, which was comparable with the results reported in previous studies. However, the blastocyst formation rate of Inj_H2O embryos was decreased to 26.47%, which could be attributed to the potentially detrimental impact of the microinjection process. Nevertheless, the introduction of ZFP42 mRNA resulted in a significant improvement in the HMC blastocyst rate when compared with the Inj_H2O group (34.15% vs. 26.47%, respectively; p < 0.01, Fig. 4C).
These findings imply that the overexpression of ZFP42 can effectively enhance the early development of HMC embryos, suggesting a positive role for ZFP42 in the embryonic development of pig HMC embryos.
Exploring the role of ZFP42 in pig HMC embryo development through RNA-seq analysis
To investigate the potential gene networks that facilitate the overcoming of reprogramming barriers in pig HMC embryos, we conducted RNA-seq on 8-cell stage HMC embryos (Supplementary Table S2). The data revealed that ZFP42 mRNA levels were significantly elevated in the 8-cell stage Inj_ZFP42 embryos compared with the other two groups (Fig. 5A). A substantial number of DEGs, including pluripotent genes LMCD1, RHOC, PTPRK, LEF1, and DLX2, were all significantly higher (p < 0.01) in Inj_ZFP42 HMC embryos than the Inj_H2O group (Fig. 5B).

Possible mechanism of ZFP42 on the development of pig HMC embryos.
GO and KEGG enrichment analysis were conducted to annotate the functions and biological processes associated with these DEGs. Notably, terms related to “metabolic pathways” “focal adhesion assembly,” and pathway related to cell migration were enriched in the Inj_ZFP42 embryos compared with the control embryos (Fig. 5C; Supplementary Table S3). Similar pathways and the sarcomere organization were also observed when comparing the Inj_ZFP42 group with the Inj_H2O group (Fig. 5D; Supplementary Table S3).
These results were compatible with earlier blastocyst rate data (Fig. 4C), suggesting that enhanced cell metabolism, sarcomere organization, and migration could enhance developmental capability of embryo and result in higher blastocyst formation rates.
These findings indicated that ZFP42 overexpression may promote cellular differentiation and development of pig HMC embryos by influencing pluripotent processing pathways. The upregulation of genes associated with pluripotency and proliferation further suggested that ZFP42 could play a crucial role in enhancing developmental potential of early-stage pig HMC embryos.
Discussion
In the past six decades, over 20 species have been cloned by SCNT. HMC as an alternative to tranditional SCNT technique has advantages of low cost and easy operation, and more and more labs applied HMC to clone pig for pig models have great application potential in agriculture and biomedical research (Du et al., 2007; Liu et al., 2020; Polejaeva et al., 2000; Yin et al., 2002). However, despite a few advances, the underlying mechanism leading to developmental arrest of pig embryos remain largely unknown. In the present study, we charted the gene expression profiles across various embryonic stages (2-cell, 4-cell, 8-cell, 16-cell, and blastocyst), and found that the histone methyltransferases NSD3 and SUV39H2 were upregulated in HMC embryos during pre-blastocyst stages compared with NF embryos.
NSD3 is one member of the NSD family involved in the demethylation of histone H3 at lysine 36 (H3K36). It has been reported that NSD3 could result in dysregulated modification of H3K36 in various cancers (Lawrence et al., 2015; Rathert, 2021). There were also studies showing that the loss of H3K36me2 by knocking out NSD1 in embryonic stem cells could cause significant epigenetic reorganization of cells (Chen et al., 2022; Li et al., 2021).
The results of those studies lead us to assume that H3K36me loss in HMC embryos by dysregulating NSD3 may disrupt embryo division via as-yet-unknown molecular pathways.
The trimethylation of histone H3 at lysine 9 (H3K9me3) has been identified as a critical epigenetic barrier to the reprogramming of SCNT embryo in different species (Chung et al., 2015; Jafarpour et al., 2020; Matoba et al., 2014). A previous study showed that inhibiting an enzyme responsible for H3K9me3, SUV39H2 could enhance SCNT embryo development (Jafarpour et al., 2020).
This is in accordance with our observation that higher SUV39H2 expression correlates with lower blastocyst formation rates of HMC embryos. Conversely, KDM4A, a demethylase of H3K9me3, displayed an inverse expression pattern to SUV39H2 in the present study. It has been reported that reprogramming barriers resulted from H3K9me3 could be removed by KDM4A during the early development of mice and humans' zygote (Chung et al., 2015; Sankar et al., 2020), and KDM4A overexpression can significantly improve blastocyst formation rate (Chung et al., 2015). Those reports implied that low expression of KDM4A in pig HMC embryos could be one of the primary factors contributing to low blastocyst rates.
ZFP42 is an embryonic stem cell marker encoding a protein that comprises four Cys-His type zinc-fingers and plays a role in DNA binding and transcriptional regulation (Pérez-Palacios et al., 2021; Rogers et al., 1991). Previous studies showed that ZFP42 mRNA expression was upregulated following zygotic transcription initiation, with levels in both mouse and pig embryos gradually increasing from the 8-cell stage to the blastocyst stage (Climent et al., 2013; Mohana Kumar et al., 2012).
In the present study, we also observed similar results that ZFP42 mRNA in pig blastocyst-stage embryos was more abundant than in embryos at earlier stages. Interestingly, ZFP42 mRNA was nearly undetectable in HMC embryos from the 2-cell to 16-cell stages.
We observed that overexpressing ZFP42 via mRNA injection into mature pig oocytes could significantly increase the blastocyst formation rate of HMC embryos from 26.47% (the Inj_H2O group) to 34.15%. Subsequent RNA profile analysis supported this point. Compared with Inj_H2O embryos at the 8-cell stage, lots of genes associated with pluripotency such as LEF1, LMCD1, RHOC, DLX4, and PTPRK were greatly increased in Inj_ZFP42 embryos.
However, most of their expression differences were moderate between Inj_H2O and Inj_ZFP42, which was an equal match with their blastocyst rate. Another important point is that overexpression of ZFP42 might reduce the influence of mechanical operations by enhancing embryo metabolism. As we interestingly observed that pathways related to sarcomere organization and metabolism such as “endocytosis” and “metabolic pathways” were enriched in the Inj_ZFP42 cells. Besides, we also found terms related to “positive regulation of gene expression” and pathways related to cell migration were enriched in Inj_ZFP42 embryos when compared with the Inj_H2O group.
This study sheds light on the distinct transcription profiles of pig HMC embryos and highlights the dysregulation of epigenetic and transcriptional regulation genes, including NSD3, SUV39H2, ZFP42, and KDM4A in HMC embryos from 2-cell to blastocyst stages. Further experiments confirmed that overexpressing ZFP42 in matured pig oocytes used for HMC could reduce the reprogramming barrier to some extent and increase the blastocyst formation rate.
Footnotes
Acknowledgment
The authors are thankful to TsingKe Ltd. for helping ZFP42 and GFP vector construction.
Authors' Contributions
T.L. contributed data analysis. Y.W. performed a part of the work in partial fulfillment of his Master's thesis. L.L. and T.Z. performed experiments. X.Z. and Y.L. designed and wrote the article.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
The study is supported by GuangDong Basic and Applied Basic Research Foundation (No. 2019B1515210028) and the Science and Technology Innovation Strategy Projects of Guangdong Province (2019B020203002).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.