
Abstract
This study explores the protective mechanism of mesenchymal stem cell-derived extracellular vesicles (MSC-EVs) in diabetic foot ulcer (DFU). Human umbilical cord MSCs (HucMSCs) were identified via osteogenesis and adipogenic differentiation, as well as flow cytometry. EVs were isolated from HucMSCs and characterized using transmission electron microscopy, nanoparticle tracking analysis, and Western blotting. Fluorescence microscopy revealed the uptake of PKH67-labeled EVs and Cy3-labeled microRNA-21-5p (miR-21-5p) by human skin fibroblasts (HSFs). EVs were cocultured with HSFs, and cell proliferation and migration were assessed using Cell Counting Kit-8, colony formation, scratch, and Transwell assays. miR-21-5p overexpression in EVs was evaluated for its role in promoting HSF functions. The expression levels of miR-21-5p, Krüppel-like factor 6 (KLF6), α-smooth muscle actin, and collagen type I alpha 1 chain were analyzed via quantitative real-time PCR and Western blotting. The interaction between miR-21-5p and KLF6 was confirmed through a dual-luciferase reporter gene assay. HucMSC-derived EVs enhanced the proliferation and migration of HSFs under high glucose by delivering miR-21-5p, which targeted and inhibited KLF6. Overexpression of KLF6 counteracted the pro-proliferative and migratory effects of EVs carrying miR-21-5p. Overall, these findings suggest that HucMSC-EVs promote HSF proliferation and migration by downregulating KLF6 via miR-21-5p delivery, offering a potential therapeutic strategy for DFU.
Introduction
Diabetic foot ulcer (DFU) is a severe and common complication of diabetes, representing one of the leading causes of hospitalization among diabetic patients (Volmer-Thole and Lobmann, 2016). DFU precedes 80% of lower extremity amputations in diabetic patients and contributes significantly to increased mortality rates (Armstrong et al., 2023). While various cell types are involved in the wound healing process, fibroblasts play a central role in tissue repair and reconstruction (Smith et al., 2019). As the primary cells of the skin, fibroblasts are crucial for wound healing, interacting with keratinocytes, endothelial cells, and other cell types during tissue reconstruction (Zhu et al., 2021). In diabetic ulcers, fibroblasts exhibit aberrant phenotypes, including impaired proliferation, altered cytokine secretion, and premature senescence, which hinder the healing process (Abedin-Do et al., 2021).
Mesenchymal stem cells (MSCs) are multipotent stem cells derived from mesodermal tissue with high self-renewal capacity and the ability to differentiate into various cell types (Casado-Díaz et al., 2020). In addition to their multipotent differentiation potential, MSCs exhibit robust in vitro expansion capabilities and possess significant immunomodulatory effects, making them promising candidates for tissue repair and regeneration (Gao et al., 2016). Specifically, MSCs have shown the ability to enhance diabetic wound healing by promoting epithelialization and angiogenesis (Jackson et al., 2012).
Extracellular vesicles (EVs) are nanoscale membrane-bound particles secreted by numerous cell types that transport nucleic acids, lipids, and proteins to modulate signaling pathways and facilitate intercellular communication (Shao et al., 2018; Thery et al., 2018). EVs can be generally split into the following three subtypes based on their mechanism of biogenesis: exosomes, shed microvesicles, and apoptotic bodies (Kalra et al., 2016). In brief, exosomes are formed via the inward budding of the cell plasma membrane and the subsequent formation of multivesicular bodies. Conversely, shed microvesicles are generated via the outward budding of the plasma membrane. Apoptotic bodies are released during the process of cellular apoptosis (Kalra et al., 2016). Most proteins destined to enter the extracellular space or various intracellular compartments must pass through the cellular endocrine pathway. The first step is to recruit and transport cargos from the endoplasmic reticulum (ER) lumen to the Golgi apparatus through the coat protein complex II. Additional ER transmembrane proteins that assist in cargo recruitment are called cargo receptors (Shen et al., 2023; Tao et al., 2023). Surfeit 4 is a polytopic transmembrane protein that primarily resides in the ER membrane and has been implicated in ER export of diverse cargos, such as EPO, α1AT, and proinsulin (Wang et al., 2021). MSC-EVs closely resemble MSCs in their angiogenic, immunomodulatory, and therapeutic functions (Casado-Díaz et al., 2020). Studies have shown that MSC-EVs enhance the proliferation and migration of fibroblasts from chronic wounds in a dose-dependent manner (Shabbir et al., 2015). Moreover, MSC-EVs significantly accelerate wound closure, reduce scar formation, increase collagen deposition, and promote reepithelialization in diabetic wounds compared with controls (Chang and Li, 2023). Despite these promising findings, the precise molecular mechanisms through which MSC-EVs regulate human skin fibroblast (HSF) proliferation and migration in DFU remain unclear. This study aims to investigate the underlying regulatory mechanisms of MSC-EVs in the context of DFU, providing insights into their potential therapeutic applications.
MicroRNAs (miRNAs) are a highly conserved class of noncoding RNAs that play a crucial role in maintaining cellular homeostasis by negatively regulating gene expression (Lu and Rothenberg, 2018). The deregulation of miRNAs has been implicated in the progression of DFU and impaired wound healing (Tang et al., 2022). Emerging evidence suggests that miR-21-5p holds significant potential in promoting skin wound healing (Chen et al., 2022; Li et al., 2022; Ross, 2021; Xie et al., 2022). Overexpression of miR-21-5p has been shown to enhance wound healing in mice by stimulating fibroblast proliferation and migration (Xiao et al., 2021).
EVs loaded with miRNAs have been identified as a promising therapeutic tool to accelerate diabetic wound healing by modulating immune responses and cellular functions (Shen et al., 2022). For instance, miR-21-5p delivered by human adipose stem cell-derived exosomes can promote diabetic cutaneous wound healing (Lv et al., 2020). However, it remains unclear whether miR-21-5p can regulate the proliferation and migration of HSFs via transport through MSC-EVs.
Considering this knowledge gap, this study aims to explore the molecular mechanism by which MSC-EVs, carrying miR-21-5p, influence HSF proliferation and migration. The findings may provide novel insights into the therapeutic potential of MSC-EVs and miR-21-5p for the treatment of DFU.
Materials and Methods
Cell culture and treatment
Human umbilical cord MSCs (HucMSCs) (FH-H042) and HSFs (FH0189) were obtained from Shanghai Fuheng Biotechnology Co., Ltd (Shanghai, China). HucMSCs were cultured in MSC medium (7511, ScienCell, San Diego, CA, USA), supplemented with 10% fetal bovine serum (FBS) and 100 U/mL penicillin/streptomycin. HSFs were cultured in Dulbecco’s modified Eagle’s medium (09211, ScienCell) added with 10% FBS and 100 U/mL penicillin/streptomycin. HSFs were divided into two experimental groups as follows: the high-glucose group (HG) with 30 mM
Identification of differentiation ability of HucMSCs
HucMSCs were seeded at a concentration of 1 × 105 cells/mL in 6-well plates. To induce osteogenic differentiation, the OriCell® Human Stem Cell Osteogenic Differentiation Kit (HUXXC-90021, Cyagen, Jiangsu, China) was used, whereas adipogenic differentiation was induced using the OriCell® HucMSC Adipogenic Differentiation Kit (HUXUC-90031, Cyagen). Osteogenic differentiation was confirmed through Alizarin Red staining, whereas adipogenic differentiation was assessed using Oil Red O staining.
Flow cytometry
HucMSCs were digested with trypsin (R001100, Thermo Fisher Scientific, Waltham, MA, USA), and 1 × 106 cells were resuspended in 1 mL of phosphate-buffered saline (PBS). A 200 µL aliquot of the cell suspension was placed into Eppendorf tubes and incubated in a humidified environment at 37°C with 5% CO2. The surface markers CD90 (1:5000, ab11155, Abcam, Cambridge, MA, USA), CD105 (1:500, ab11415, Abcam), and CD34 (1:2000, ab18227, Abcam) were detected via flow cytometry to confirm the mesenchymal phenotype of the HucMSCs.
Western blot
Cells were lysed using enhanced radio-immunoprecipitation assay lysate (PS0013, Leagene, Beijing, China) containing protease inhibitors. Protein concentration was determined with a bicinchoninic acid (BCA) protein quantification kit (PT0001, Leagene). Equal amounts of protein were separated on 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gels and transferred onto polyvinylidene difluoride membranes. Membranes were blocked with 5% bovine serum albumin to prevent nonspecific binding and incubated overnight at 4°C with primary antibodies. After washing, the membranes were incubated with horseradish peroxidase-conjugated goat anti-rabbit secondary antibody at room temperature for 1 hour. Protein bands were visualized using an enhanced chemiluminescence solution (Millipore, Billerica, MA, USA). Band intensity was quantified using Image-Pro Plus 6.0 (Media Cybernetics, San Diego, CA, USA), with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) serving as the loading control. The primary antibodies used were rabbit polyclonal antibody Krüppel-like factor 6 (KLF6) (1:1000, PA5-79560, Thermo), rabbit monoclonal antibody CD63 (1:5000, ab134045, Abcam), rabbit monoclonal antibody CD81 (1:5000, ab109201, Abcam), rabbit monoclonal antibody CD45 (1:5000, ab40763, Abcam), and rabbit polyclonal antibody GAPDH (1:2500, ab9485, Abcam).
Isolation and identification of EVs
The conditioned medium from HucMSCs was collected and centrifuged at 2000×g for 10 minutes. The supernatant was passed through a 0.22-µm filter (SteritopTM Millipore, MA, USA) to eliminate large debris and dead cells. It was then centrifuged at 10,000×g for 30 minutes to remove any remaining cell debris, followed by ultracentrifugation at 100,000×g for 3 hours at 4°C to isolate EVs. The morphology of the isolated EVs was observed under a transmission electron microscope (TEM, HT7800, HITACHI High-Technologies, Japan). The particle size and concentration of EVs were analyzed using a nanoparticle tracker (NTA, ZetaView, Particle Metrix, Germany). EV protein concentration was quantified using a BCA Assay Kit (PT0001, Leagene), and 50 µg of EVs were used for subsequent experiments. The expression of EV marker proteins CD63 and CD81, as well as the negative marker proteins CD45, was confirmed via Western blot. As a control, conditioned medium (GW) from HucMSCs treated with the EV inhibitor GW4869 (15 µm, 52321ES10, Yeasen, Shanghai, China) was also analyzed.
Cell transfection
The miR-21-5p mimic, KLF6 overexpression plasmid (KLF6), and their respective negative controls (mimic NC and NC) were obtained from GenePharma (Shanghai, China). HucMSCs were transfected with miR-21-5p mimic or mimic NC using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA), after which EVs were isolated from the transfected HucMSCs to obtain EVs-miR-21-5p and EVs-NC. HSFs were transfected with either the KLF6 overexpression plasmid or the NC plasmid. All experiments were conducted 24 hours after transfection.
Endocytosis of EVs
EVs were treated with RNase I (EN0601, Thermo) or Triton X-100 (HFH10, Thermo) for 30 minutes, followed by heat inactivation. The expression levels of miR-21-5p in the EVs were subsequently detected. For tracking EV uptake, 50 µg of EVs were labeled with 4 µg of PKH67 (PKH67GL, Sigma, St Louis, USA) for 5 minutes at room temperature. After washing with PBS, the labeled EVs were recovered by ultracentrifugation and passed through a 0.22-µm filter to remove insoluble substances. HSFs were incubated with the PKH67-labeled EVs in 24-well plates, and the nuclei were counterstained with 4’, 6-diamino-2-phenylindole (DAPI) (D9542, Sigma). Images were captured using a fluorescence microscope (LSM700B, Carl Zeiss, Jena, Germany). Cyanine 3 (Cy3)-miR-21-5p, provided by Sangon (Shanghai, China), was transfected into HucMSCs, and the nuclei were counterstained with DAPI (D9542, Sigma). HucMSCs were incubated with green fluorescent protein (GFP)-labeled HSFs in the presence or absence of the EV inhibitor GW4869 (15 µM, 52321ES10, Yeasen) for 48 hours. Fluorescent images were obtained using a fluorescence microscope (LSM700B, Zeiss).
Cell Counting Kit-8 assay
HSFs were seeded in 96-well plates at 5 × 103 cells/well. After 48 hours of incubation, 10 µL of Cell Counting Kit-8 (CCK-8) reagent (CK04, Dojindo Laboratories, Japan) was added to each well, followed by 3 hours of incubation. Absorbance at 450 nm was measured using a microplate reader (Bio-Rad, Hercules, CA, USA).
Clone formation assay
HSFs were seeded into 96-well plates at 1 × 103 cells/well and incubated at 37°C for 2 weeks to allow colony formation. Colonies were fixed with methanol and stained using 0.1% crystal violet solution. The number of colonies was counted to assess the proliferative capacity of the HSFs.
Cell scratch assay
HSFs were seeded in 12-well plates at 2 × 105 cells/well. Once the cells reached approximately 90% confluence, a straight scratch was made using a 1-mL pipette tip. The wells were gently rinsed with PBS to remove detached cells. Images of the scratch were captured at 0 hours and again at 24 hours postscratch. The migration area was quantified using Image Pro Plus 6.0 software, and the percentage of migration was calculated using the following formula:
Migration area (%) = [(0 hours gap area–24 hours gap area)/0 hours gap area] × 100%.
Transwell assay
The cells were harvested by centrifugation, resuspended in a serum-free medium, and counted. A total of 1 × 104 cells were seeded into the apical chamber of a Transwell insert (BD Biosciences, San Jose, CA, USA), whereas the basolateral chamber was filled with a complete medium containing 10% FBS. After 24 hours of incubation, cells that migrated to the basolateral chamber were fixed with 4% paraformaldehyde and stained with 0.1% crystal violet. The number of migrated cells was imaged and counted using ImageJ software.
Quantitative Real-Time PCR
Total RNA was extracted using the RNeasy Mini Kit (Qiagen, Valencia, CA, USA) and reverse transcribed into cDNA using the Reverse Transcription Kit (RR047A, Takara, Toyko, Japan). For miRNA detection, the miRNA First Strand cDNA Synthesis (Tailing Reaction) Kit (B532451-0020, Sangon, Shanghai, China) was used for reverse transcription to generate complementary DNA (cDNA). Quantitative real-time PCR (qRT-PCR) was performed using the SYBR® Premix Ex TaqTM II (Perfect Real Time) Kit (DRR081, Takara) on an ABI 7500 Real-Time PCR System (ABI, Foster City, CA, USA). Primers were synthesized by Sangon (Shanghai, China), as listed in Table 1. Gene expression was quantified using the 2-ΔΔCt method (Livak and Schmittgen, 2001), with GAPDH or U6 small nuclear RNA (U6) serving as the internal reference (Cai et al., 2020).
Quantitative PCR Primers
miR-21-5p, microRNA-21-5p; KLF6, Krüppel-like factor 6; α-SMA, α-smooth muscle actin; COL1A1, collagen type I alpha 1 chain; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Bioinformatics analysis
The downstream target genes of miR-21-5p were predicted using several bioinformatics tools, including starBase (Cai et al., 2020) (http://starbase.sysu.edu.cn/index.php), miRDB (Chen and Wang, 2020) (https://mirdb.org/), and miRWalk (Sticht et al., 2018) (http://mirwalk.umm.uni-heidelberg.de/).
Dual-Luciferase reporter assay
The 3' untranslated region (UTR) of KLF6, containing the binding site for miR-21-5p, was synthesized and cloned into a pMIR-reporter plasmid (AM5795, Thermo), referred to as KLF6-WT. A mutated version of the KLF6 3' UTR, where the miR-21-5p binding site was altered, was similarly cloned into the pMIR-reporter plasmid, designated as KLF6-MUT. These plasmids were cotransfected into HSFs along with either mimic NC or miR-21-5p mimic. After 48 hours, the cells were lysed, and luciferase activity was measured using the Luciferase Assay Kit (K801-200, BioVision, Mountain View, CA, USA).
Statistical analysis
Data analysis and graphical presentation were conducted using SPSS 21.0 (IBM Corp., Armonk, NY, USA) and GraphPad Prism 8.0 (GraphPad Software Inc., San Diego, CA, USA). Normal distribution and homogeneity of variance were assessed for each dataset. A t-test was used for comparisons between two groups, whereas one-way or two-way analysis of variance (ANOVA) followed by Tukey’s multiple comparison test was adopted for comparisons among multiple groups. A value of p < 0.05 was considered statistically significant.
Results
Identification of HucMSCs and EVs
We verified the multilineage differentiation potential of HucMSCs through osteogenic and adipogenic differentiation experiments (Fig. 1A). Flow cytometry analysis confirmed that HucMSCs were positive for surface markers CD90 and CD105, but negative for CD34 (Fig. 1B). EVs were successfully isolated via ultracentrifugation, and TEM revealed the typical cup-shaped morphology of EVs (Fig. 1C). NTA determined the average EV diameter to be 111.8 ± 30.7 nm, with a particle concentration of 1.66 × 109 particles/mL (Fig. 1D), aligning with standard EV characteristics. Western blot analysis further confirmed the presence of EV marker proteins CD63 and CD81 and the absence of CD45 (Fig. 1E). These results confirm that HucMSCs possess multilineage differentiation capacity, and EVs were successfully isolated.
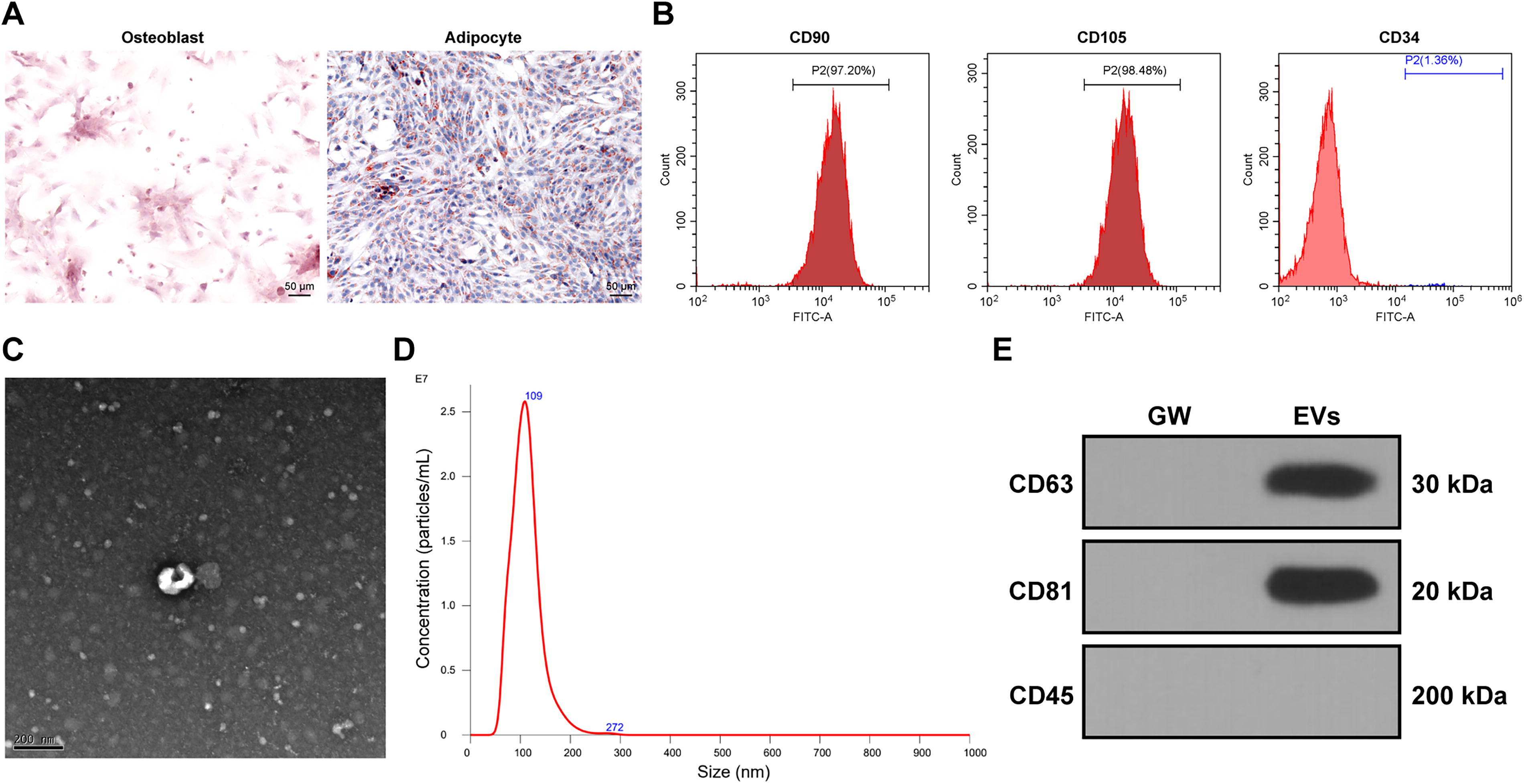
Identification of HucMSCs and EVs.
HucMSC-EVs promote HSF proliferation, migration, and collagen deposition under high-glucose conditions
To assess the impact of HucMSC-EVs on HSF proliferation and migration, HSFs were treated with varying concentrations of
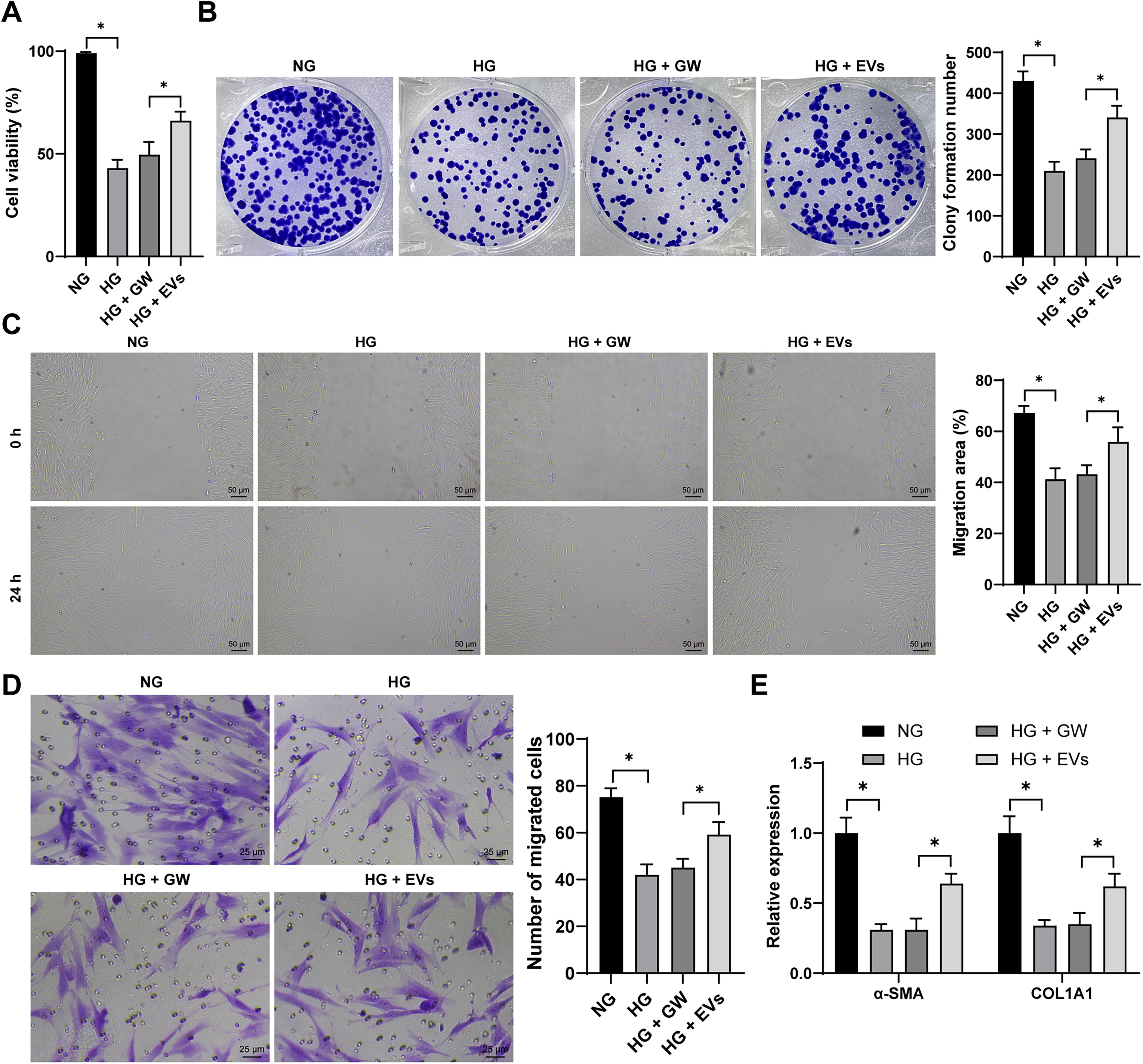
HucMSC-EVs promote the proliferation, migration, and collagen deposition of HSFs induced by high glucose. HSFs were supplemented with 30 mM
HucMSC-EVs deliver miR-21-5p into HSFs
Previous studies have shown that miR-21-5p plays a positive role in diabetic wound healing (Madhyastha et al., 2012). Based on this, we hypothesized that EVs may promote cell proliferation and migration by delivering miR-21-5p into cells. After treating the isolated EVs with RNase I, miR-21-5p expression remained largely unchanged. However, when treated with both RNase I and Triton X-100, miR-21-5p expression was significantly reduced (p < 0.05, Fig. 3A), suggesting that miR-21-5p is encapsulated within EVs. Furthermore, PKH67-labeled EVs were taken up by HSFs after coincubation (Fig. 3B), leading to a significant increase in miR-21-5p expression in HSFs (p < 0.05, Fig. 3C). When HucMSCs transfected with Cy3 (red fluorescence)-labeled miR-21-5p were coincubated with GFP-labeled HSFs, a substantial amount of red fluorescence was observed in HSFs. This red fluorescence was absent when the EV secretion inhibitor GW4869 was applied (Fig. 3D). These findings demonstrate that HucMSC-EVs can successfully transport miR-21-5p into HSFs, potentially offering protective effects in DFUs.
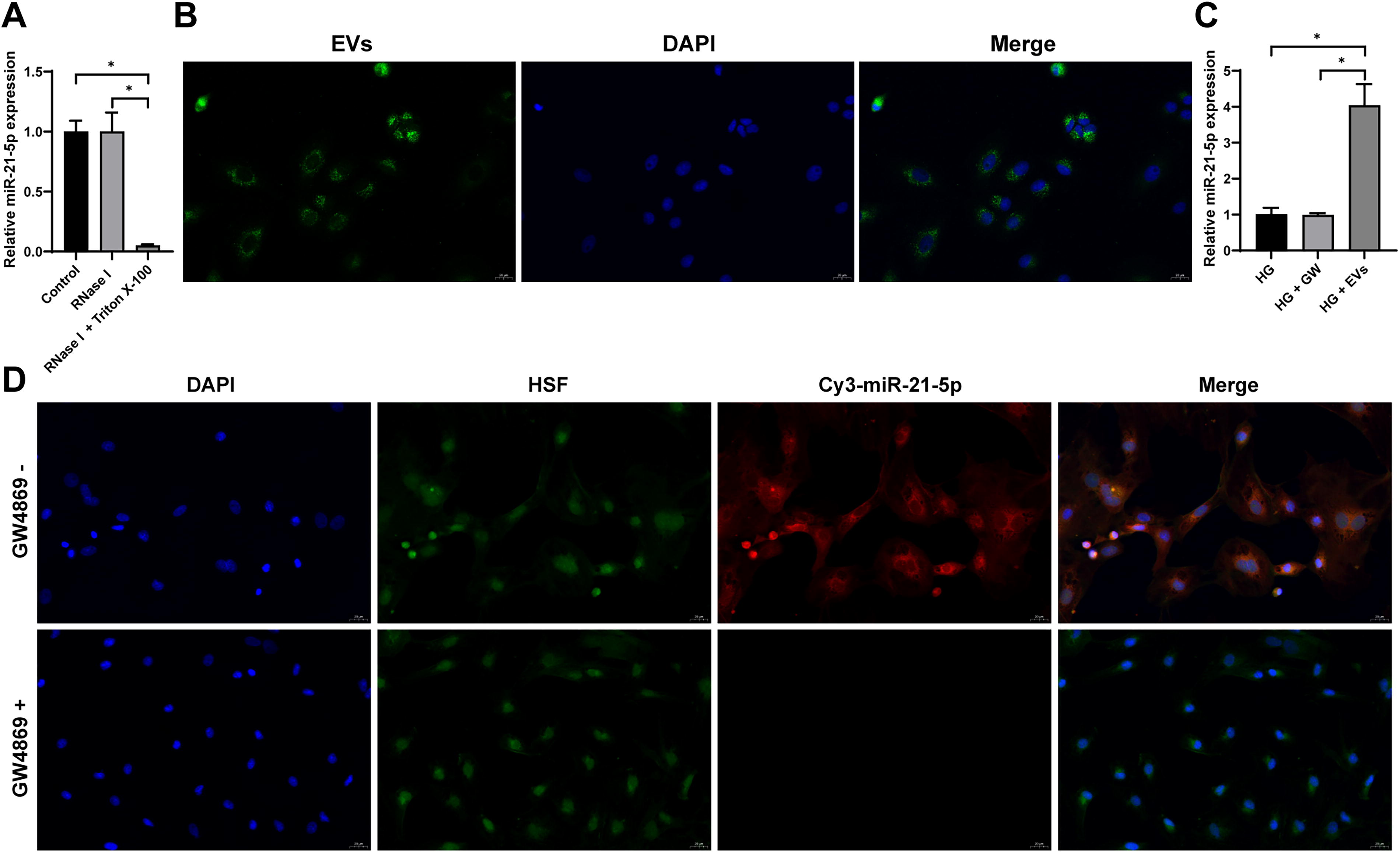
HucMSC-EVs carry miR-21-5p into HSFs.
HucMSC-EVs deliver miR-21-5p to promote HSF proliferation and migration
To further investigate the role of miR-21-5p, we overexpressed miR-21-5p in HucMSCs (p < 0.05, Fig. 4A) and subsequently isolated EV-miR-21-5p. miR-21-5p expression was significantly elevated in these EVs (p < 0.05, Fig. 4B). Upon incubation of EV-miR-21-5p with high glucose-induced HSFs, miR-21-5p levels in HSFs were significantly increased (p < 0.05, Fig. 4C). The CCK-8 and colony formation assay showed improved cell viability and proliferation in HSFs treated with EV-miR-21-5p (p < 0.05, Fig. 4D, E). In addition, scratch and Transwell assays demonstrated enhanced cell migration (p < 0.05, Fig. 4F, G). Overexpression of miR-21-5p also resulted in significant upregulation of α-SMA and COL1A1 expression (p < 0.05, Fig. 4H). Collectively, these results suggest that HucMSC-EVs promote the proliferation and migration of HSFs through the delivery of miR-21-5p.
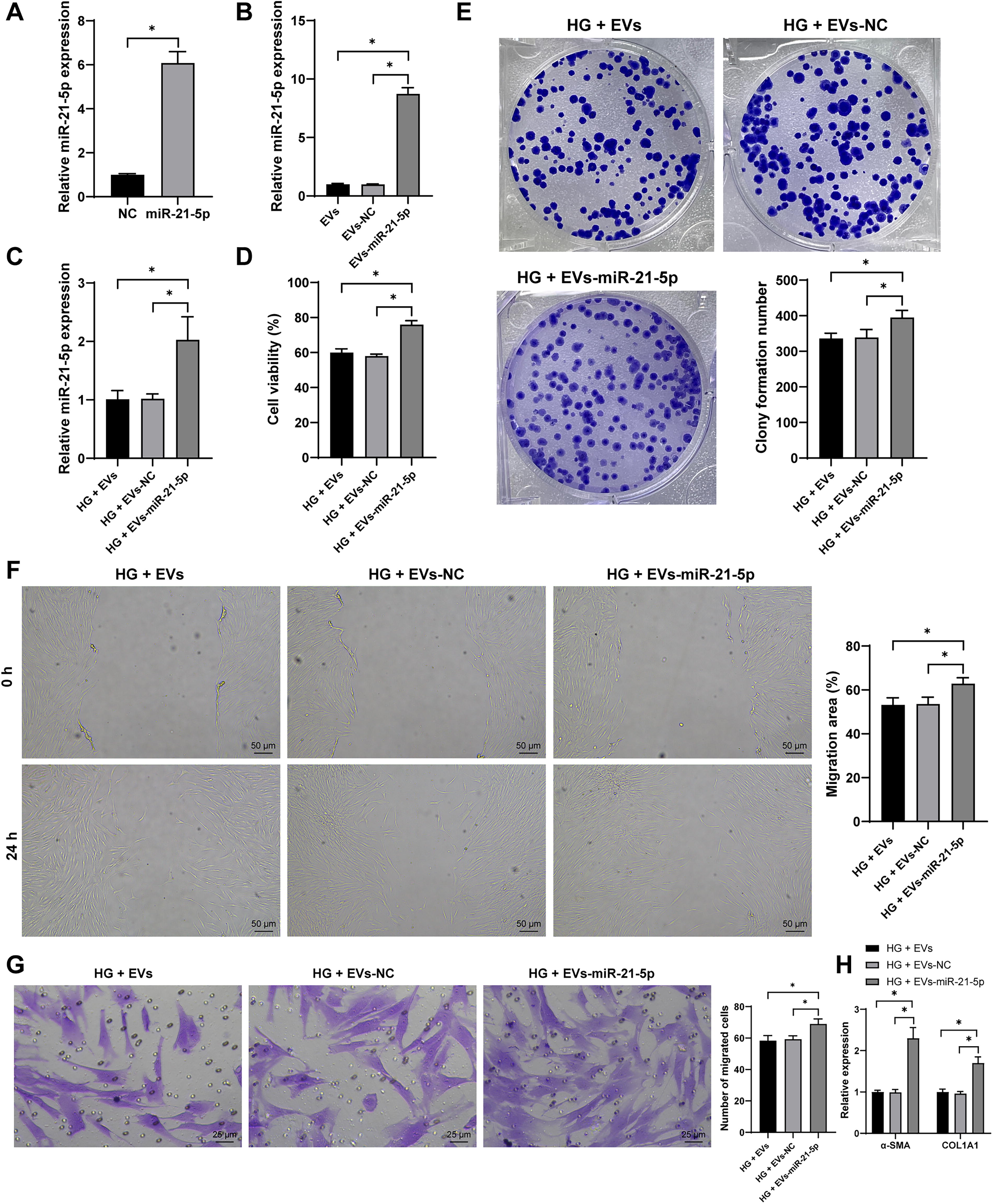
HucMSC-EVs carry miR-21-5p into HSFs to promote HSF proliferation, migration, and collagen deposition. HucMSCs were transfected with miR-21-5p mimic (miR-21-5p), with mimic NC (NC) as control. Then, EVs were isolated (EVs-miR-21-5p, EVs-NC) and incubated with HSFs under high-glucose conditions.
miR-21-5p targets KLF6 expression
Using the starBase, miRDB, and miRWalk databases, we predicted the downstream targets of miR-21-5p and created a Venn diagram to highlight the shared targets (Fig. 5A). Among these, KLF6 was identified as a key candidate. KLF6 has been shown to repress human keratinocyte proliferation and migration, which impedes diabetic foot wound healing (Zhao et al., 2023). Based on this, we hypothesized that HucMSC-EVs might carry miR-21-5p into HSFs, influencing their proliferation and migration by targeting KLF6. A dual-luciferase reporter assay confirmed the direct interaction between miR-21-5p and KLF6 (p < 0.05, Fig. 5B, C). Treatment with HucMSC-EVs led to a decrease in KLF6 expression, which was further reduced upon treatment with EV-miR-21-5p (p < 0.05, Fig. 5D, E). These results suggest that HucMSC-EVs deliver miR-21-5p into HSFs to suppress KLF6 expression.
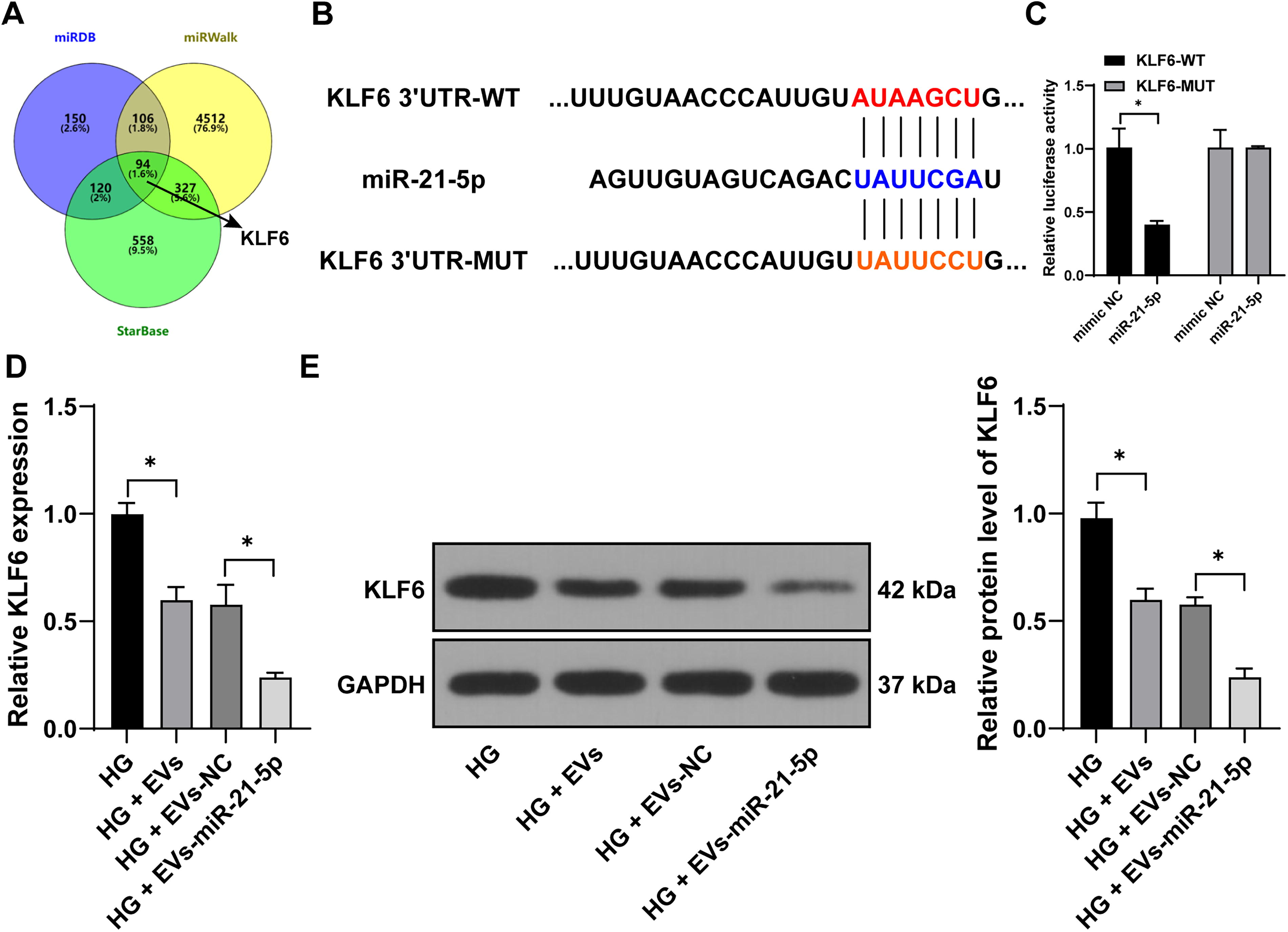
miR-21-5p targets KLF6 expression.
Overexpression of KLF6 reverses the effect of EV-miR-21-5p on HSF proliferation, migration, and collagen deposition
To further validate the mechanism, we overexpressed KLF6 in HSFs (p < 0.05, Fig. 6A–C) and then conducted experiments combining KLF6 overexpression with EV-miR-21-5p treatment. The overexpression of KLF6 significantly attenuated the positive effects of EV-miR-21-5p on cell proliferation and migration (p < 0.05, Fig. 6D–G). In addition, KLF6 overexpression notably downregulated the expression of α-SMA and COL1A1 (p < 0.05, Fig. 6H). These findings indicate that KLF6 overexpression partially reverses the effects of EV-miR-21-5p on HSF proliferation, migration, and collagen deposition.
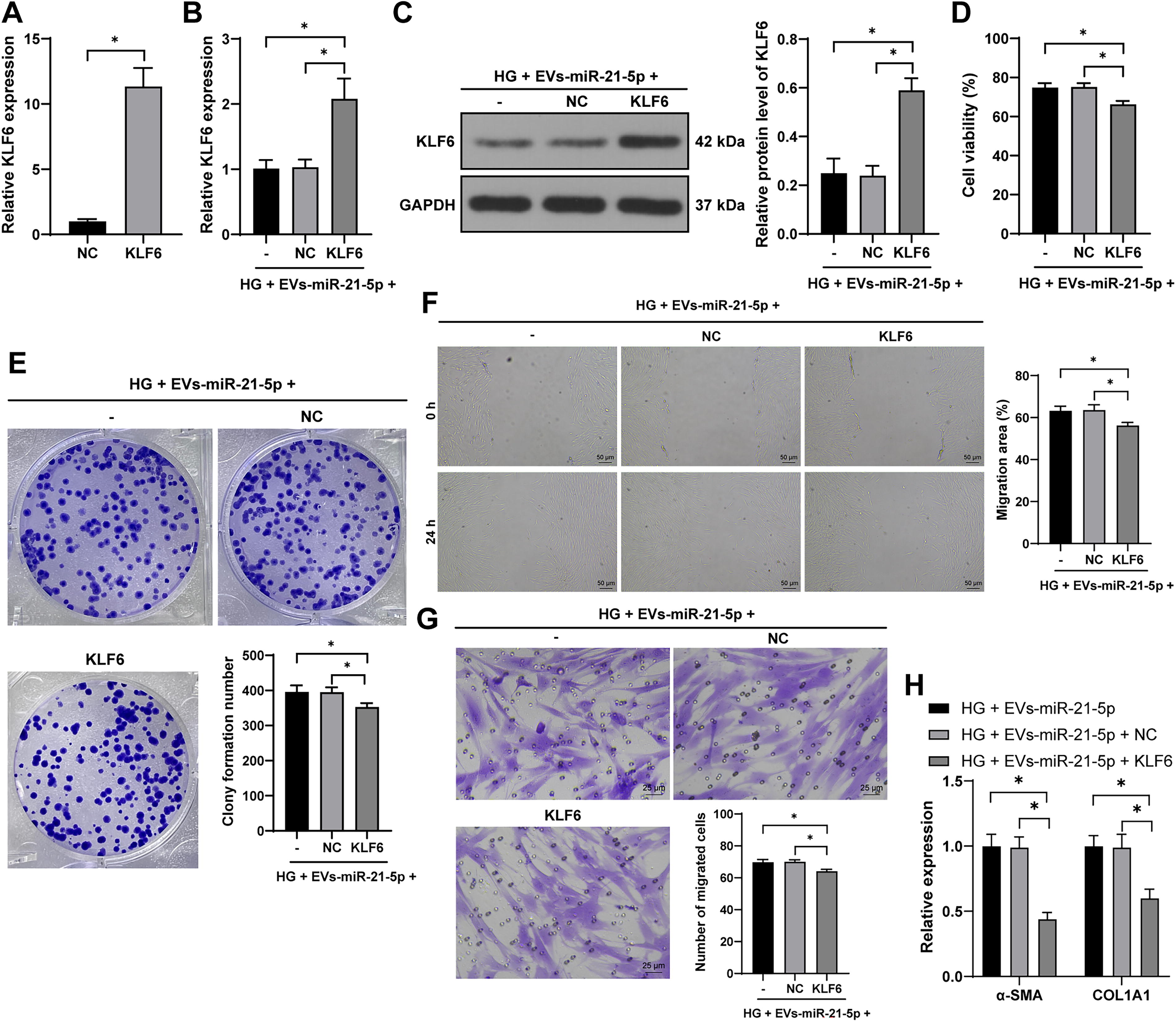
Overexpression of KLF6 reverses the promoting effect of EV-miR-21-5p on HSF proliferation, migration, and collagen deposition. HSFs were transfected with KLF6 overexpression plasmid (KLF6), with empty vector as control (NC).
Discussion
The hyperglycemic environment and various biological factors associated with diabetes significantly impair wound healing, often resulting in chronic ulcers (Everett and Mathioudakis, 2018). Recently, EVs derived from MSCs have gained attention in cell-free regenerative medicine, particularly for their potential in treating DFUs (An et al., 2021). In this study, we demonstrate that HucMSC-EVs, which carry miR-21-5p, promote the proliferation and migration of HSFs in DFU by targeting KLF6.
The ability of MSCs to differentiate into multiple lineages makes them highly promising candidates in tissue engineering, regenerative medicine, and clinical therapy. Our findings confirmed the multilineage differentiation potential of HucMSCs through osteogenic and adipogenic differentiation experiments, and we successfully isolated EVs from these cells. MSC-EVs hold significant therapeutic potential in skin regeneration due to their anti-inflammatory, antioxidant, and wound-healing properties (Gui et al., 2024). Previous studies have highlighted the beneficial role of MSC-EVs in wound healing by key signaling pathways and promoting the release of growth factors (Shabbir et al., 2015). In addition, MSC-EVs have been shown to support skin rejuvenation and counteract skin aging (Gui et al., 2024), enhance the proliferation of human dermal fibroblast in vitro (Zhang et al., 2018), and promote scar-free wound healing in vivo (Jiang et al., 2020).
In our study, we explored the effects of HucMSC-EVs on HSF proliferation and migration under high-glucose conditions. We observed that high glucose reduced HSF viability, proliferation, and migration, whereas EV treatment successfully reversed these impairments. This suggests that HucMSC-EVs can enhance HSF proliferation and migration, even in the challenging environment induced by high-glucose levels.
EVs are known to shuttle various genetic materials, including miRNAs, enabling them to mediate cell-to-cell communication and regulate molecular activities in recipient cells (Thery et al., 2018). miRNAs transported by EVs have been shown to enhance diabetic wound healing by suppressing inflammation and promoting revascularization, epithelialization, and extracellular matrix remodeling (Shen et al., 2022). Among these, miR-21-5p stands out as one of the most differentially regulated miRNAs in DFU fibroblasts (Liang et al., 2016). We hypothesized that EVs could promote HSF proliferation and migration by delivering miR-21-5p into HSFs. Our findings confirmed that miR-21-5p was encapsulated in EVs and transferred into HSFs by HucMSC-EVs. Previous studies have demonstrated that bone marrow stromal cells (BMSCs)-derived exosomes can enhance wound healing by upregulating miR-21-5p, which improves angiogenesis and fibroblast functions (Wu et al., 2020). The role of miR-21-5p in promoting fibroblast viability and migration during wound healing has also been well-documented (Xiao et al., 2021). In our study, we further validated this by overexpressing miR-21-5p in HucMSCs and isolating EV-miR-21-5p. When these EVs were coincubated with high glucose-induced HSFs, we observed a marked improvement in cell viability, proliferation, and migration. These findings suggest that the beneficial effects of HucMSC-EVs on HSF proliferation and migration are mediated by miR-21-5p. Similarly, miR-21-5p has been shown to enhance regenerative wound healing through TGF-β receptor inhibition in umbilical cord blood-derived MSCs (Zhang et al., 2021), and BMSC-derived EVs upregulate miR-21-5p to improve angiogenesis and fibroblast function in wound healing (Wu et al., 2020).
Next, we aimed to identify the downstream targets of miR-21-5p. Using online prediction databases, we focused on KLF6, a DNA-binding protein with a triple zinc-finger motif, which is involved in various diabetic complications such as diabetic retinopathy (Cao et al., 2022), diabetic kidney injury (Horne et al., 2018), and diabetic pulmonary fibrosis (Zou et al., 2017). KLF6 has also been implicated in impaired wound healing in DFUs, with its upregulation diminishing the ability of keratinocytes to counteract high glucose-induced damage (Zhao et al., 2023). Notably, miR-21-5p carried by adipose mesenchymal stem cell-derived apoptotic bodies has been reported to target KLF6, inducing M2 macrophage polarization and enhancing skin wound healing (Li et al., 2022). Based on these findings, we speculated that miR-21-5p carried by HucMSC-EVs promotes HSF proliferation and migration by targeting KLF6. Our dual-luciferase reporter gene assay confirmed the binding between miR-21-5p and KLF6, and subsequent functional rescue experiment showed that KLF6 overexpression reversed the positive effects of miR-21-5p-enriched EVs on HSF proliferation and migration.
In summary, HucMSC-EVs deliver miR-21-5p into HSFs, targeting KLF6 expression and thereby enhancing HSF proliferation and migration, which could play a significant protective role in promoting wound healing in DFUs. However, this study was limited to in vitro experiments, and the mechanism has not yet been validated in vivo. To fully elucidate the therapeutic potential of HucMSC-EVs in DFU, further animal studies are essential. In addition, the role of KLF6 as a transcription factor requires more in-depth investigation to understand its broader regulatory effects. Future research will integrate both in vivo and in vitro approaches to further explore the protective mechanisms and therapeutic efficacy of HucMSC-EVs in DFU treatment.
Footnotes
Authors’ Contributions
J.Z. conceived and designed the study and drafted the article. Y.G. performed the experiments and analyzed the data. P.H. supervised the project and critically reviewed the article.
Author Disclosure Statement
The authors declare no conflicting financial interests.
Funding Information
No funding was received for this article.