
Abstract
Spermatogenesis constitutes a complex and intricate cascade of differentiation, indispensable for the male reproductive competence. The intercellular communication conduits of Sertoli cells (SCs) are pivotal in orchestrating this cascade ensuring sustenance and development of germ cells. Single cells and bioinformatics recently demonstrated articles are used for the regulatory modalities through which SCs modulate spermatogenesis, specifically via androgen receptors (ARs), the transforming growth factor-beta/Smad axis, mitogen-activated protein kinases, cAMP/protein kinase A (PKA), phosphatidylinositol 4,5-bisphosphate 3-kinase (PI3k)/AKT serine threonine kinase (Akt), AMP-activated protein kinase, and AR pathways. Within this framework, homeostasis of gap junction dynamics, cryptic sites and the activities at tight junctions and adherens junctions, with the integrity of the testicular barrier, glucose assimilation, lactate distribution, being governed also along with SC maturation. Disruptions in activities or abnormal concentration in derangements in AR, cAMP/PKA, and PI3k/Akt pathways, and as well as the molecules that comprise them, would present male infertility.
Introduction
Spermatogenesis is a complex physiological cascade unfolding within the seminiferous tubule epithelium, culminating in generating spermatozoa (Oyedokun et al., 2023). This highly complicated process initiates with spermatogonial stem cells (SSCs), derivative from primordial germ cells termed gonocytes. Hence, the cells divide to form spermatogonia now under well-regulated hormonal as well as paracrine functions (Lu et al., 2024; McCarrey, 2023). They continue to make several mitotic divisions and become primary spermatocytes. Following the influence of hormonal functions, they go further into a change to secondary spermatocytes and then to spermatids following meiotic divisions (Hashemi Karoii and Azizi, 2023; McCarrey, 2023). Finally, all the spermatids are all set to go through a wide morphological and functional differentiation called spermiogenesis to be known as spermatozoa proper (Meroni et al., 2019). A distinct population of stem cells, endowed with the self-renewal capacity and pluripotency pivotal to the ontogenesis of male reproductive capabilities, essential in the propagation of genetic information to the progeny. These cells are strategically positioned along the basal membrane of the seminiferous tubules, maintaining intimate proximity to Sertoli cells (SCs) and are subject to stringent regulation within a specialized “niche” microenvironment. Furthermore, the testicular milieu harbors several numbers of intratubular, as well as interstitial somatic cells, supportive in the maintenance of normal spermatogenesis (Lin et al., 2016). SCs are the most important of the somatic cells in a testis and the only kind of somatic cell in seminiferous tubules, described in the 19th century by Enrico Sertoli, with features such as having an irregular shape. SCs attach to the basal layer, extending their apical ends to about near the lumen of the seminiferous tubule. SCs play many important functions in spermatogenesis.
SSCs need a special niche to self-renew and differentiate. The secretion of growth factors such as Glial cell line-derived neurotrophic factor (GDNF), fibroblast growth factor 2 (FGF-2), bone morphogenetic protein 4 (BMP4), stem cell factor, epidermal growth factor, and paracrine signals makes the processes of the SCs in niche formation and the refreshing balance between SSCs proliferation and differentiation central (Zheng et al., 2018). Besides, in the mammalian testes, spermatogenesis development is carried out by the unique structure of tight junctions, called the blood–testis barrier (BTB), located between adjacent SCs, which builds a suitable environment. This structure acts to physically divide the seminiferous tubule epithelium into basal and lateral sections with a physiological barrier that provides a niche with a specific composition required by the germ cells. The latter looks for SSC niches in those areas of the seminiferous tubules that abut interstitial tissue (Wang et al., 2019a). In the BTB, these adherens junctions (AJs) are regarded as ectoplasmic specializations (ES) and are distributed into the basal ES interface occurring at the junctions between SCs and the apical ES interface occurring at the junctions between SCs and spermatids (Wen et al., 2018). In the mammals’ testicles, basal ES, desmosomes, cleft junctions, and tight junctions between SCs form the BTB (Shen et al., 2018). In the last decades, most researchers have investigated the spermatogenesis synchronization, but few have considered the roles of SCs (Ahmed et al., 2018). So far, many signaling pathways have been found in SCs, including the androgen signaling pathway, AMP-activated protein kinase (AMPK) signaling pathway, follicle stimulating hormone (FSH)/adenylyl cyclase/cyclic adenosine monophosphate/protein kinase A (PKA) signaling pathway (Deng et al., 2018), hippo signaling pathway (Sharma et al., 2019), signaling pathway by integrin (Levasseur et al., 2017), janus kinases signal transducer, transcription signaling pathway activator (Sharma and Majumdar, 2017), mitogen-activated protein kinases (MAPK) (Lei et al., 2018), κB signaling pathway (Adegoke et al., 2018), nitric oxide/soluble guanylyl cyclase/guanosine monophosphate/protein kinase signaling pathway(Gao et al., 2017), notch signaling pathway, phosphatidylinositol 4,5-bisphosphate 3-kinase (PI3k)/AKT serine threonine kinase (Akt) signaling pathway(Icli et al., 2019), The transforming growth factor-beta/SMAD signaling pathway (TGF-β/SMAD), and Wnt signaling pathway (Luo et al., 2020b). Summarization of pathways that are involved in SC function and spermatogenesis in Table 1 and Table 2.
The Function of Sertoli Cells in Modulating Signal Transduction During Spermatogenesis As Well As the Influences on the Self-Renewal and Differentiation of Stem Cells
SCs regulate spermatogenesis by many mechanisms including ARs, the TGF-β/Smad Axis, MAPK, cAMP/PKA, PI3k/Akt, AMPK, and AR pathways.
AMPK, AMP-activated protein kinase; AR, androgen receptors; BTB, blood–testis barrier; CaMKKβ, Ca2+/calmodulin-dependent protein kinase kinase β; ERK, extracellular signal-regulated kinase; FAAH, fatty acid amide hydrolase; FGF-2, fibroblast growth factor 2; FSH, follicle stimulating hormone; GCs, germ cells; GDNF, glial cell line-derived neurotrophic factor; GLUT, glucose transporter; ICAM, intercellular adhesion molecule 1; IL-1α, interleukin-1 α; JAM-B, junctional adhesion molecule-B; JNK, c-Jun N-terminal kinase; LDH, lactate dehydrogenase; MAPK, mitogen-activated protein kinase; MCT, monocarboxylate transporter; mTOR, mechanistic target of rapamycin; PCNA, proliferating cell nuclear antigen; PI3k/Akt, phosphatidylinositol 4,5-bisphosphate 3-kinase/AKT serine threonine kinase; SCs, Sertoli cells; T3, 3,3,5-triiodothyronine; TGF-β, transforming growth factor-beta; TNF-α, tumor necrosis factor-alpha; ZO-1, zonula occludens-1.
Signaling Pathway Related to Proliferation and Differentiation in Sertoli Cell
TGF-β/SMAD Signaling Pathway
TGF-β is an imperative cell constituent with diverse roles that encompass cellular development, differentiation, and tissue architecture. The cytokines constituting the TGF-β family are secreted from a range of cells, including macrophages, keratinocytes, and fibroblasts. These cytokines orchestrate and modulate a spectrum of crucial biological processes, including cellular homeostasis, differentiation, proliferation, tissue restoration, angiogenesis, programmed cell death (apoptosis), and inhibition of proliferation. In addition, they contribute to the stimulation of the synthesis of the extracellular matrix and play roles in immunomodulation and immunosuppression (de Freitas et al., 2016). TGF-β contains more than 40 members including three hemdimeric TGF-β proteins (TGF-β1, -β2, and -β3), activin family, inhibin, BMPs, growth and differentiation factors (GDF), and TGF-β hemodimer proteins. In this pathway, the process induces through Smad-dependent and non-Smad-dependent pathways (Zheng et al., 2019). Smad1,3,5,6,7,8, active at birth, stages V, VII, VIII, and XV. TGF-β receptors (TGF-βR1, R2) are dual-acting kinases that exhibit both tyrosine kinase activity and threonine/serine kinase activity (Ni et al., 2019). After closing the heterodimeric complex of TGF-βR1, R2 receptors, a signaling cascade is formed, that begins with phosphorylation by R2 and causes TGF-βR1 serine autophosphorylation, and during this process, a number of phosphorylation proteins (especially the family of Smad transcription factors) are activated (Pierucci-Alves et al., 2018). TGF-βR2 actuates SMAD2 and SMAD3. Smads join up with a co-SMAD termed SMAD4 that activates TGF-βs. SMAD2/3 complexes together with subtranscription factors can trigger genes involved in differentiation. The TGF-β/SMAD signaling pathway affects stem cell capacity during spermatogenesis. Gja1, encoding connexin 43, influences SSC proliferation indirectly by forming gap junctions that facilitate intercellular communication, synchronize cell cycles, and maintain tissue homeostasis, but it does not directly initiate DNA synthesis. In contrast, FGF-2 directly promotes DNA synthesis and cell proliferation by binding to its receptor on the cell surface, triggering downstream signaling pathways such as MAPK/extracellular signal-regulated kinase (ERK). FGF-2 acts through both autocrine and paracrine mechanisms to regulate cell cycle progression and DNA replication, making its influence on cellular proliferation more direct than that of Gja1 (Fig. 1). Different members of the TGF-β superfamily may perform their functions at various stages (Morikawa et al., 2016; Yang et al., 2019).
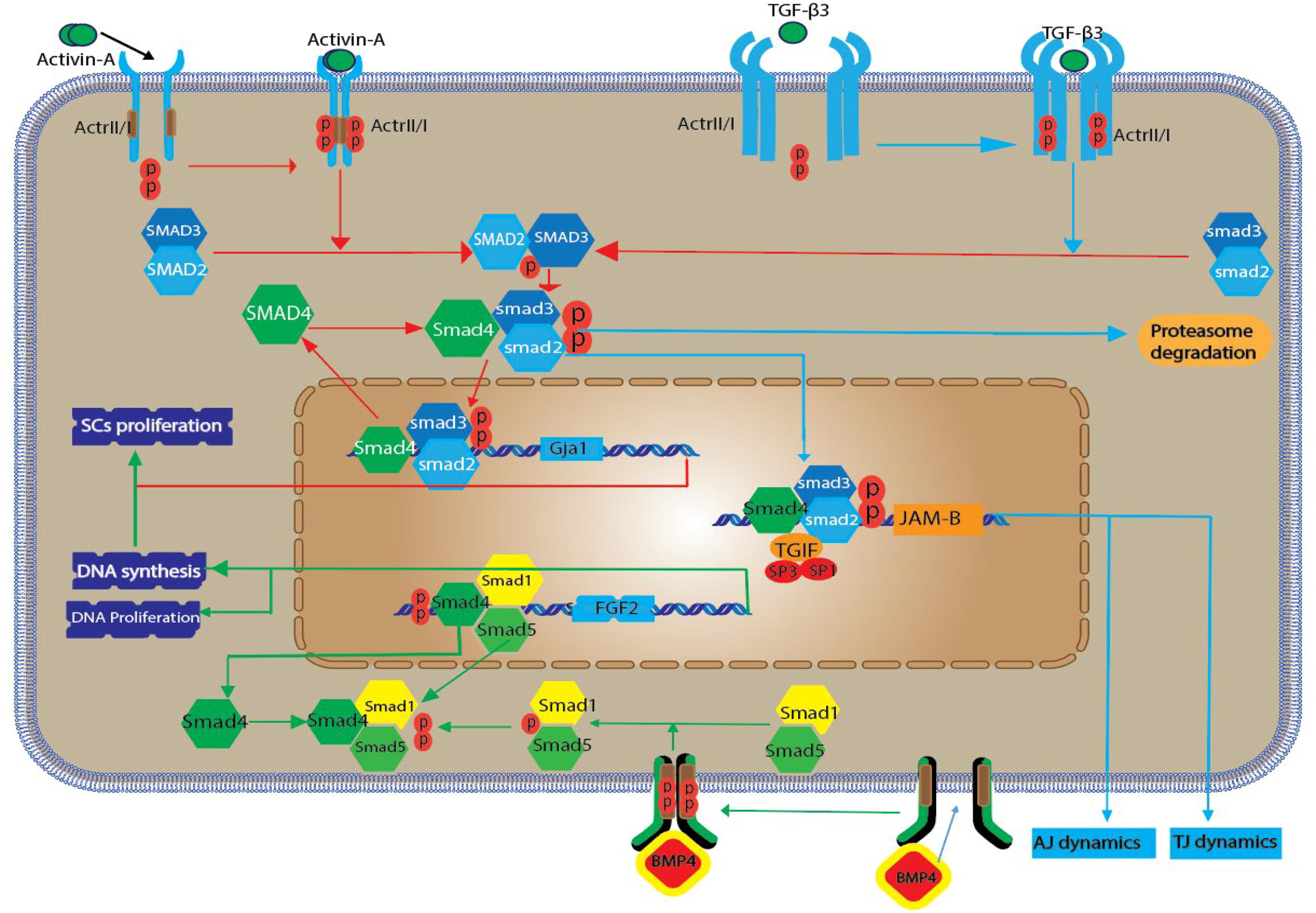
The schematic delineates the intricate processes by which the TGF-β/SMAD signaling cascade influences spermatogenesis within SCs. It elucidates that ligands of the TGF-β family initiate this cascade by engaging with type II (TGFβRII) and type I (TGFβRI) serine/threonine kinase receptors. This interaction is specific, with different TGF-β isoforms selectively binding to distinct receptor complexes. Upon activation, all TGFβRIs are competent to catalyze the phosphorylation of SMAD family members, which are categorized into three distinct classes: receptor-regulated SMADs (R-SMADs) encompassing the co-mediator SMAD (Co-SMAD), represented solely by SMAD4; and the inhibitory SMADs (I-SMADs), namely SMAD6 and SMAD7. Subsequent to R-SMAD phosphorylation by TGFβRI, the R-SMAD dissociates from the receptor and forms a complex with SMAD4. This heteromeric complex translocates to the nucleus, where it associates with the promoter regions of target genes, modulating transcription in concert with additional transcriptional co-regulators, as indicated by the color red. The activin molecule specifically interacts with type IIA activin receptors, triggering the activin/SMAD2/SMAD3 pathway, which upregulates the transcription of genes such as Gja1 and Serpina5, thereby influencing the maturation of the Sertoli cell barrier. Similarly, BMP4 engages the BMP4/SMAD1/SMAD5 axis through its binding to BMPRII, culminating in DNA replication and SC proliferation. The influence of Gja1 on DNA synthesis and SSC proliferation is indirect. Gap junctions, formed by connexin 43, can modulate the cellular environment by synchronizing cell cycles, spreading proliferation signals and FGF-2 activates pathways such as MAPK/ERK, which promote cell cycle progression and DNA replication. TGF-β2 and TGF-β3 are highlighted (in green) as they are known to suppress the expression levels of JAM-B. Specifically, TGF-β3 precipitates the degradation of the JAM-B protein via the ubiquitin-proteasome pathway through the TGF-β3/SMAD2/SMAD3 signaling route. Meanwhile, TGF-β2 modulates SMAD3 activity, which competes with transcription factors Sp1 and Sp3, resulting in the repression of JAM-B transcription, as depicted in blue. BMP, bone morphogenetic protein; ERK, extracellular signal-regulated kinase; FGF-2, fibroblast growth factor 2; JAM-B, junctional adhesion molecule-B; MAPK, mitogen-activated protein kinase; SCs, Sertoli cells; TGF-β, transforming growth factor-beta.
Division and regulation of SCs
SMAD2/3 signaling helps differentiate SCs via cell proliferation and growth. Itman et al. have found that nuclear accumulation of SMAD2 and SMAD3 was induced by activin in postmitotic mouse SCs and also screened and isolated the activin target genes Gja1 and Serpina5 through microarray analysis (Hilbold et al., 2020). These two genes in order encode the connexin 43 gene and the serine protease inhibitor, which require SCs for maturation. In SMAD3, delayed SCs differentiation and decreased size of the testicle are associated with inhibition of androgen receptors (ARs) and SMAD2 expression. Discovering the link between SMAD2/3 and their connective may be useful in investigating the effects of SC proliferation on sperm production. BMP4 and BMP6 cause the proliferation and synthesis of DNA in stem cells through an endocrine pathway (Yang et al., 2016). BMP4 has been shown to increase the phosphorylation of SMAD1/5 and proliferation of human SC, but noggin (BMP4 antagonist) is known for its inhibitory effects. FGF-2 and SCF production were also suppressed when Hay et al. removed BMP4 in human stem cells (Zhang et al., 2018). However, the contribution of the SMAD1/5 pathway versus the ID2/3 pathway in increasing SCs proliferation, because of BMP4, has not been evaluated in their research. This problem also exists in the BMP6 study, especially, if the SMAD2/3 signaling pathway directly mediates BMP6-induced proliferation and also increases CSF levels and GDNF in human SCs (Wang et al., 2017) (Fig. 1).
Activity in tight junctions and AJs
TGF-βs and GDF9 are involved in regulating the activity of tight junctions (TJs) and AJs. Due to the fact that the transport of spermatocytes through BTB requires the opening and closing of the cell junctions, the regulation of junctional adhesion molecule-B (JAM-B) expression is very important for the migration of GCs (Ebnet et al., 2018). TGF-β2 and TGF-β3 reduce JAM-B expression through the SMAD3 signaling pathway. Wang and Lui have discovered that TGF-β2 acts as an anti-expression agent of JAM-B at the pre-transcriptional level in rat SCs (Kudipudi et al., 2019) (Fig. 1). TGF-β2 increases the level of SMAD3 phosphorylation, which competes with transcription factors Sp1 and Sp3 for the role of TG interacting factor (TGIF) and ultimately suppresses JAM-B transcription. However, TGF-β3 repair can reduce JAM-B protein levels in a posttranslational manner in mouse SCs and degrade JAM-B by administering proteasome inhibitors, including MG-132 and lactacystin. This process requires SMAD3/4 activation so deletion of SMAD3 and SMAD4 inhibits TGF-β3-induced JAM-B degradation (Garcia et al., 2017). As a result, TGF-β2 and TGF-β3 may form a regulatory network for BTB degradation and synthesis via the SMAD signaling pathway. Apart from JAM-B, SMAD3 also helps transport preleptotene spermatocytes by reducing CLDN11 expression. CDN11 is crucial for the migration of spermatocytes through the BTB to the adluminal compartment (Pörtner et al., 2020).
AMPK Signaling Pathway
AMPK, functioning as a pivotal regulator of cellular energy homeostasis, orchestrates lactate secretion and maintains the integrity of TJs within SCs. It is, in fact, a heterotrimeric serine/threonine-specific protein kinase. Disruption of this critical pathway can have detrimental consequences on testicular function and spermatozoa viability (Nishimura and L’Hernault, 2017). It has been observed that if α1AMPK is deleted in mouse SCs, mutant mice show thin-headed spermatozoa, decreased expression of binding proteins (β-catenin, vimentin, occludin, and zonula occludens-1 [ZO-1]) and regulatory problems such as energy homeostasis. These findings confirm the involvement of the AMPK signaling pathway in SC during spermatogenesis (Fig. 2). The members of the AMPK large family perform their functions in different stages (Bertoldo et al., 2016; Eid et al., 2019).
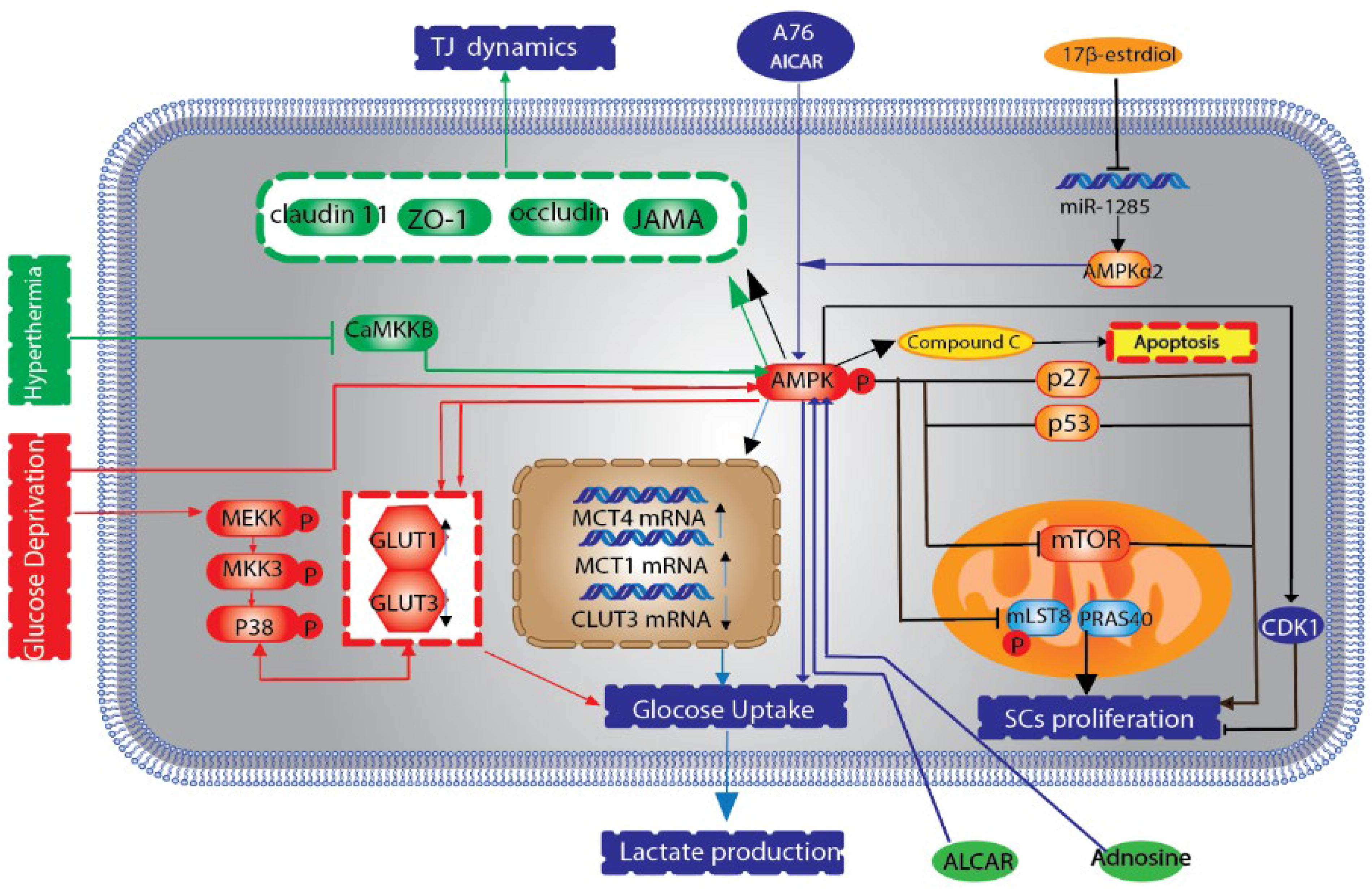
The schematic delineates the influence of the AMPK signaling cascade on spermatogenic processes in SCs. Activation of AMPK is prompted by elevated intracellular AMP concentrations. This activation is further modulated by phosphorylation through kinases such as LKB1, in concert with AMP binding, which collectively potentiates AMPK (depicted in green). A paucity of glucose orchestrates the modulation of GLUT3 while concurrently precipitating the activation of AMPK and the p38 MAPK pathways, which in turn recalibrate GLUT1 expression. Induction of the AMPK signaling cascade via AICAR adenosine analogues augments the expression of GLUT1 and MCT4, albeit diminishing MCT1 and GLUT3 mRNA expression, resulting in enhanced glucose assimilation and lactate efflux (highlighted in red). Additionally, adenosine fosters lactate excretion and reinforces the stabilization of ZO-1 within the SC membrane by engaging the AMPK route. Hypertensive conditions undermine CaMKKβ, thus impeding the AMPK pathway and subsequently influencing the expression of tight junction proteins. On the contrary, 17β-estradiol mitigates miR-1285 expression to sustain α2AMPK levels, thereby preserving AMPK activation. This sustained activation escalates the expression of p53 and p27, yet suppresses mTOR and Skp2 expression, culminating in a reduction of Sertoli cell numbers (illustrated in black). AMPK, AMP-activated protein kinase; CaMKKβ, Ca2+/calmodulin-dependent protein kinase kinase β; GLUT, glucose transporter; LKB1, liver kinase B1; MCT, monocarboxylate transporter; mTOR, mechanistic target of rapamycin; ZO-1, zonula occludens-1.
Lactate production
SCs mainly provide support to spermatocytes and spermatids by converting glucose to lactate, which acts as the main energy source for these cells. The regulation of glucose metabolism during this conversion is critically managed by glucose transporters (GLUTs), which modulate the entry of glucose across cellular membranes. Meanwhile, the diffusion of lactate to germ cells is regulated by monocarboxylate transporters (MCTs), controlling the movement of lactate within the cellular milieu (Luo et al., 2020a). It has also been proven that adenosine monophosphate and AICAR, its analog, induce lactate secretion through AMPK activation. AICAR can produce lactate through AMPK-induced glucose uptake in rat stem cells by increasing GLUT1 protein levels and mRNA levels of MCT4 and decreasing mRNA levels of MCT1GLUT3. In general, adaptation to the environment and response to molecules through the AMPK signaling pathway in the SC provides an appropriate lactate source for energy reserve in the GC (Mateus et al., 2018; Yu et al., 2018) (Fig. 2).
Activities of TJs and AJs
The AMPK signaling cascade plays a crucial role in sustaining the intricate connections within the testicular structure. When AMPK is activated by adenosine, it fortifies the presence of ZO-1 at the membranes of SCs, whereas the inhibition of AMPK through compound C leads to a reduction in adenosine-associated ZO-1 levels (Duan et al., 2017). In addition, heat stress can impair the function of TJs in the testis, which is restored to normal by inhibition of the Ca2+/calmodulin-dependent protein kinase kinase β (Jin et al., 2020). Yang and colleagues discovered that elevated temperatures suppress the AMPK signaling pathway, leading to decreased levels of CLDN11, JAMA, occludin, and notably ZO-1 in pig SCs. Furthermore, the study delineated the interaction among the 26S proteasome inhibitor bortezomib, the AMPK pathway, and AJs within SC and germ cells in mice (Zhang et al., 2021). Bortezomib has been shown to stimulate AMPK, which subsequently triggers the ERK signaling cascade within granulosa cells (germ cells). Over time, this process can lead to dysregulation of AJs, resulting in a reduced sperm count (Yang et al., 2018; Yu et al., 2018).
MAPK Signaling Pathway
MAPKs are a constituent of the serine/threonine-specific protein kinase family. Within mammalian cells, four principal MAPK families have been identified: the prototypical ERK1/2, the stress-activated protein kinase/c-Jun N-terminal kinases (JNKs), the p38 MAPK isoforms (α, β, γ, and δ), and the ERK5. Each family is characteristically activated via a distinct MAPK kinase, also known as MAPKK or MAP2K. In the context of rat testicular physiology, SCs exhibit signal transduction pathways that include ERK1, JNK1, p38 MAPK, and JNK3 (Wang et al., 2019b). According to the data, most of the genes associated with the MAPK pathway are present in immature mouse stem cells, which indicate the presence of MAPK signaling cascades in SCs (Fig. 3) (Gautam et al., 2018; Wang et al., 2018).
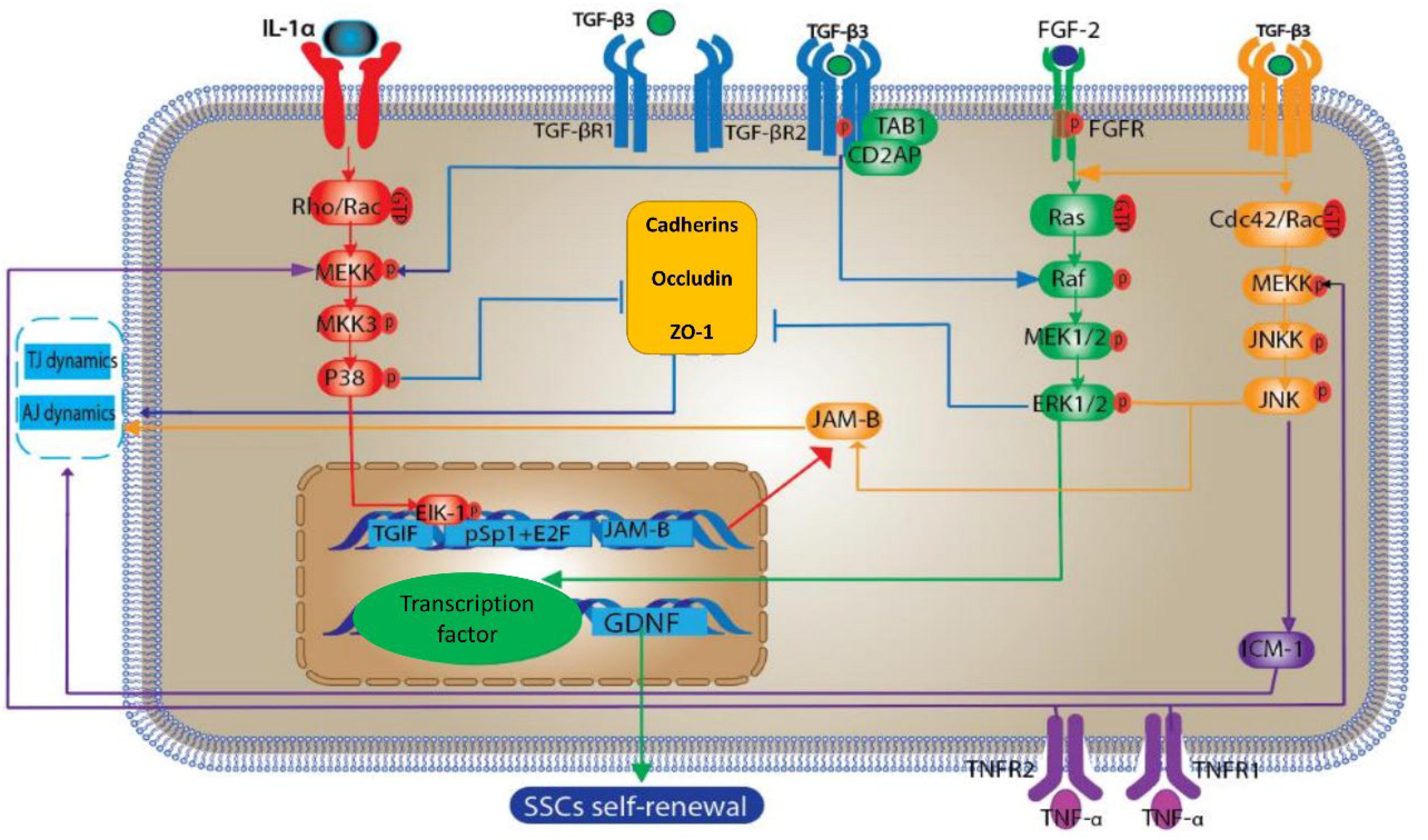
The schematic delineates the influence of the MAPK signaling cascade on spermatogenic processes within SCs. This cascade encompasses JNKs, ERKs, and p38 MAPKs. Sequential activation occurs through phosphorylation, commencing with MAPK kinase (MAP2K), which in turn activates MAPK. Subsequently, activated MAPK phosphorylates a specific substrate (indicated in red). IL-1α is actuated via the p38 MAPK route. Furthermore, p38 MAPK catalyzes the phosphorylation of Elk-1, facilitating its association with TGIF and the pSp1 + E2F complex, culminating in the enhancement of JAM-B transcription. Activation of ERK and JNK pathways by transforming TGF-β3 leads to messenger RNA anomalies in JAM-B. Concomitant interaction of TGF-β activated kinase 1 (TAB1) and CD2-associated protein (CD2AP) with the TGF-β3 receptor complex results in the downregulation of ZO-1 and cadherin expressions, disrupting AJs between SCs and GCs, and the BTB. TNF-α attenuates ZO-1 expression via the p38 MAPK pathway (denoted in blue), and concurrently upregulates ICAM-1 through the JNK pathway, thereby modulating the activity of AJs and TJs (represented in purple). Lastly, FGF-2 initiates the ERK pathway to promote the expression of GDNF, which in turn stimulates the proliferation of SSCs. AJs, adherens junctions; BTB, blood–testis barrier; GCs, germ cells; GDNF, glial cell line-derived neurotrophic factor; ICAM, intercellular adhesion molecule 1; IL-1α, interleukin-1 α; JNKs, c-Jun N-terminal kinases; SSCs, spermatogonial stem cells; TGIF, TG interacting factor; TJs, tight junctions; TNF-α, tumor necrosis factor-alpha.
MAPK p38 Signaling Pathway: TJs and AJs Activity
The p38 MAPK signaling cascade is implicated in an array of regulatory pathways that modulate JAM-B expression. Previous sections delineated the mechanism by which both TGF-β2 and TGF-β3 attenuate JAM-B expression via the TGF-β/SMAD signaling axis. In this context, the role of the p38 MAPK pathway is augmented, playing a critical role in the upregulation of interleukin-1α and subsequent transcriptional activation of JAM-B in murine stem cells (Zhou et al., 2019). The active p38 MAPK phosphorylates the ETS domain transcription factor (Elk-1). Phosphorylation allows Elk-1 to affect TGIF and proximal Sp1 + E2F function. These interactions eventually increase JAM-B transcription by activating Sp1 and NRSF. We can also mention two other MAPK subfamilies, JNK and ERK, for their effects on JAM-B mRNAs through posttranscriptional regulation on TGF-β3 stimulation in mouse SCs (Tao et al., 2019).
AR Signaling
AR signaling plays a crucial role in the regulation of spermatogenesis, which is a complicated and necessary process for male fertility. This signaling occurs specifically inside SCs. When androgens attach to the AR, the AR experiences a change in its shape, causing it to separate from heat shock proteins. After that, it forms homodimers. These AR dimers go into the nucleus and attach to androgen response elements in the promoter regions of certain genes. This process affects the activity of gene transcription, either increasing or decreasing the expression of the genes. Transcriptional regulation plays a crucial role in creating and maintaining the optimal conditions necessary for spermatogenesis in the seminiferous tubules (Tao et al., 2019).
AR signaling in SCs triggers a series of subsequent events that have a substantial effect on cellular processes. A crucial mechanism includes the initiation of Src kinase, which adds phosphate groups to the epidermal growth factor receptor (EGFR) (Wang et al., 2022). The phosphorylation event acts as a stimulus for the Ras kinase pathway, which then initiates the activation of the MAPK signaling cascade. Upon activation, the MAPK pathway leads to the phosphorylation and subsequent activation of transcription factors, including cAMP-response element-binding protein (CREB) (Zhou et al., 2019). After being activated, CREB moves into the nucleus and affects the transcription of genes that are important for cell proliferation, differentiation, and survival. These activities are essential for the maintenance and growth of SSCs and the overall development of spermatogenesis. The AR signaling pathway goes beyond just regulating gene transcription and also triggers other important signaling pathways. A crucial route includes the activation of Src kinase, which then phosphorylates the EGFR. Phosphorylation plays a crucial role in activating the Ras-MAPK signaling cascade. Once initiated, the MAPK pathway transmits a series of phosphorylation events, resulting in the activation of the CREB. Once CREB is activated, it moves into the nucleus and controls the transcription of genes that control important cellular functions including proliferation, differentiation, and survival. The complex network of AR-mediated signaling is essential for the preservation and appropriate functioning of SSCs and overall spermatogenesis (Wang et al., 2019b).
Recent studies underscore the importance of AR signaling in SCs for proper spermatogenesis. For example, research by O’Hara and Smith (2015) highlights that disrupting AR expression in SCs leads to impaired spermatogenesis and infertility in male mice, further emphasizing the role of AR-mediated transcriptional regulation in the maintenance of SSCs and the broader spermatogenic process. Additionally, studies have shown that AR signaling can influence the expression of growth factors such as GDNF and FGF-2, which are vital for SSC maintenance and proliferation (Jin et al., 2020) (Fig. 4).
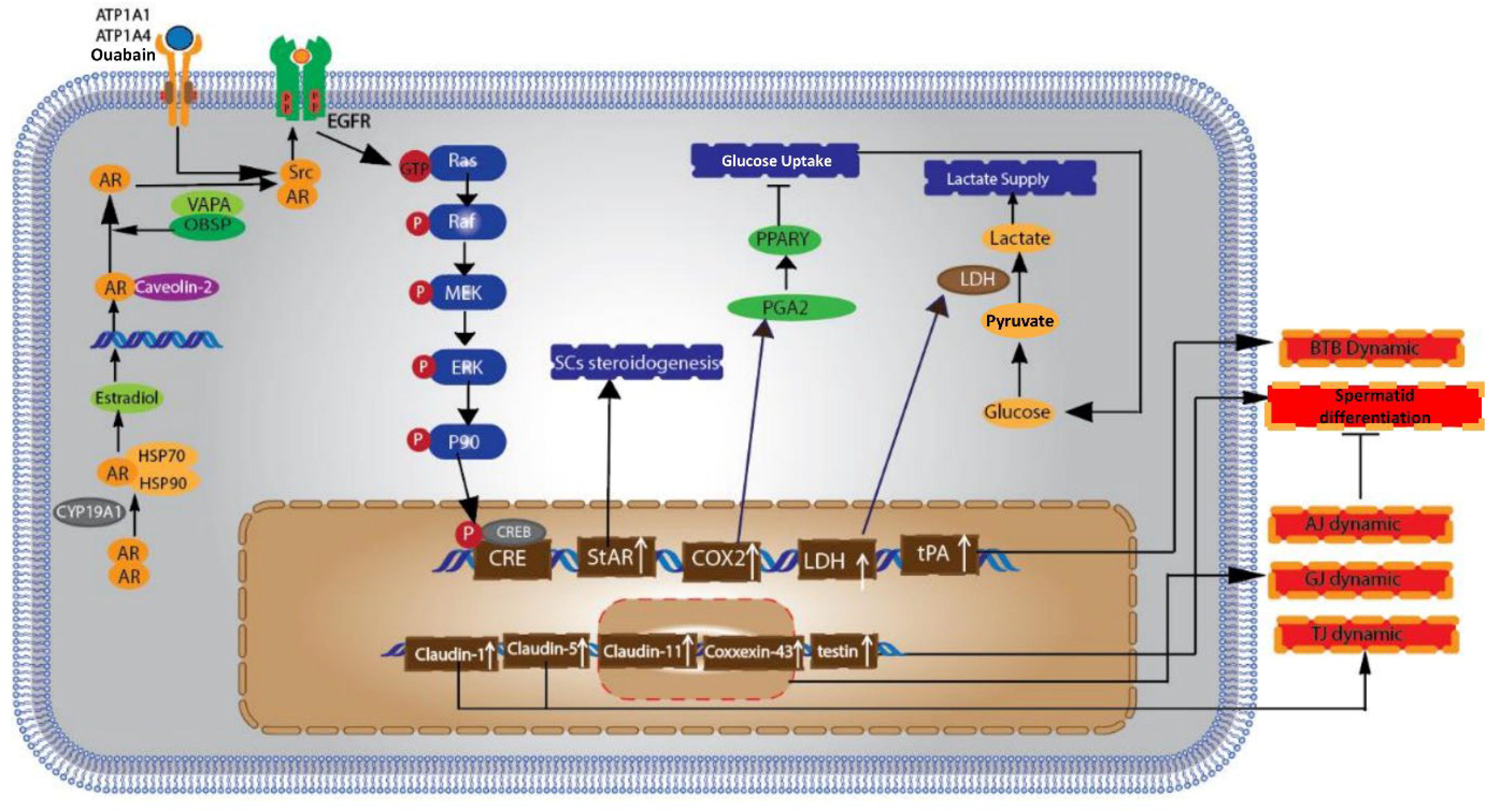
The schematic delineates the impact of the AR signaling cascade within SCs on spermatogenetic processes. Initially, AR dissociates from heat shock proteins and translocate to the nucleus following dimerization. Subsequently, the AR dimer engages with ARE in the genomic loci of target genes, thereby modulating their transcriptional activity—either inducing or repressing it. AR signaling facilitates the activation of Src kinase, which, in turn, catalyzes the phosphorylation of the EGFR. This phosphorylation event triggers the activation of Ras kinase, leading to the initiation of the MAPK signaling cascade. Culminating the sequence of phosphorylation events, the activated MAPK translocate into the nucleus where it stimulates the CREB, effectuating transcriptional responses. AR, androgen receptor; ARE, androgen response elements; CREB, cAMP-response element-binding protein, EGFR, epidermal growth factor receptor.
PI3k/Akt Signaling Pathway
The phosphoinositide 3-phosphate kinase (PI3K) signaling axis orchestrates SSC equilibrium, modulating the dichotomy between SSC proliferation and differentiation, thereby underpinning the incessant genesis of spermatozoa. This pathway is pivotal for spermatogenesis, with aberrations in PI3K signaling within the testicular milieu potentially precipitating an escalation in differentiation and a concomitant diminution in SSC populations or unchecked SSC expansion, culminating in male sterility or oncogenesis within testicular tissue. The initiation of PI3K/Akt signaling transpires in the spermatogonial constituents residing within the testicular niche, instigated by the release of glial cell line-derived neurotrophic factor (GDNF) by SCs.
Numerous researches indicate that the PI3k/Akt signaling pathway is influenced by FSH, insulin, and insulin-like growth factor (IGF). Initially, the attachment of IGF and insulin to their receptor, IGFR, triggers the phosphorylation of insulin receptor substrate 1, followed by the activation of PI3K (Cannarella et al., 2019; Cannarella et al., 2018;Wang et al., 2020). The phospholipids known as phosphatidylinositol-4,5-bisphosphate undergo a transformation into phosphatidylinositol-3,4,5-triphosphate (PIP3) as a result of phosphorylation by PI3K. Subsequently, this PIP3 instigates a series of phosphorylation events involving phosphoinositide-dependent kinase 1, Akt, and associated downstream signaling molecules (Yang and Yang, 2020) (Fig. 5).
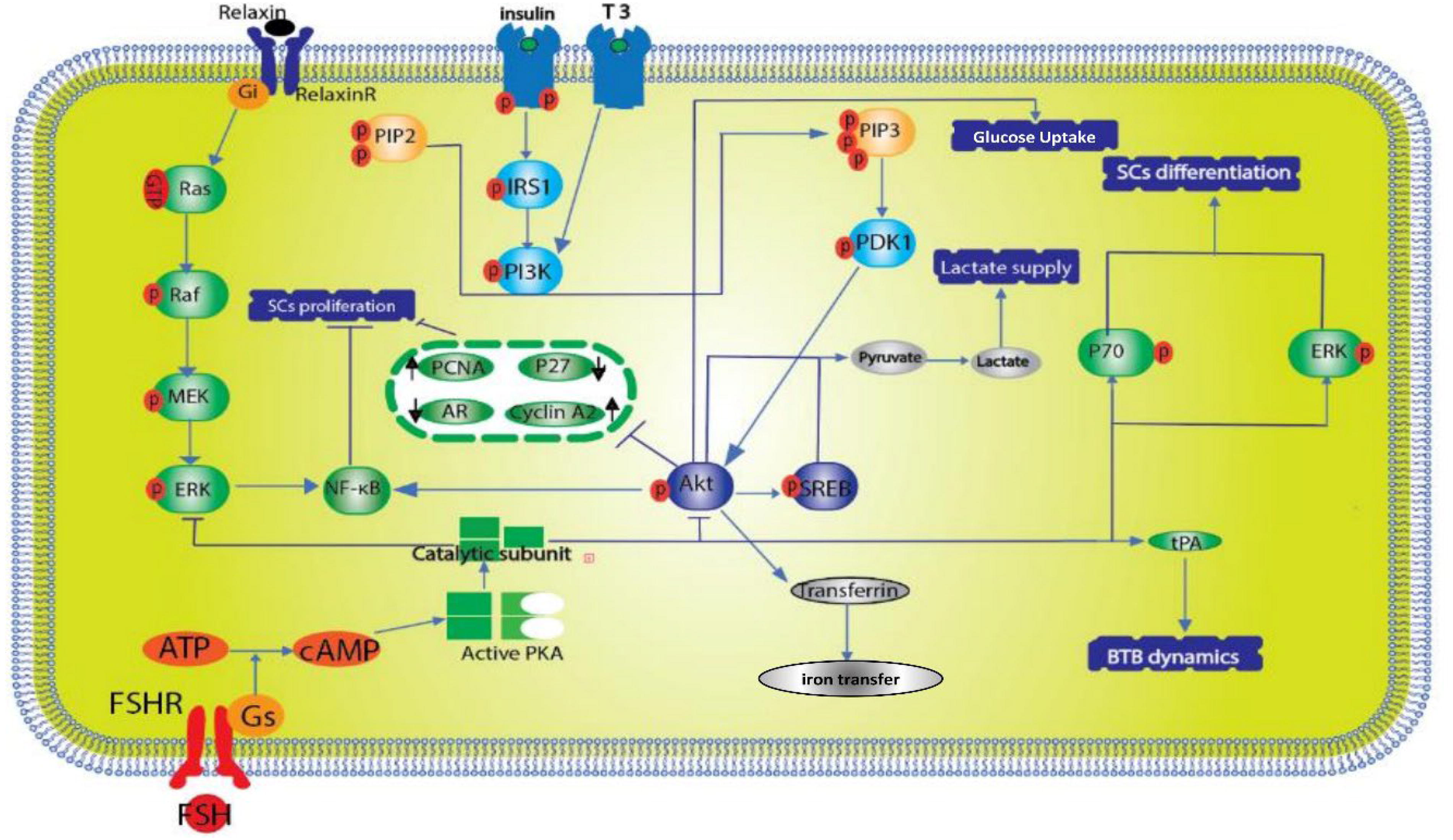
Scientific research has demonstrated that FSH and relaxin exert a significant influence on the differentiation of SCs by modulating the ERK1/2 and PI3k/Akt signaling pathways. Specifically, FSH enhances glucose assimilation, LDH activity, and transferrin synthesis. T3 downregulates cyclin A2 and PCNA expression via PI3k inhibition, while upregulating p27kip1 and AR expression, culminating in alterations to SC maturation. Additionally, PGD2 amplifies the transcription and enzymatic functionality of LDH through the PI3k/Akt/CREB signaling axis. Furthermore, FSH augments tPA expression, potentially influencing the dynamics of the BTB (indicated in blue). FSH, follicle stimulating hormone; LDH, lactate dehydrogenase; PCNA, proliferating cell nuclear antigen; PGD2, prostaglandin D2; PI3k/Akt, phosphatidylinositol 4,5-bisphosphate 3-kinase/AKT serine threonine kinase; T3, triiodothyronine; tPA, tissue plasminogen activator.
Mechanistic Target of Rapamycin Signaling Pathway
The mTOR signaling pathway, also known as the mechanistic target of rapamycin signaling route, plays a crucial role in regulating SSCs, which are the fundamental cells responsible for sperm generation in males. This pathway regulates many essential activities required for preserving SSC activity and promoting spermatogenesis. mTOR combines signals pertaining to nutrition availability, growth factors, and cellular energy levels in order to control SSC activities. The pathway exerts an effect on cell growth, proliferation, and survival via its two complexes, mTORC1 and mTORC2. mTORC1, specifically, is responsive to food levels and growth cues, stimulating protein synthesis and cell development by activating targets such as S6K1 and suppressing 4EBP1. This guarantees that SSCs possess the necessary resources and circumstances for accurate division and differentiation. mTOR also influences metabolic processes, ensuring that energy consumption is synchronized with the substantial requirements of spermatogenesis (Zhang et al., 2021).
Disruption of mTOR signaling has a substantial impact on spermatogenesis. Excessive activation of mTORC1 may cause an increase in cell growth but impede the normal process of cell specialization, leading to a decrease in the quantity of fully functioning sperm. In contrast, inadequate mTOR activity might hinder the preservation and functioning of SSCs, which may result in infertility. Therefore, it is crucial to regulate this pathway properly in order to maintain the SSC pool and ensure efficient spermatogenic development. Gaining a comprehensive understanding of these systems is crucial for effectively treating reproductive concerns and devising precise therapies for associated illnesses.
Conclusion
SCs play a pivotal role in the sustenance, nourishment, and safeguarding of spermatogenic cells via an array of signaling cascades. Accumulated research over the previous decade has illuminated the salient roles of the MAPK, AMPK, and TGF-β/SMAD pathways in the orchestration of spermatogenesis. Particularly, the MAPK cascade is instrumental in modulating the functions of occluding and anchoring junctions, alongside the proliferation and meiosis of germinal cells, and the propagation and lactate synthesis within SCs. Concurrently, both the AMPK and TGF-β/SMAD pathways exert influence over the dynamics of junctional complexes and the proliferation of SCs, with AMPK specifically governing lactate provision. Interactions among TGF-β/SMAD, AMPK, MAPK, AR, and additional cascades within SCs are crucial for spermatogenesis, as they regulate the activity of cellular junctions, proliferation of SC and germ cell, and lactate distribution. These pathways coalesce into an intricate regulatory network essential for spermatogenesis.
Authors’ Contributions
D.H.K: Assembly of data, experiment design, data analysis, article—writing and editing. G.J.: Data analysis and article editing. H.Z.: supervisor and edited the article. F.F.M.A.: Data analysis and article editing. M.A.: Data analysis and article editing.
Ethical Approval
This article does not contain any studies with human or animal participants performed by any of the authors. All procedures were performed in accordance with the ethical standards of the responsible committee on human experimentation and with the Helsinki Declaration.
Consent for Participation
None declared.
Consent for Publication
None declared.
Footnotes
Acknowledgements
Data Availability
None declared.
Author Disclosure Statement
The authors declare that they have no conflicts of interest.
Funding Information
This review was conducted with the financial assistance of the Center for International Scientific Studies and Collaboration (CISSC) under the auspices of the Ministry of Science (grant no. 01DS13003).