
Abstract
Cloning by somatic cell nuclear transfer (SCNT) remained challenging for Rhesus monkeys, mostly due to its low efficiency and neonatal death. Genome-scale analyses revealed that monkey SCNT embryos displayed widespread DNA methylation and transcriptional alterations, thus including loss of genomic imprinting that correlated with placental dysfunction. The transfer of inner cell masses (ICM) from cloned blastocysts into ICM-depleted fertilized embryos rescued placental insufficiency and gave rise to a cloned Rhesus monkey that reached adulthood without noticeable abnormalities.
Main Text
The somatic cell nuclear transfer (SCNT) technology allows the replacement of an oocyte genome with a lineage-committed cell nucleus (Moura, 2023; Solter, 2000). The enucleated oocyte has the ability to reprogram the donor cell by removing epigenetic marks and chromatin components associated with the differentiated cell state and thus providing the cellular context for the reacquisition of totipotency (Matoba and Zhang, 2018; Wilmut et al., 1997). The development of SCNT reprogramming offered numerous applications, which included basic research, conservation efforts, support to genetic engineering, and generation of genetically-tailored stem cells (Moura, 2023). For several species, the most prominent application of SCNT cloning was to make whole animals from genetically modified somatic cells (Polejaeva, 2021; Wolf et al., 1999).
There was much expectation for cloning non-human primates (NHP). Coupled with genetic engineering, NHP cloning could provide animal models of human disease, particularly for late-onset conditions with systemic complications. The attempts to clone NHP were far fewer than other mammals, due to financial, ethical, and logistical drawbacks. Based on previous work on mice (Matoba and Zhang, 2018), authors used epigenetic tweaks, namely the chemical inhibition of histone deacetylation (Kishigami et al., 2006; Rybouchkin et al., 2006) and exogenous supplementation of gene transcripts encoding a histone 3 lysine 9 demethylase (Matoba et al., 2014) to facilitate the process of epigenetic reprogramming (Fig. 1). This cloning approach ultimately led to the birth of cloned Cynomolgus monkeys (Macaca fascicularis) (Liu et al., 2019, 2018), and the two female clones are now over 6 years old and healthy. However, it remained unclear what could be the roadblocks to the cloning of other NHP widely used in biomedical research.
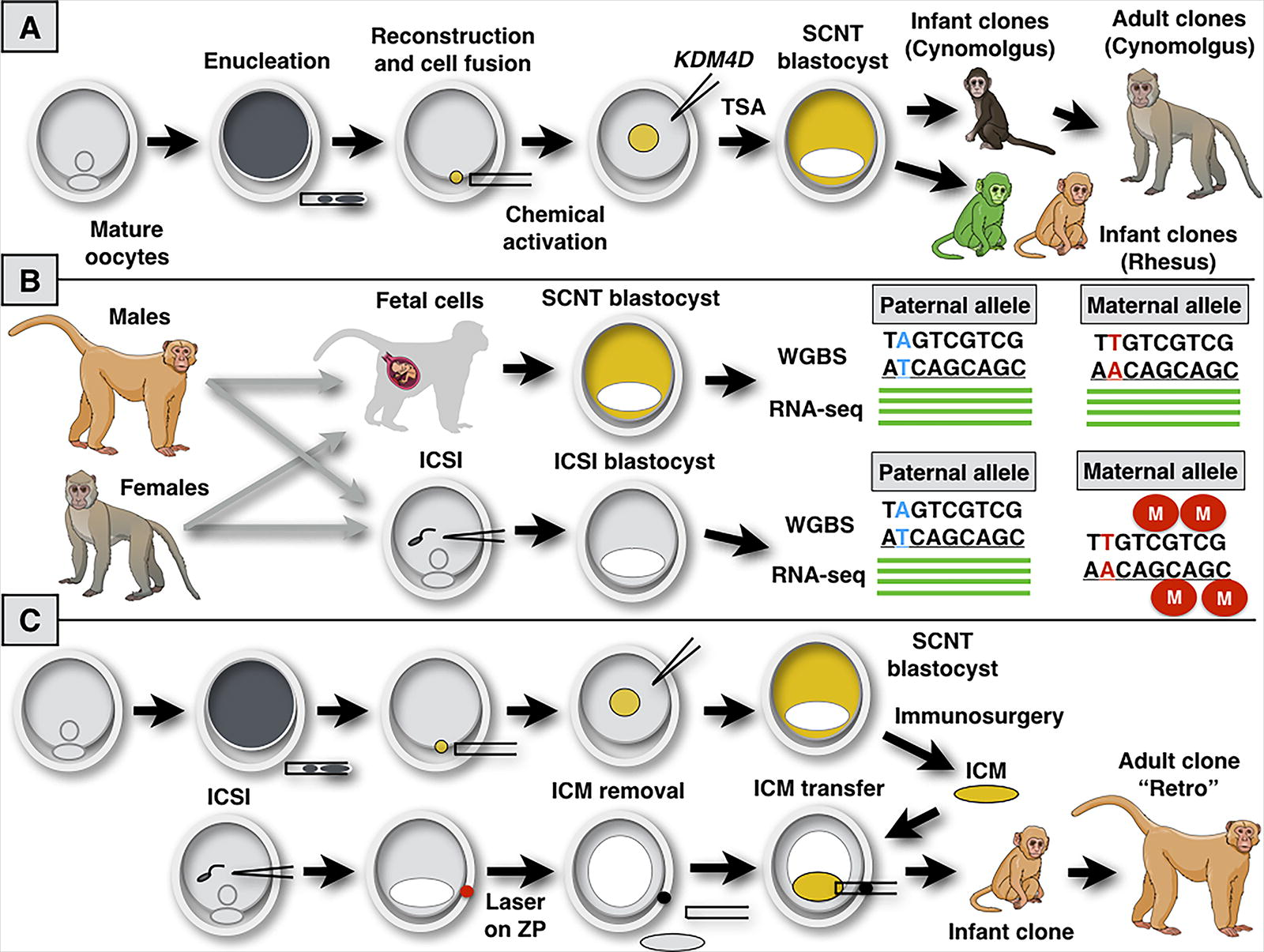
Improving the cloning protocol for Rhesus monkeys through biological insights generated by functional genomics.
The Rhesus monkey (Macaca mulatta) has been a prominent NHP model in the context of reproductive and therapeutic cloning (Byrne et al., 2007; Meng et al., 1997; Wolf et al., 1999). Previous work demonstrated the cloning of Rhesus monkeys from blastomeres (Meng et al., 1997) and the derivation of embryonic stem cell lines from SCNT blastocysts (SCNT-ESCs) (Byrne et al., 2007). More recently, cloned Rhesus monkeys were obtained from a 12-year-old transgenic male but died shortly after delivery (Kang et al., 2022). It was long clear that technical details of SCNT technology were not hindering full-term development of cloned Rhesus monkeys (Mitalipov and Wolf, 2006) but most likely the incomplete epigenetic reprogramming.
Liao and colleagues now shed light on a biological barrier to the cloning of Rhesus monkeys (Liao et al., 2024). Initial work found similar blastocyst rates between SCNT embryos and those derived from intracytoplasmic sperm injection (ICSI), albeit cloned embryos established fewer pregnancies (7.23% for SCNT vs. 14.82% for ICSI) that mostly succumbed around implantation. One full-term gestation give rise a cloned Rhesus monkey that survived for almost a day. Collectively, these results closely resembled the previous work on Rhesus SCNT cloning (Kang et al., 2022), thus suggesting some consistent biological barrier hindering full-term development and neonatal viability.
To explore the potential causes of such incomplete reprogramming, Liao and colleagues investigated the global DNA methylation landscape of blastocysts obtained by ICSI and SCNT. An unusual result was a global hypomethylation of the genome in SCNT embryos in contrast to ICSI controls, albeit hypermethylated regions in SCNT embryos were suggestive of somatic-cell-of-origin and overlapped gene bodies to quite some extent. Based on these results, authors turned their focus to imprinted genes. This gene subset was appealing due to their role in development (and placentation), allele-specific epigenetic control, and found aberrantly expressed in mammalian clones and their placentas, which included monkeys (Chu et al., 2021; Ogura et al., 2021). A transcriptomic analysis using bulk RNA-sequencing explored the transcriptional changes associated with epigenetic reprogramming. Both SCNT and ICSI embryos used for this analysis had hybrid genomes (Macaca fascicularis x Macaca mulatta) to facilitate allele-specific analyses. Therefore, authors mapped 115 imprinted genes in hybrid monkey blastocysts. A total of 420 gene transcripts were differently expressed between SCNT and ICSI blastocysts, thus reinforcing the notion that imprints are part of the molecular differences in such embryos.
Despite the potential of single “omics” (e.g., WGBS, RNA-seq), these approaches alone have limited capacity to capture the complexity of biological systems. The analytical integration of multiple “omics” datasets has greater resolution to pinpoint molecular processes of casual effect. Liao and colleagues took a candidate approach to merge a small sample of these datasets and find more appealing biological evidence of improper reprogramming. The selected candidate loci were a subset of imprinted genes that displayed both altered DNA methylation and transcriptional levels, which led to four aberrant imprinted genes (DNMT1, RHOBTB3, SIAH1, and THAP3). Of note, these losses of imprinting marks were also found in donor somatic cells, which rules out the cloning process from causing these molecular changes. Despite these interesting findings, authors could have used a SNP-free approach for mapping more imprinted genes (Chu et al., 2021).
Liao and colleagues went out to explore in vitro and in vivo post-implantation development to explore the epigenetic errors around implantation. In both experimental settings, post-implantation SCNT embryos had strikingly higher DNA methylation levels and loss-of-imprinting (i.e., THAP3, DNMT1, and SIAH1) in contrast to ICSI counterparts. There was also heterogeneity in these few samples (n = 3), such that some cloned placentas displayed normal (n = 2) or abnormal DNA methylation levels (n = 1). Additional analyses showed anatomical and physiological abnormalities in cloned placentas (thicker placentas, deposition of calcium salts, and calcified clouds). Hence, these findings alongside previous work (Kang et al., 2022) pointed out that placentation was a major issue hampering Rhesus cloning, and it overlapped with loss of imprinting.
In search for ways to improve placenta function in clones, Liao and colleagues explored the possibility of generating tetraploid recipient embryos, which are biased toward placenta differentiation, and have very limited contribution to the fetus (Li et al., 2005; Nagy et al., 1990). The injection of ICMs from SCNT embryos or SCNT-ESCs into tetraploid embryos (known as tetraploid embryo complementation assay) ameliorates placental insufficiency and allowed cloning from hard-to-clone cell types (Eggan et al., 2004; Hochedlinger and Jaenisch, 2002; Nagy et al., 1990). The fusion of blastomeres in 2-cell embryos led to the formation of healthy blastocysts. The transfer of these fused embryos to recipients led to pregnancies and births, thus unequivocally demonstrating that many embryos did not undergo polyploidization. The analysis of fertilized embryos harboring a H2B-EGFP reporter gene showed two modes of resistance to tetraploid nuclear formation, which displayed varying physical approximation of nuclei after fusion (coming close or remaining distant), and ultimately prevented the formation of bona fide tetraploid blastocysts.
To circumvent this issue, the authors used a laser bean to rupture the zona pellucida of ICSI blastocysts and subsequently remove the ICM by micromanipulation, without affecting the viability of the trophectoderm (Fig. 1). This process named trophectoderm replacement (TR) did not leave residual ICM cells, as demonstrated by cellular analysis of post-implantation embryos and lack of births after embryo transfer to recipient female monkeys. The ICM-depleted recipient embryos received ICMs of SCNT blastocysts (isolated by immunosurgery) and gave rise to viable cloned blastocysts after the second round of micromanipulation (Fig. 1). From a total of 11 SCNT blastocysts, three early gestations were found in two recipients (one carrying twins) and the singleton cloned pregnancy reached full-term development. The cloned Rhesus monkey named “Retro” was born healthy, reached adulthood, and remains healthy for over 2 years. A careful analysis of the full-term placenta showed exact imprinting control, and adequate DNA methylation levels, thus showing that the strategy culminated in a functional placenta.
The small number of transferred SCNT embryos precluded the estimation of cloning efficiency for comparisons with the previous SCNT protocol (Fig. 1), albeit served as a relevant proof-of-principle work for cloning Rhesus monkeys. The use of fetal fibroblasts as donor cells by Liao and colleagues did not address if adult cells can lead to clones, albeit these cells are attractive for sophisticated genetic engineering before reaching replicative crisis and senescence (Meehan et al., 2008). Nonetheless, this novel SCNT protocol using TR is more technically demanding than previous approaches. This fact suggests that follow-up work should focus on additional attempts to improve epigenetic reprogramming that improves placenta specification, differentiation, and function.
The report of progress in cloning Rhesus monkeys inevitably sheds light to the possibility of human reproductive cloning, which should not be attempted due to numerous ethical concerns. Nonetheless, there are more attractive possibilities that surge from this scientific achievement than reasons for caution. This report shows the potential of functional genomics to explore SCNT reprogramming and improve cloning outcomes. It remains unexplored the potential of fully integrating multiple “omics” datasets (i.e., multi-omics) for exploring epigenetic reprogramming in the context of animal cloning. Nonetheless, this technological advance by Liao and colleagues will likely boost the development of primate models of human late-onset diseases, and perhaps fuel both wildlife cloning and de-extinction efforts.
Footnotes
Authors’ Contributions
M.T.M.: Writing—original draft, review and editing.
Author Disclosure Statement
The author declares no conflicting financial interests.
Funding Information
No information to declare.