
Abstract
From the first cloning of animals—salamanders—to the cloning of primates—monkeys—nuclear transfer research has spanned an extensive 96-year history. Over the course of nearly a century, it has addressed fundamental scientific questions and found applications across a wide range of practical fields. This review provides a comprehensive overview of the key milestones in its development, its practical applications, and the challenges it continues to face.
Introduction
The successful cloning of monkeys from somatic cells in 2018 reignited global interest in animal cloning, thrusting the topic back into the spotlight—much like the cloning of Dolly the sheep did decades earlier. This landmark achievement sparked widespread discussion and renewed an essential question: what precisely is animal cloning?
Cloning refers to a form of asexual reproduction that does not involve gametes but instead produces new individuals directly through the division of a parent organism. This mode of reproduction is common among unicellular organisms and simpler multicellular organisms with less specialized structures and functions. In contrast, organisms with greater structural and functional complexity—such as vertebrates and other bilaterian animals—typically reproduce sexually. In sexual reproduction, fertilized eggs result from the fusion of male and female gametes, facilitating extensive genetic variation and enhancing the adaptability of offspring to natural selection. Given this, why would we choose to revert from a more straightforward and adaptable form of sexual reproduction to asexual reproduction through a complex scientific methodology? Furthermore, how has this been achieved?
Owing to the intricate nature of sexual reproduction, scientists have leveraged the genetic uniformity inherent in asexual reproduction to explore and address a series of fundamental scientific questions. This approach is achieved through a technique known as nuclear transfer. As the term implies, nuclear transfer involves the transplantation of a nucleus from one cell into another. While the term “cloning” is widely used in popular discourse, “nuclear transfer” is the precise technical terminology.
To provide readers with a comprehensive understanding of nuclear transfer, this article reviews the key scientific milestones in its historical development, highlighting the breakthroughs, applications, and challenges associated with this groundbreaking technique.
Research on artificially created twin animals: A theoretical and experimental foundation for nuclear transfer
Sexually reproducing animals develop from a single-celled fertilized egg formed by the combination of male and female gametes. However, does the embryonic blastomere, formed after the fertilized egg undergoes division, still retain this developmental ability? The following experimental studies on artificially created twin animals provide a scientific answer.
The First Case of Artificial Twin Animals—Sea Urchins
In 1885, Hans Driesch carried out pioneering experiments by separating 2-cell stage embryos of sea urchins, members of the phylum Echinodermata (invertebrates). He observed that each isolated blastomere had the capacity to develop into a fully independent individual (Chanoine, 2022). This remarkable discovery marked the first documented case of artificially produced twin animals in scientific history, as shown in Figure 1.

Driesch
The First Case of Artificial Twin Vertebrates—Salamanders
In 1903, Hans Spemann built upon the experimental work initiated with sea urchins by applying a fine hair to ligate 2-cell stage salamander embryos, thereby separating them into two independent blastomeres. His experiments demonstrated that each blastomere retained the ability to develop into a complete and independent individual (De Robertis, 2006). This breakthrough marked the first instance of artificially created twin vertebrates, as shown in Figure 2.

Spemann
Furthermore, Spemann extended his studies by attempting to isolate blastomeres from embryos at later developmental stages. He discovered that these later-stage cells had lost their capacity to develop into independent individuals, indicating a progressive restriction in developmental potential as embryonic differentiation proceeds.
These two artificial twin experiments definitively addressed a fundamental scientific question: at the 2-cell stage, each blastomere is totipotent, possessing the full developmental potential of a fertilized egg to give rise to a complete organism. However, this totipotency progressively diminishes as the embryo advances through subsequent developmental stages.
Significant Developments in Nuclear Transfer: Blastomere Nuclear Transfer and Somatic Cell Nuclear Transfer
Blastomere Nuclear Transfer
The first case of nuclear transfer in animals—Salamanders
In 1928, building upon his previous artificial twin experiments, Spemann conducted the first nuclear transfer experiment using salamanders. He partially ligated a fertilized salamander egg, creating one half containing only cytoplasm and the other half containing the nucleus. When the nucleated half developed to the 16-cell stage, Spemann removed the ligature and inserted a cell nucleus from a blastomere into the previously non-nucleated cytoplasm, creating a reconstructed embryo. He then separated this embryo and found that it could develop into an independent organism (De Robertis, 2006), as shown in Figure 3.
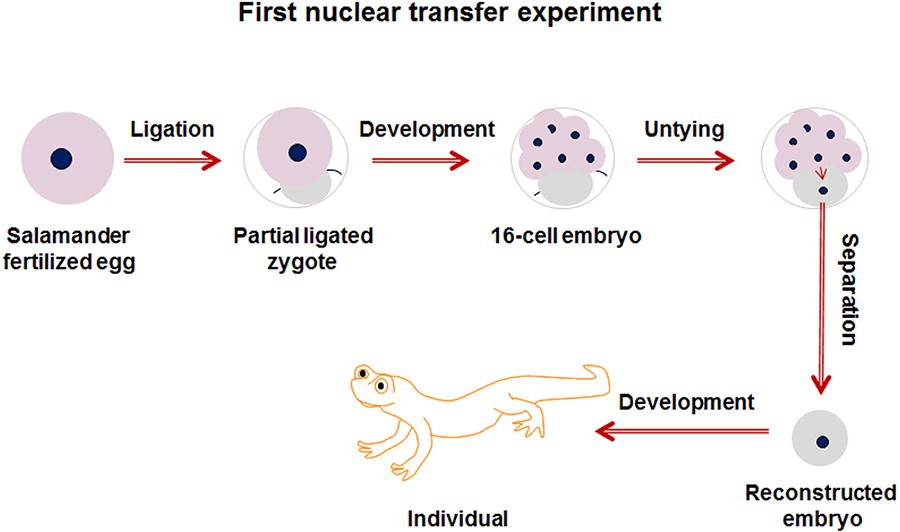
The process of Spemann’s nuclear transfer.
This was the first nuclear transfer experiment in human history, and it addressed the following key scientific questions: (1) embryonic cells, under the influence of the fertilized egg cytoplasm, can complete the development of an entire organism and (2) the embryonic cell nucleus directs the process of embryonic development, rather than the cytoplasm.
Furthermore, Spemann hypothesized that if later-stage embryonic cells or even cells from a fully developed individual were used for nuclear transfer, it might be possible to produce a new individual. For these groundbreaking contributions, Spemann was awarded the Nobel Prize in Physiology or Medicine in 1935.
The first case of nuclear transfer using oocyte induction—Leopard frogs
In 1952, Robert Briggs and Thomas Joseph King conducted the first reported nuclear transfer experiment using oocytes as recipients. Their method represented a significant advancement on Spemann’s work, developing a nuclear transfer procedure that is still used today (as shown in Fig. 4). The procedure involved removing the nucleus from a leopard frog (Rana pipiens) oocyte and injecting the nucleus from a separated embryo cell of the same species into the enucleated oocyte to create a reconstructed embryo. They found that this reconstructed embryo could continue to develop into an independent organism (Briggs and King, 1952), although the efficiency was quite low (similar to Spemann’s nuclear transfer experiments with salamanders), particularly when using nuclei from later-stage embryos as donors.
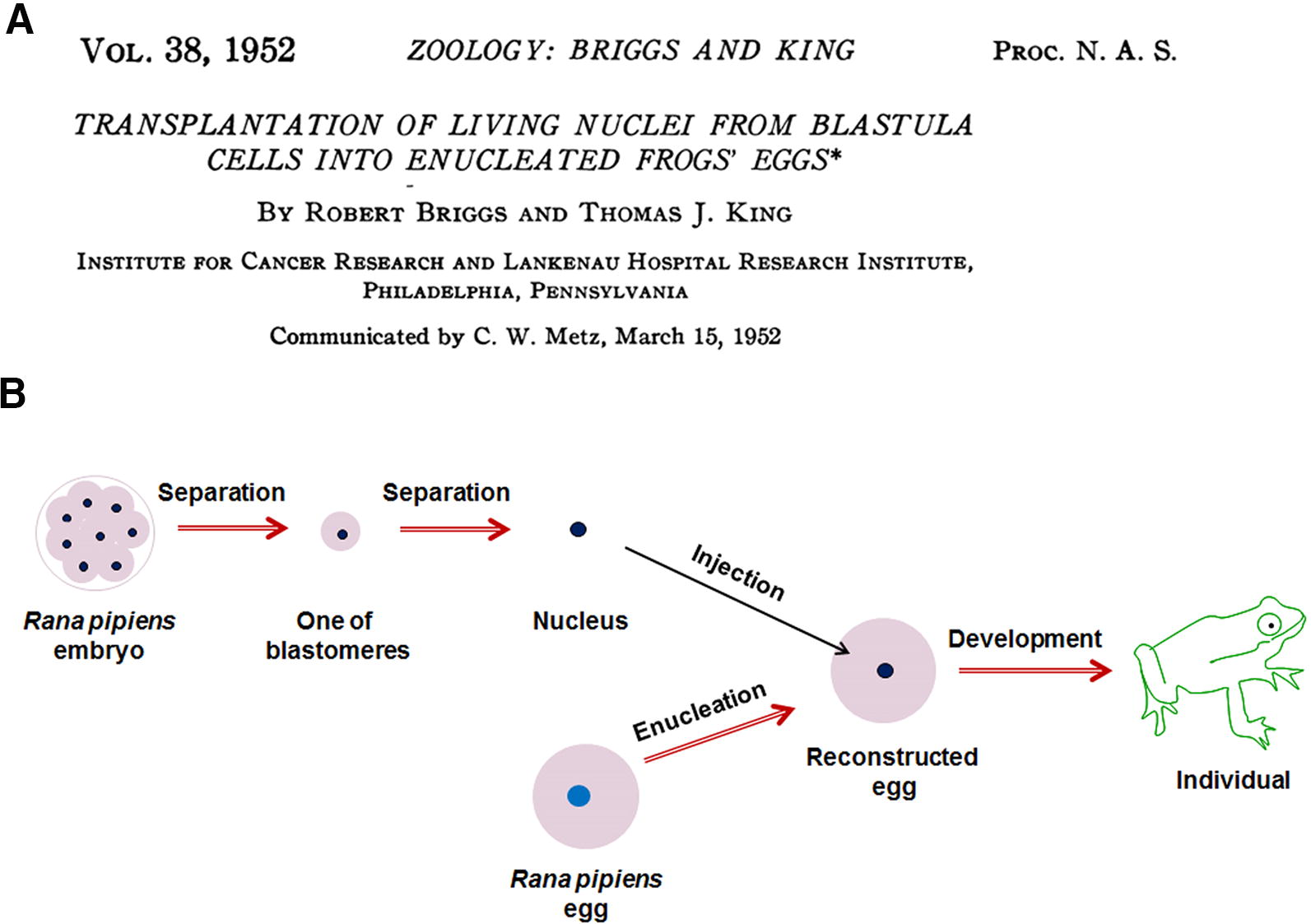
The nuclear transfer study with frog embryos by Robert Briggs and Thomas Joseph King, reported in The Proceedings of the National Academy of Sciences in 1952
This experiment addressed several critical scientific questions: (1) the cell nucleus directly guides cellular growth and individual development and (2) Early-stage embryonic cells are more effective for nuclear transfer than those from late-stage embryos.
Furthermore, in this study, they conducted a fascinating experiment in which they transferred an embryo cell nucleus from a bullfrog (Rana catesbeiana) into an enucleated oocyte of a leopard frog. They discovered that the reconstructed embryo could develop up to the blastocyst stage before halting and dying. This was likely the first interspecies nuclear transfer experiment in human history.
The first case of nuclear transfer in mammalian embryos
Due to the external fertilization of amphibians and their relatively large oocytes, external experimental manipulations were comparatively straightforward. However, in mammals, which fertilize internally, the oocyte is significantly smaller than that of amphibians, with a size ratio of approximately 30–50:1, making in vitro manipulation far more challenging. In 1975, J. Derek Bromhall developed a nuclear transfer procedure for mammals. Using a finely drawn and flame-polished glass needle, he injected the nucleus from a rabbit embryo into an enucleated oocyte. He found that the reconstructed embryo could develop to the morula stage, marking the first documented case of nuclear transfer in a mammalian embryo (Bromhall, 1975), as shown in Figure 5.
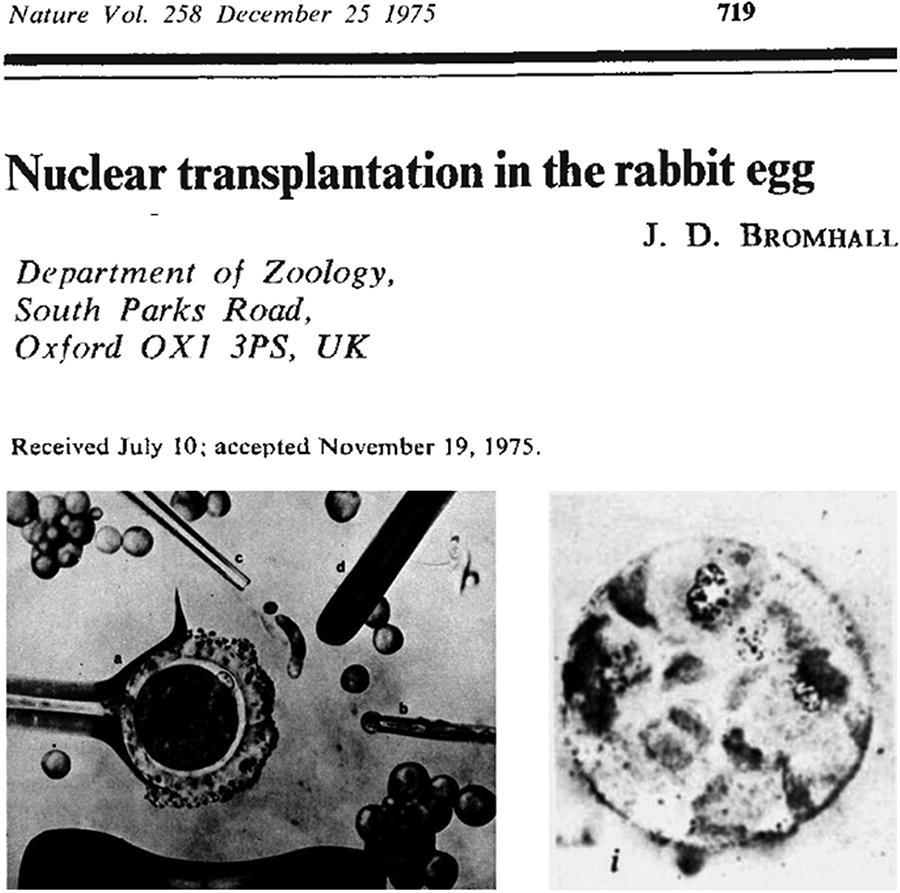
J. Derek Bromhall’s independent study on rabbit nuclear transfer, published in Nature in 1975. The lower left illustrates the nuclear transfer process in a rabbit oocyte, while the lower right shows a cloned rabbit embryo at the morula stage.
This research also suggested the possibility that nuclear transfer in mammals could eventually lead to the development of a fully formed organism.
The first mammal produced by nuclear transfer—Sheep
In 1986, Steen Willadsen successfully performed the first nuclear transfer experiment in mammals using sheep. He separated a blastomere from an 8-cell stage sheep embryo and fused it with an enucleated oocyte using electrical fusion to create a reconstructed embryo. He found that this newly reconstructed embryo could continue to develop in vitro and successfully resulted in the birth of three live lambs after implantation into a surrogate uterus (Willadsen, 1986), as shown in Figure 6.

Steen Willadsen’s independent publication in Nature in 1986 on the production of cloned sheep using sheep embryo cell nuclear transfer.
This study marked the first instance in human history where a mammal was produced through nuclear transfer, demonstrating the feasibility of this technique in mammals. Although early-stage embryonic cells were used as nuclear donors, this experiment laid the foundation for later developments in somatic cell nuclear transfer (SCNT) in mammals.
The first primate produced by nuclear transfer—Rhesus monkey
In 1997, Li Meng, Don Wolf, and their team applied the nuclear transfer method used in sheep to conduct the first successful nuclear transfer experiment in primates. They reconstructed embryos by fusing a blastomere from a rhesus monkey embryo with an enucleated oocyte and observed that these embryos developed normally in vitro. After implantation into a surrogate monkey’s uterus, two healthy rhesus monkeys were born (Meng et al., 1997), as shown in Figure 7.

The successful 1997 cloning of two rhesus monkeys by Li Meng and Don Wolf’s team using embryonic cell nuclear transfer.
This was the first documented case of nuclear transfer in a primate, providing crucial experimental evidence for somatic cell cloning in monkeys.
SCNT
The first case of SCNT in animals—African clawed frog
In 1958, John Gurdon built upon earlier nuclear transfer experiments with frog embryo cells by conducting a groundbreaking study using the African clawed frog (Xenopus laevis). He transferred the nucleus from an intestinal cell of a juvenile frog into an enucleated oocyte and found that the resulting embryo was capable of developing into an independent organism, with a genome identical to that of the donor cell (Gurdon et al., 1958), as shown in Figure 8. This marked the first successful case of SCNT in animals.

John Gurdon
This experiment addressed the following key scientific questions: (1) A differentiated cell can, under certain conditions, revert to an earlier state and redevelop into a complete organism; (2) A single cell retains all the genetic material required for development, even after undergoing division and differentiation. For this groundbreaking discovery, John Gurdon was awarded the Nobel Prize in Physiology or Medicine in 2012.
The first mammal cloned by SCNT—Dolly the sheep
In 1997, Ian Wilmut, Keith Campbell, and their team reported the first successful cloning of a mammal using SCNT. Building on earlier nuclear transfer experiments with sheep embryos, they used mammary epithelial cells from an adult Finn Dorset sheep and enucleated oocytes from a Scottish Blackface sheep. These cells were fused using electrical stimulation to create reconstructed embryos, which successfully developed to the morula and blastocyst stages in vitro. After transferring the embryos into a surrogate sheep’s uterus, one live lamb was born (Wilmut et al., 1997), as shown in Figure 9.

Ian Wilmut and Keith Campbell’s Nature cover article in 1997, reporting the cloning of Dolly the sheep using the mammary gland cells of an adult sheep.
This sheep, famously named Dolly, became the first mammal in history to be cloned from a somatic cell, representing a groundbreaking achievement with profound scientific and societal implications. Dolly quickly became a household name and an enduring symbol of cloning research.
The first genetically modified mammal cloned via SCNT—Polly the sheep
In 1997, Angelika Schnieke, Keith Campbell, Ian Wilmut, and their team achieved a groundbreaking milestone in reproductive biology by cloning a genetically modified sheep named Polly through SCNT. The researchers introduced the human coagulation factor IX gene, essential for blood clotting, into ovine fetal fibroblasts. These genetically modified cells were then used as nuclear donors in the SCNT process, ultimately leading to the creation of Polly (Fig. 10). This marked the first successful cloning of a genetically modified mammal using SCNT (Schnieke et al., 1997).

The first genetically modified mammal cloned via SCNT—Polly sheep. SCNT, somatic cell nuclear transfer.
This achievement underscored the transformative potential of SCNT for precise genetic modifications in animals, laying the foundation for future innovations in biomedical research, agriculture, and pharmaceuticals. Today, SCNT is used to generate genetically modified pigs for organ transplantation, mouse models for fundamental scientific studies, and livestock engineered for improved meat production or milk with enhanced nutritional properties.
The first cloned animal from a frozen deceased body—Mouse
In 2007, Teruhiko Wakayama, who had previously reported the first successful cloning of mice (Wakayama et al., 1998), announced the successful cloning of a mouse that had been preserved without any cryoprotectant at −20°C for 16 years (Wakayama et al., 2008). In this groundbreaking study, Wakayama and his team attempted nuclear transfer using cells from various tissues of the deceased mouse, finding that brain-derived cells achieved the highest efficiency for both in vitro embryo development and the generation of embryonic stem cells (ESCs). They subsequently used ESCs derived from brain cells as donor nuclei for successive rounds of nuclear transfer, ultimately producing four cloned mice, two of which exhibited normal reproductive capability, as shown in Figure 11.
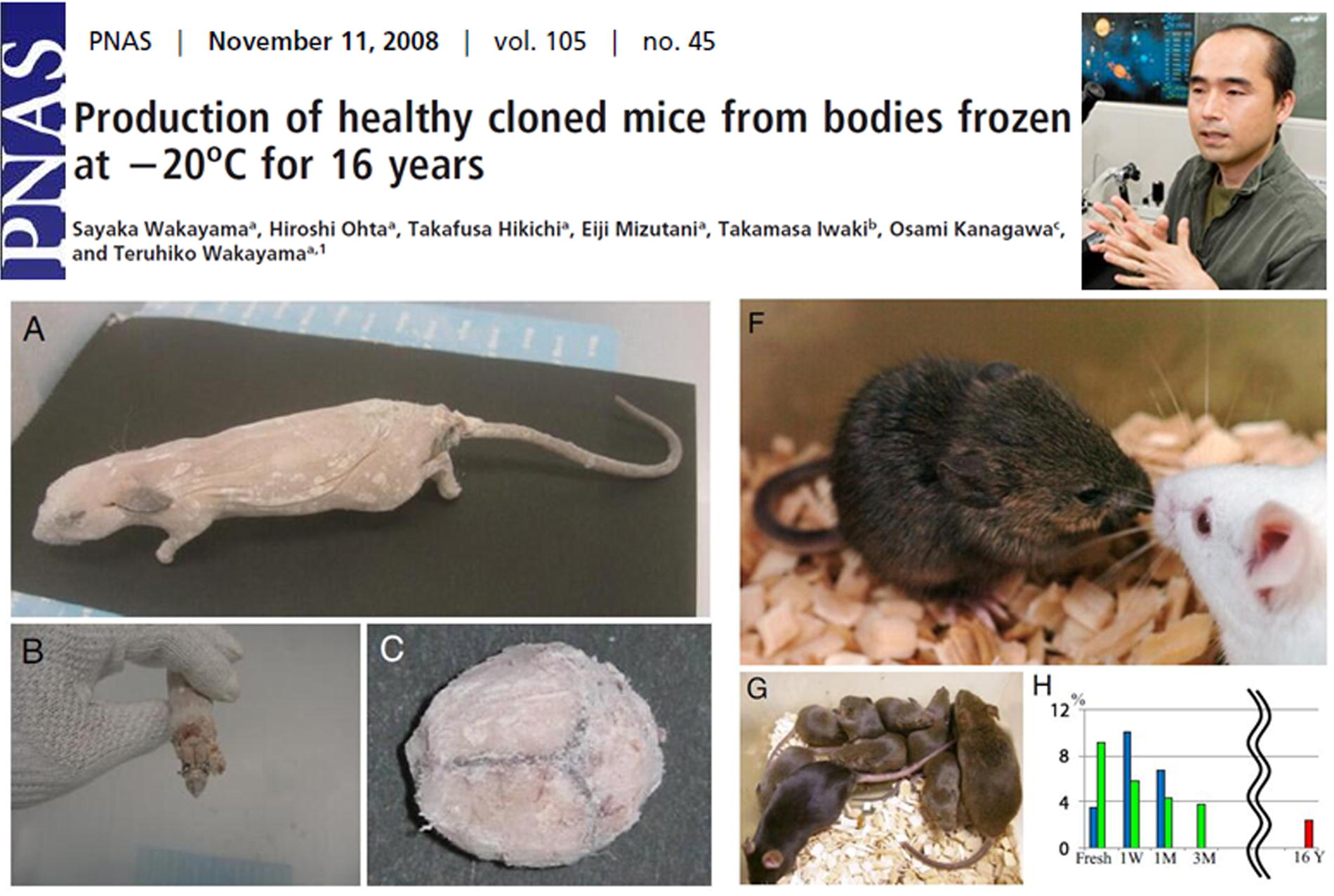
Teruhiko Wakayama et al. successfully cloned a mouse from a carcass that had been cryopreserved at −20°C for 16 years without any cryoprotectants in 2007.
This pioneering research addressed several critical scientific questions: (1) even if all the cells in a long-preserved frozen body are dead, cloning is still possible as long as the genetic material remains intact; (2) it provides a novel method for preserving the entire genomes of species; and (3) it offers new insights and experimental foundations for the potential resurrection of extinct species.
The first cloned primate by SCNT—Long-tailed macaques
In 2018, Qiang Sun, Zhen Liu, and their team successfully reported the first-ever cloned primates using SCNT, employing fetal fibroblast nuclei from the skin of crab-eating macaques. This resulted in the birth of two healthy cloned individuals (Liu et al., 2018), as shown in Figure 12. This achievement marks the first successful cloning of a primate species using SCNT in human history.

A Cell cover story in February 2018, reported the successful cloning of two rhesus monkeys by Sun Qiang and Liu Zhen from the Chinese Academy of Sciences, using fetal skin fibroblasts.
Since nonhuman primates are regarded as the most relevant models for studying human diseases, especially neurological or psychiatric disorders, due to their closer genetic similarity to humans, the use of cloning, which ensures genetic uniformity, eliminates the individual differences inherent in traditional model organisms. Therefore, this research holds significant scientific value for biomedical studies and disease modeling.
First genetically modified cloned primate by SCNT—Macaque monkey
In 2019, Liu Zhen, Sun Qiang, Hung-Chun Chang, and their team achieved a significant milestone by applying SCNT in combination with CRISPR/Cas9 gene-editing technology to generate genetically modified macaque monkeys. They used fibroblasts from a young adult macaque, in which the core circadian transcription factor BMAL1 was knocked out using the CRISPR/Cas9 system, to conduct SCNT. This method led to the successful cloning of five macaque monkeys carrying homozygous BMAL1 mutations without mosaicism (Fig. 13, Liu et al., 2019).
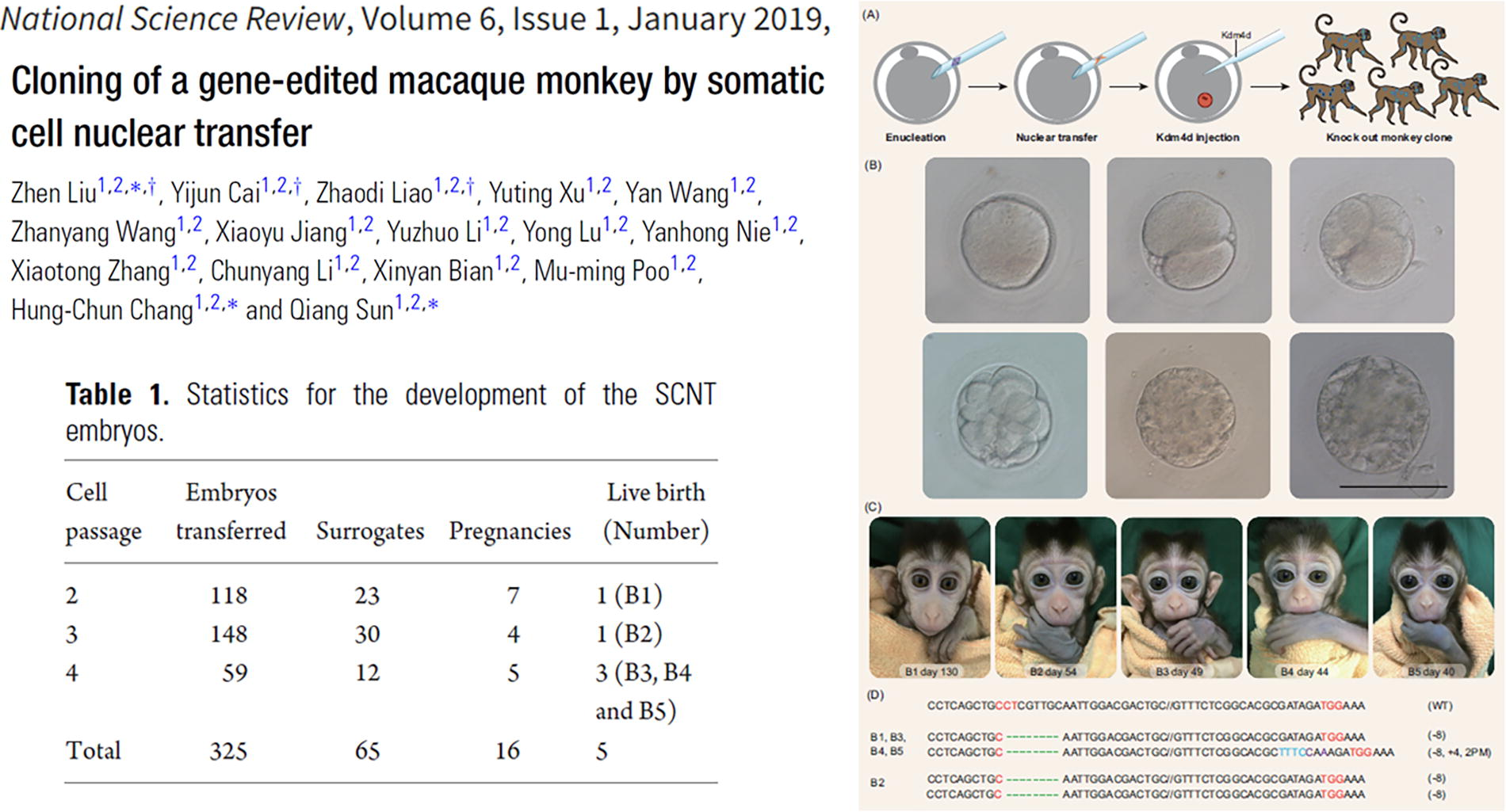
First genetically modified cloned primate by SCNT—macaque monkey.
This groundbreaking study highlighted the power of combining SCNT with precise gene-editing technologies, establishing a reliable method for creating genetically modified nonhuman primates with stable and targeted genetic modifications. The approach offers a valuable tool for generating uniform disease models in macaque monkeys, advancing both biological research and therapeutic innovation.
First organ transplant from a genetically modified cloned animal into a human (pig heart)
In 2022, Bartley Griffith, Muhammad Mohiuddin, and their team performed the first heart transplant from a genetically modified pig into a human patient. The pig was cloned via SCNT using fibroblasts that had been genetically edited with 10 specific modifications to enhance xenograft compatibility. Key modifications included the knockout of three xenoantigen carbohydrates (galactose-alpha-1,3-galactose, Sda blood group antigen, and N-glycolylneuraminic acid) to prevent hyperacute rejection, as well as the knockout of the growth hormone receptor to control graft size. Human genes were added to improve graft function, including CD46, decay-accelerating factor, thrombomodulin, and endothelial protein C receptor, among others. The recipient was a 57-year-old man with end-stage heart failure who was dependent on extracorporeal membrane oxygenation. Post-transplant, the pig heart functioned without rejection for 49 days. However, graft failure occurred due to unexplained diastolic thickening and edema, and life support was withdrawn on day 60. An autopsy revealed edema and myocyte necrosis, with no signs of conventional rejection. Further studies are ongoing to identify the underlying mechanisms (Fig. 14) (Griffith et al., 2022).
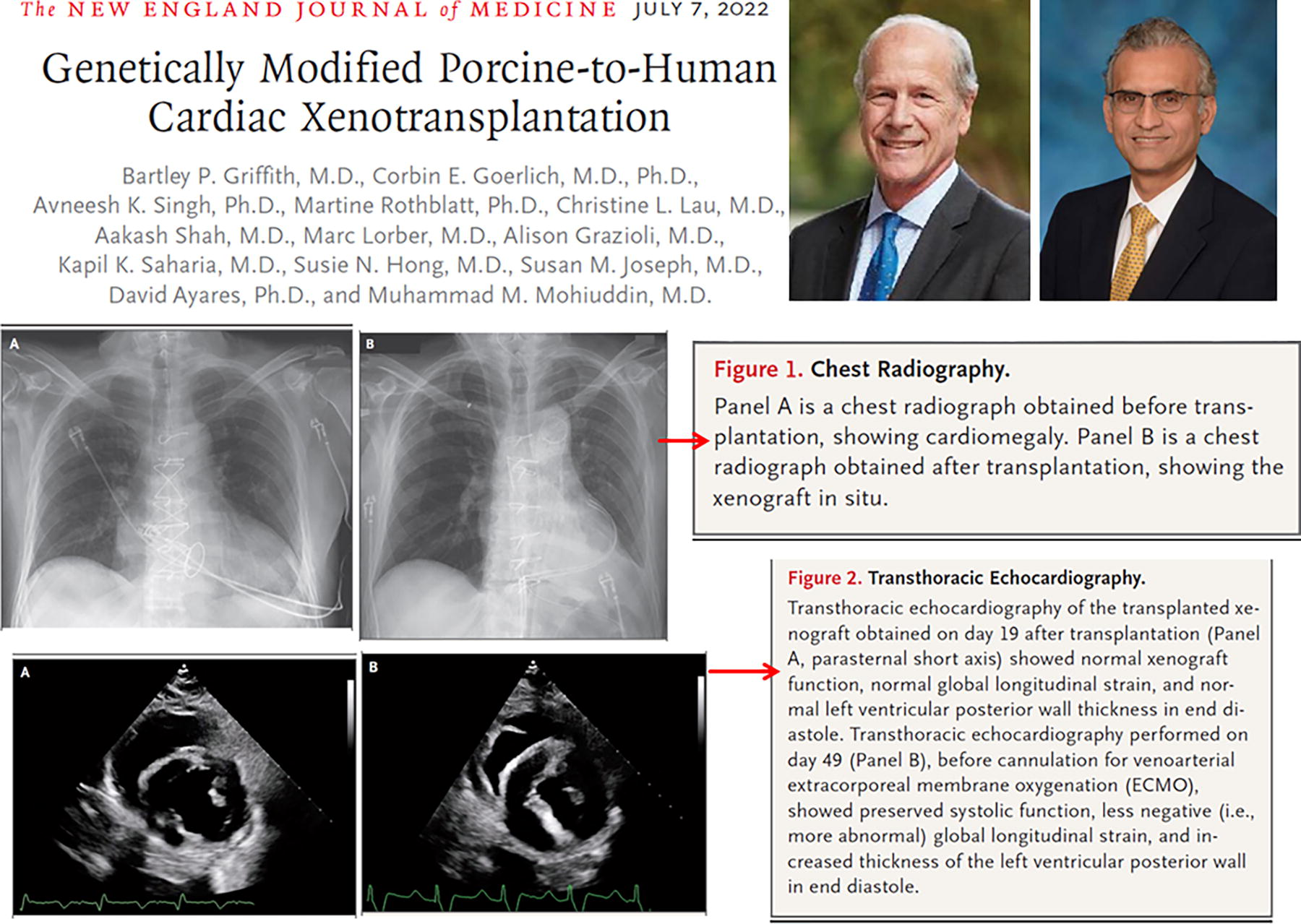
First organ transplant from a genetically modified cloned animal into a human (pig heart).
This landmark case demonstrated the feasibility of using genetically modified organs derived from SCNT for xenotransplantation. It set the stage for advancing formal, large-scale clinical trials, highlighting the potential to generate a new and sustainable supply of animal organs suitable for transplantation into human patients.
The club of first cloned mammal species by SCNT
Since the birth of Dolly the sheep, the feasibility of SCNT has been demonstrated in various other species. To date, the following 19 species have been successfully cloned and reported in formal international scientific journals as healthy, surviving mammals: Cattle (Cibelli et al., 1998), mice (Wakayama et al., 1998), goats (Baguisi et al., 1999), pigs (Polejaeva et al., 2000), mouflon sheep (Loi et al., 2001), rabbits (Chesné et al., 2002), cats (Shin et al., 2002), horses (Galli et al., 2003), rats (Zhou et al., 2003), mules (Woods et al., 2003), African wild cats (Gómez et al., 2004), dogs (Lee et al., 2005), ferrets (Li et al., 2006), wolves (Kim et al., 2007), water buffalo (Shi et al., 2007), red deer (Berg et al., 2007), dromedary camels (Wani et al., 2010, and nonhuman primates—long-tailed macaques (Liu et al., 2018) (Fig. 15).

The first cloned mammals derived from somatic cells (all animal photos are from original papers). From left to right, top to bottom: Dolly, cow, mouse, goat, pig, European mouflon, rabbit, domestic cat, African wildcat, rat, ferret, dog, wolf, buffalo, mule, red deer, dromedary, horse, and long-tailed macaque.
In addition, there are also two reported cases of abnormal interspecies cloning: the cloned Indian bison (using cattle oocytes) (Lanza et al., 2000), which was aborted at 202 days of gestation, and the cloned Bactrian camel (using dromedary oocytes) (Wani et al., 2017), which died seven days after birth.
Notably, one point worth highlighting is that, following the successful cloning of monkeys, the development of somatic cell cloning can be categorized into three distinct phases (as shown in Fig. 16):
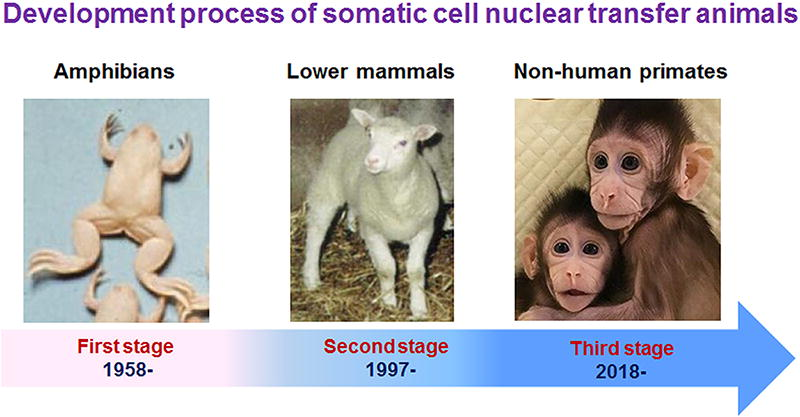
The development of somatic cell cloning in animals can be divided into three stages: (1) amphibian cloning research reported since 1958, (2) lower mammal cloning research reported since 1997, and (3) nonhuman primate cloning research reported since 2018.
The first phase: Cloning research on amphibians, beginning with reports in 1958;
The second phase: Cloning research on lower mammals, starting with reports in 1997;
The third phase: Cloning research on nonhuman primates, beginning in 2018.
If the cloning of monkeys were merely a replication of Dolly the sheep in a different species, one might ask: Was Dolly simply an extension of sheep embryo nuclear transfer using a different donor cell? Or was the cloning of frogs using somatic cells just a shift in species? The answers to these questions lie in the history of nuclear transfer: scientific innovation is not a sudden leap but a gradual progression built upon accumulated knowledge. It does not arise from unfounded imagination or unchecked flights of fancy, but from critical reflection and the inheritance of existing scientific understanding.
Significant Scientific Breakthroughs in Nuclear Transfer: Therapeutic Cloning, Prevention of Mitochondrial Diseases, and Androgenetic Haploid ESCs
The aforementioned research on cloning animals through nuclear transfer falls under reproductive cloning, which includes applications such as cloning livestock, preserving endangered species, producing genetically uniform animal models, and even cloning pets. Beyond these uses, nuclear transfer has also been instrumental in addressing several critical scientific challenges. These include the derivation of patient-specific ESCs, the prevention of mitochondrial disease transmission to future generations, and the enhancement of oocyte quality in older women. Additionally, nuclear transfer has been explored for generating sperm-like cells for in vitro culture and genetic modification, developing polyploid research models, investigating genomic stability, and uncovering mechanisms of cellular reprogramming. Furthermore, it has provided valuable insights into fundamental questions in epigenetics.
Therapeutic Cloning
Therapeutic cloning, in contrast to reproductive cloning, refers to the process where somatic cloned embryos are not implanted into a recipient but are instead used to isolate stem cells in vitro. These stem cells are referred to as SCNT ESCs. If stem cells can be obtained via nuclear transfer using patient-derived somatic cells, it would enable personalized medicine, theoretically eliminating the risk of immune rejection and holding immense clinical application potential.
Before the advent of induced pluripotent stem cells (iPSCs) (Takahashi and Yamanaka, 2006), nuclear transfer was the only method capable of reprogramming somatic cells into diploid pluripotent stem cells. The emergence of iPSCs provided an alternative approach, leading to a decline in therapeutic cloning research. This decline can be attributed to several factors. First, nuclear transfer requires highly specialized experimental equipment and advanced technical expertise. Second, the use of human oocytes is subject to strict regulatory and ethical constraints.
However, seven years after the introduction of iPSCs, human SCNT ESCs were finally successful (Tachibana et al., 2013), reigniting interest in therapeutic cloning. iPSCs offer numerous advantages, such as a more straightforward and widely applicable experimental process (currently, commercialized iPS Kits allow even ordinary laboratories to perform the procedure). However, SCNT ESCs avoid genetic modification, making their clinical applications potentially safer and more reliable. Therefore, SCNT ESCs and iPSCs can complement each other in research.
The following section summarizes the development of therapeutic cloning and the major breakthroughs achieved in this field.
First example of SCNT ESCs
In 2001, Teruhiko Wakayama and colleagues reported the isolation of ESCs from adult mouse somatic cells by SCNT, building on their previous work with cloned mice (Wakayama et al., 2001), as shown in Figure 17. This marked the first example of SCNT ESCs and provided an experimental foundation for human SCNT ESCs research.
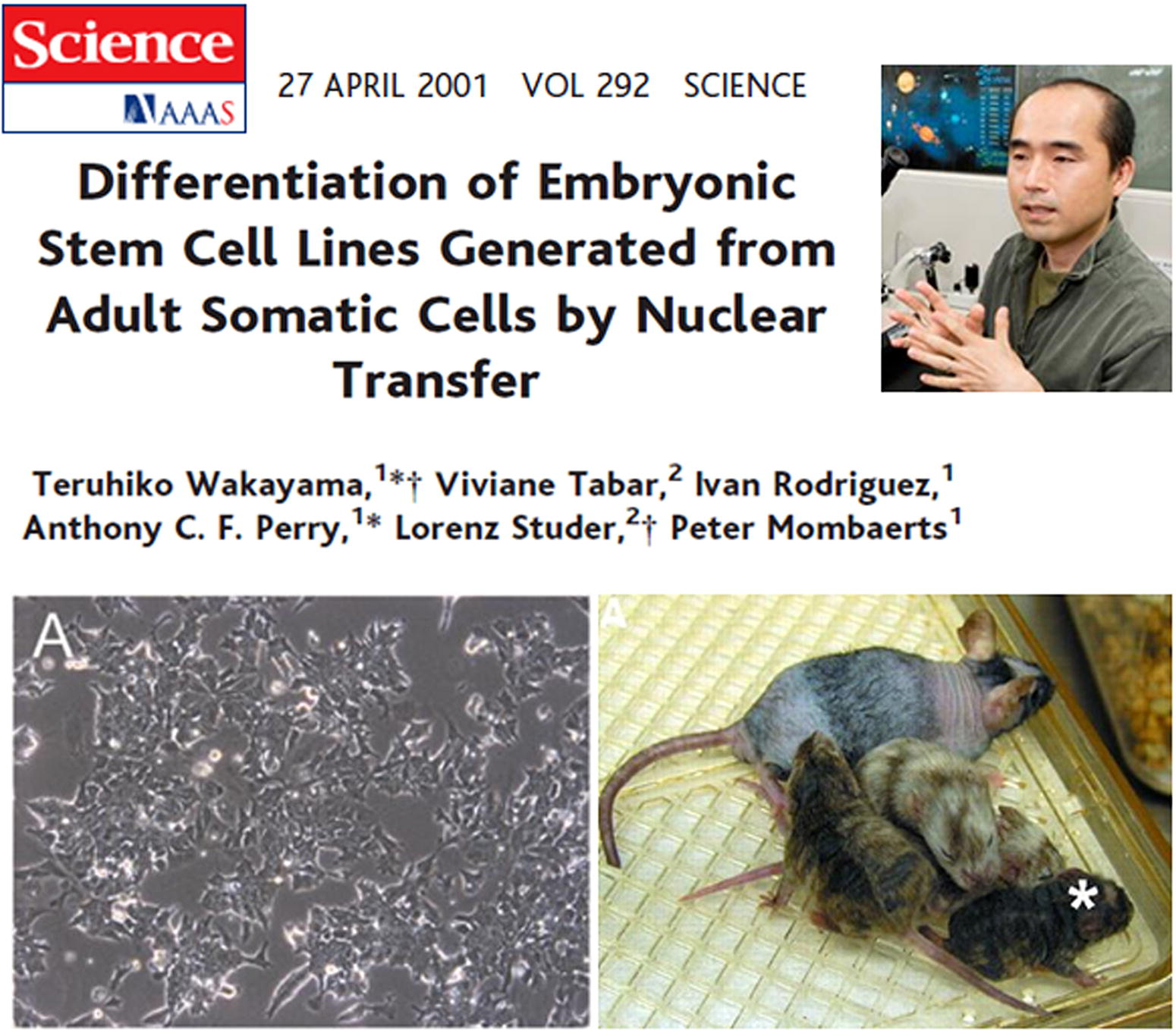
First example of somatic cell nuclear transfer ESCs. Teruhiko Wakayama et al. reported the first ESCs derived from cloned mouse embryos in Science in 2001. ESCs, embryonic stem cells.
First example of primate SCNT ESCs
In 2007, Shoukhrat Mitalipov and his team at the Oregon National Primate Research Center reported the successful isolation of two ESCs from cloned embryos created by SCNT of adult rhesus monkey skin fibroblasts. These ESCs not only exhibited a normal karyotype but also demonstrated pluripotency through in vitro and in vivo differentiation into all three germ layers (Byrne et al., 2007), as illustrated in Figure 18. This was the first example of primate SCNT ESCs.
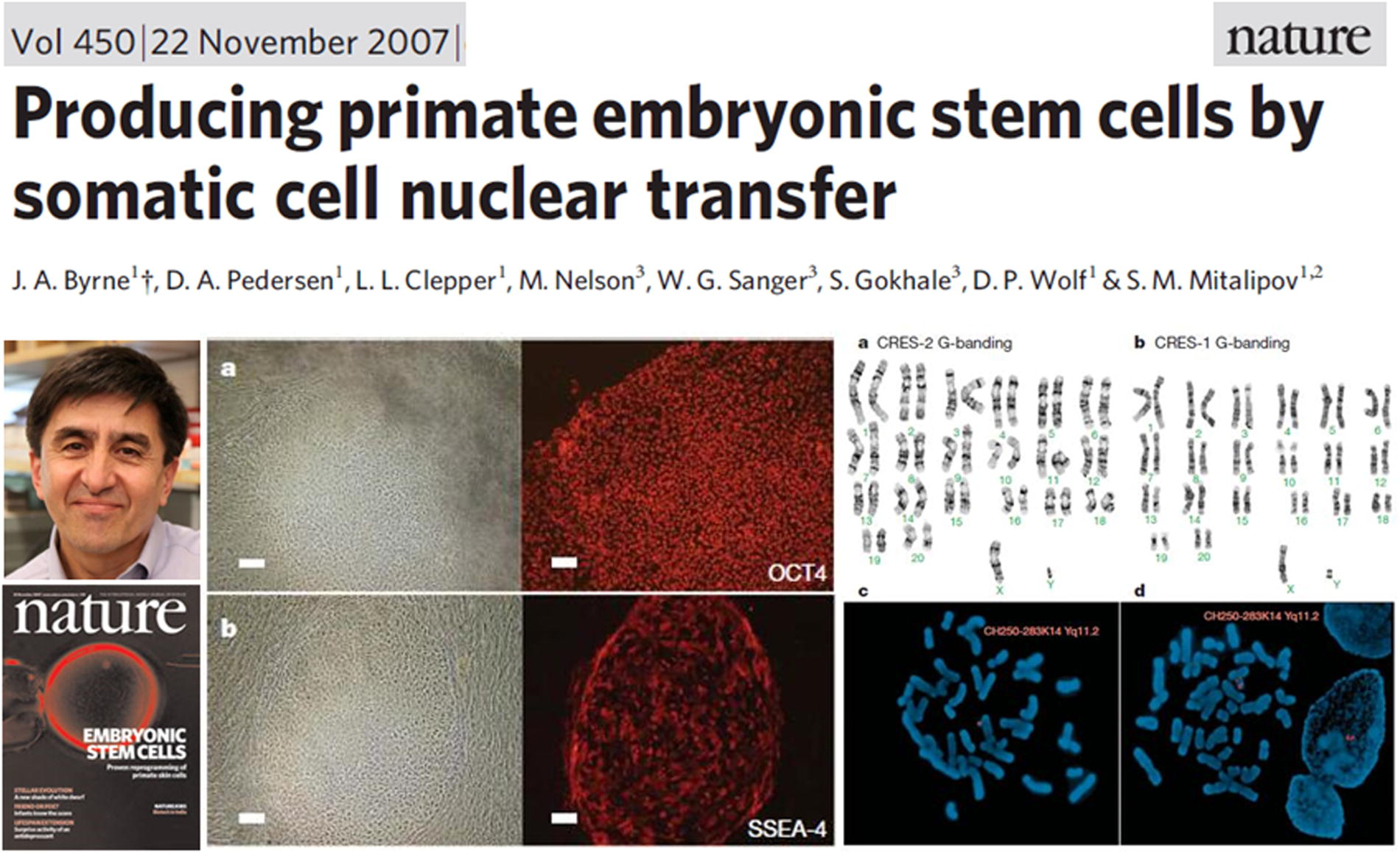
First example of primate somatic cell nuclear transfer ESCs. Shoukhrat Mitalipov et al.’s 2007 Nature cover article on nuclear transfer ESCs obtained from adult rhesus monkey skin cells.
First example of human SCNT ESCs
In 2013, Shoukhrat Mitalipov and colleagues built upon their work with rhesus monkey SCNT ESCs, successfully isolating four ESC lines with normal diploid karyotypes and pluripotent characteristics from cloned embryos generated via SCNT of human fetal skin fibroblasts (Tachibana et al., 2013), as shown in Figure 19. This marked the first successful derivation of human SCNT ESCs. Subsequently, additional laboratories reported the generation of ESCs from somatic cells of healthy adults (Chung et al., 2014), diabetic patients (Yamada et al., 2014), and individuals with age-related macular degeneration (Chung et al., 2015), reigniting interest in therapeutic cloning.
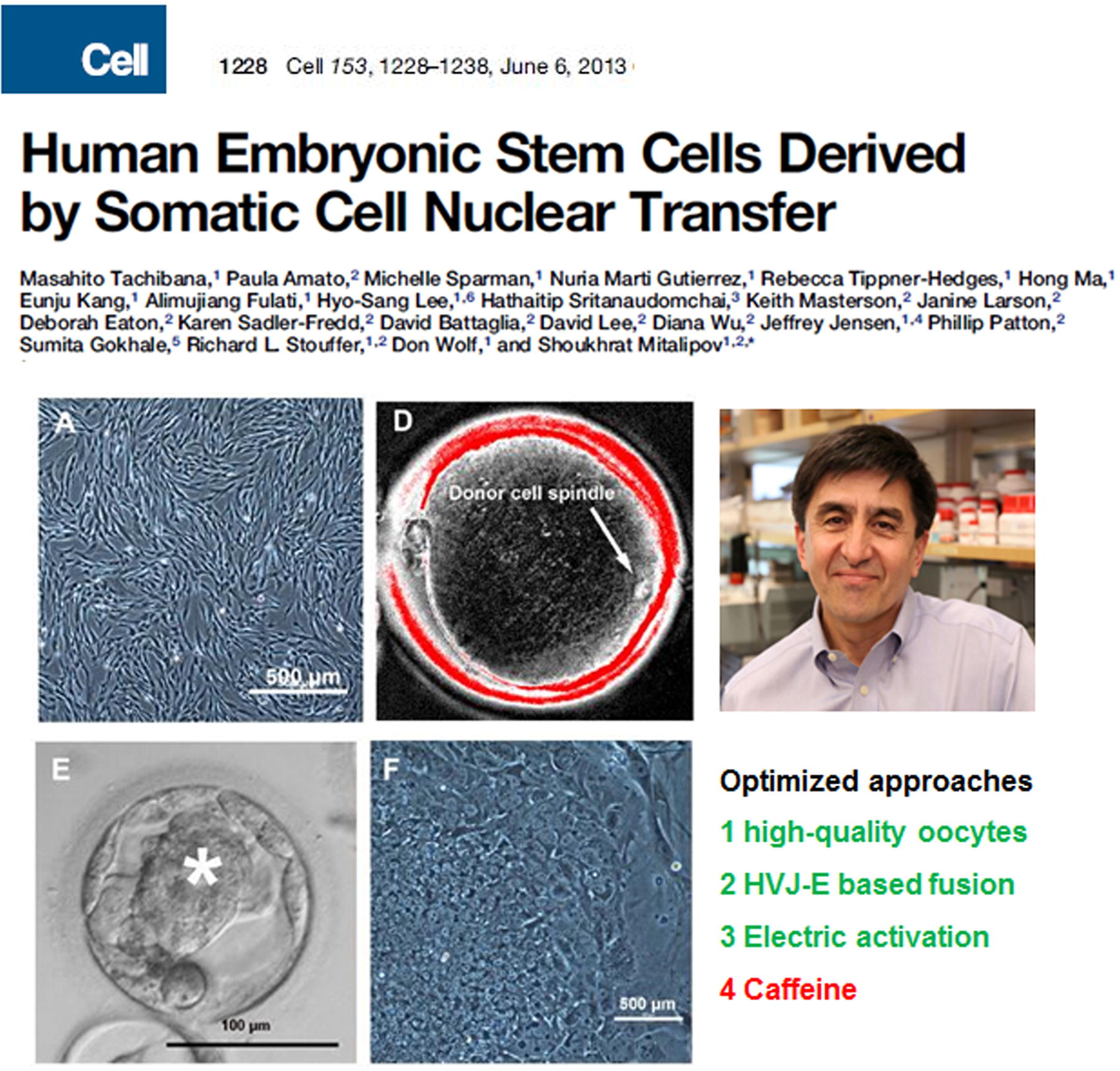
First example of human somatic cell nuclear transfer ESCs. Shoukhrat Mitalipov et al.’s 2013 study in Cell reporting nuclear transfer ESCs derived from human fetal skin cells. They improved their research by using high-quality oocytes and caffeine to enhance efficiency.
Prevention of Mitochondrial Diseases in the Next Generation
Mitochondrial diseases caused by mutations in mitochondrial DNA (mtDNA) are maternally inherited and passed down through the mother’s egg cells. Traditional assisted reproductive techniques cannot prevent the transmission of these mutations. However, nuclear transfer offers a potential solution. By replacing mutated mtDNA through approaches such as pronuclear transfer or transplanting the nucleus (or polar body) from the patient’s egg cell into a healthy, enucleated donor egg, it is possible to eliminate mutated mtDNA and prevent the disease in offspring.
Notably, despite its effectiveness, this method introduces the donor’s healthy mtDNA into the resulting child, meaning the individual inherits genetic material from one father and two mothers. This has raised ethical concerns regarding its application, particularly in relation to the implications of mitochondrial replacement on genetic identity and inheritance. As a result, its use remains a subject of debate, with regulations varying across different countries.
First mitochondrial replacement in a primate
In 2009, Shoukhrat Mitalipov and his team successfully performed the first mitochondrial replacement experiment in a primate. They extracted the spindle-chromosome complex from a rhesus monkey’s egg cell and transferred it into an enucleated egg cell from another monkey via the perivitelline space. The reconstructed egg was then fused using electrical pulses. After one to two hours, sperm was injected into the reconstructed egg’s cytoplasm to complete fertilization. The fertilized embryos were subsequently transferred into surrogate mothers, resulting in the birth of two healthy monkeys (Fig. 20) (Tachibana et al., 2009). This marked the first successful mitochondrial replacement in a primate, providing an experimental foundation for research into human mitochondrial diseases.
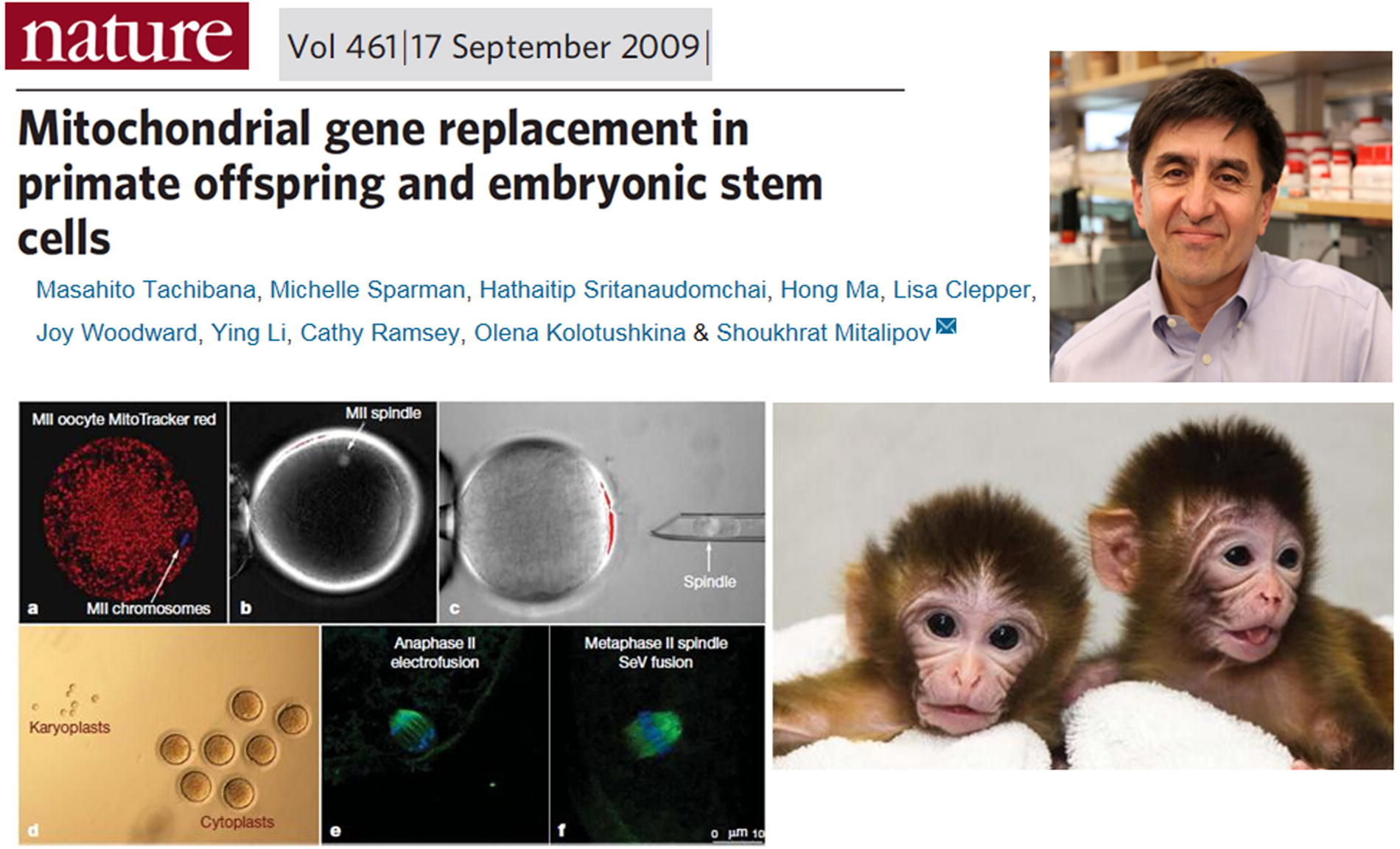
First mitochondrial replacement in a primate. Shoukhrat Mitalipov et al.’s 2009 report in Nature on the first mitochondrial replacement primate achieved through nuclear transfer in monkey oocytes.
First mitochondrial replacement—Human baby
In 2017, John Zhang and his team reported the successful application of mitochondrial replacement therapy, detailing the results of the first such procedure in humans. They used the same technique employed in the monkey experiment, transferring the spindle-chromosome complex from the egg cell of a patient carrying the mtDNA mutation responsible for Leigh syndrome into a healthy, enucleated donor egg. The newly created egg was fertilized with the husband’s sperm to form an embryo. After genetic screening of the embryo, it was implanted into the mother, resulting in the birth of a healthy baby boy (Fig. 21) (Zhang et al., 2017). This marked the world’s first baby born using egg nucleus transfer, colloquially referred to as a “three-parent baby.” However, the case raised significant ethical concerns, leading to warnings from the FDA directed at John Zhang.
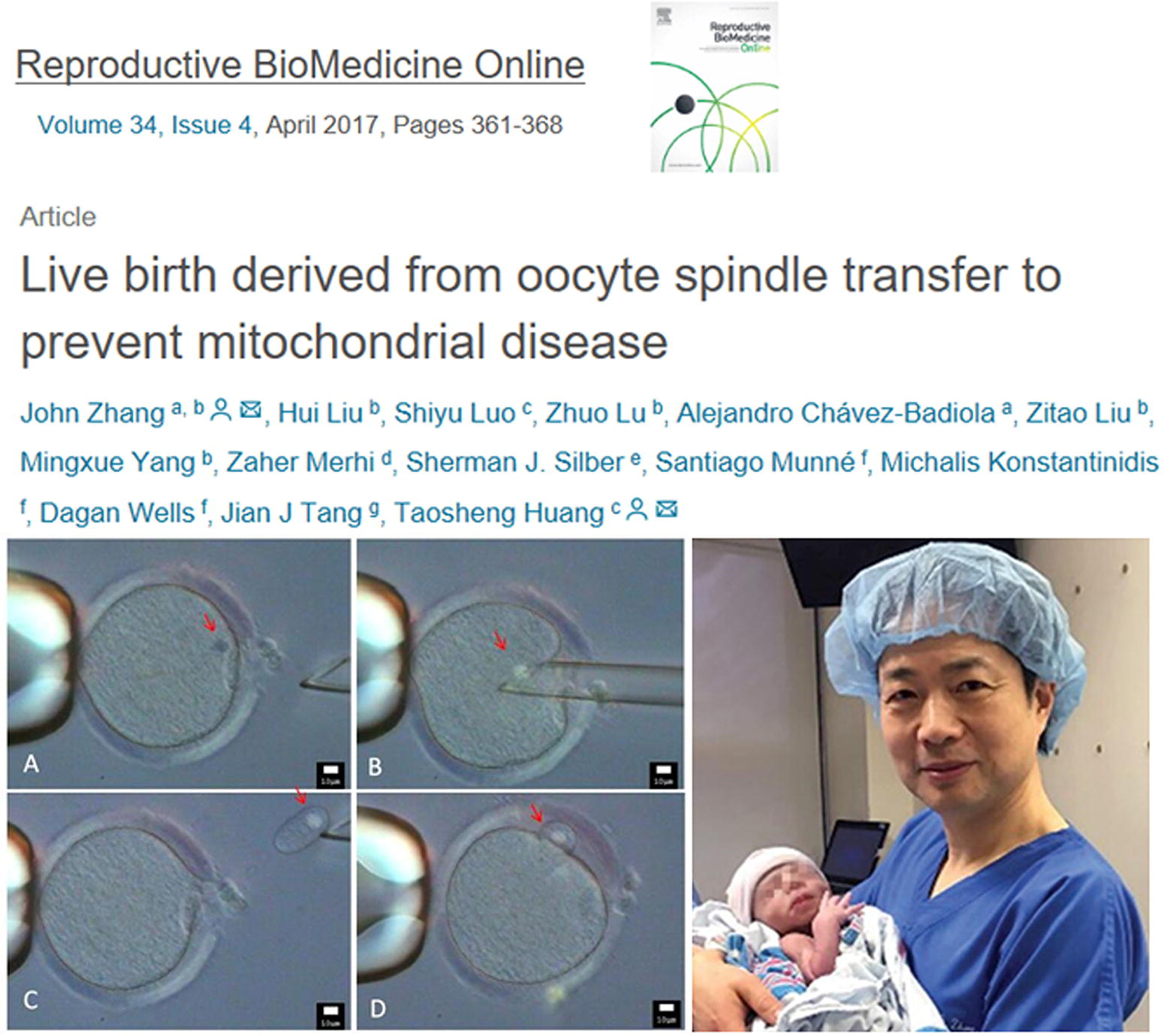
First mitochondrial replacement human baby. Zhang et al.’s, 2017 report on successfully producing a healthy baby by spindle transfer using oocytes from a Leigh syndrome carrier.
Androgenic Haploid ESCs—Sperm Cultured and Genetically Modified
In 2012, a groundbreaking study was reported by the research teams of Jinsong Li (the author’s postdoctoral mentor) and Guoliang Xu. They conducted an experiment in which sperm was injected into an enucleated mouse egg cell, followed by activation and cultivation. They observed that the reconstructed embryos developed in vitro to the blastocyst stage, from which ESCs were successfully isolated. Using fluorescence-activated cell sorting, they obtained haploid cells, which were named androgenetic haploid ESCs (AG-haESCs).
More significantly, these AG-haESCs were found to function similarly to sperm. When injected into a complete egg cell, the AG-haESC fertilized it and developed into a viable offspring. Additionally, when subjected to in vitro genetic modifications before fertilization, genetically modified mice were successfully produced (as shown in Fig. 22) (Yang et al., 2012). This study marked the first instance of transforming sperm into haploid stem cells capable of long-term in vitro cultivation, genetic manipulation, and maintaining reproductive potential. It represented a major scientific breakthrough, particularly in the context of China’s life sciences research.
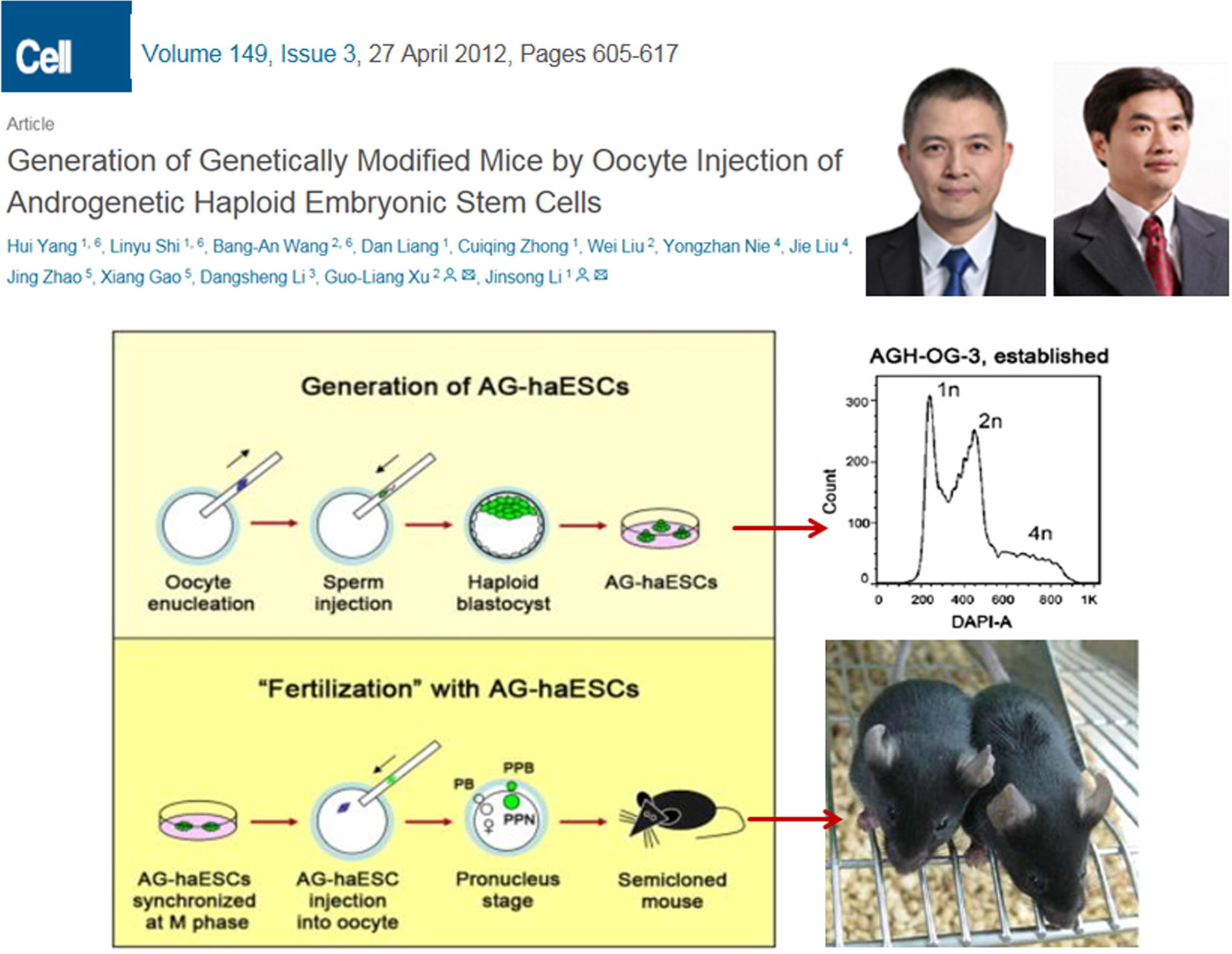
Androgenic haploid ESCs-sperm cultured and genetically modified. Li Jinsong and Xu Guoliang et al.’s 2012 study in cell on converting sperm into haploid ESCs that can be cultured and genetically manipulated in vitro using nuclear transfer methods.
Conclusion
From the creation of the first nuclear transfer animal to the first SCNT primate, nuclear transfer research has now reached over 96 years. Over nearly a century of exploration, this research has answered numerous fundamental scientific questions and has led to practical applications in various fields (Fig. 23). However, several challenges persist in nuclear transfer research:
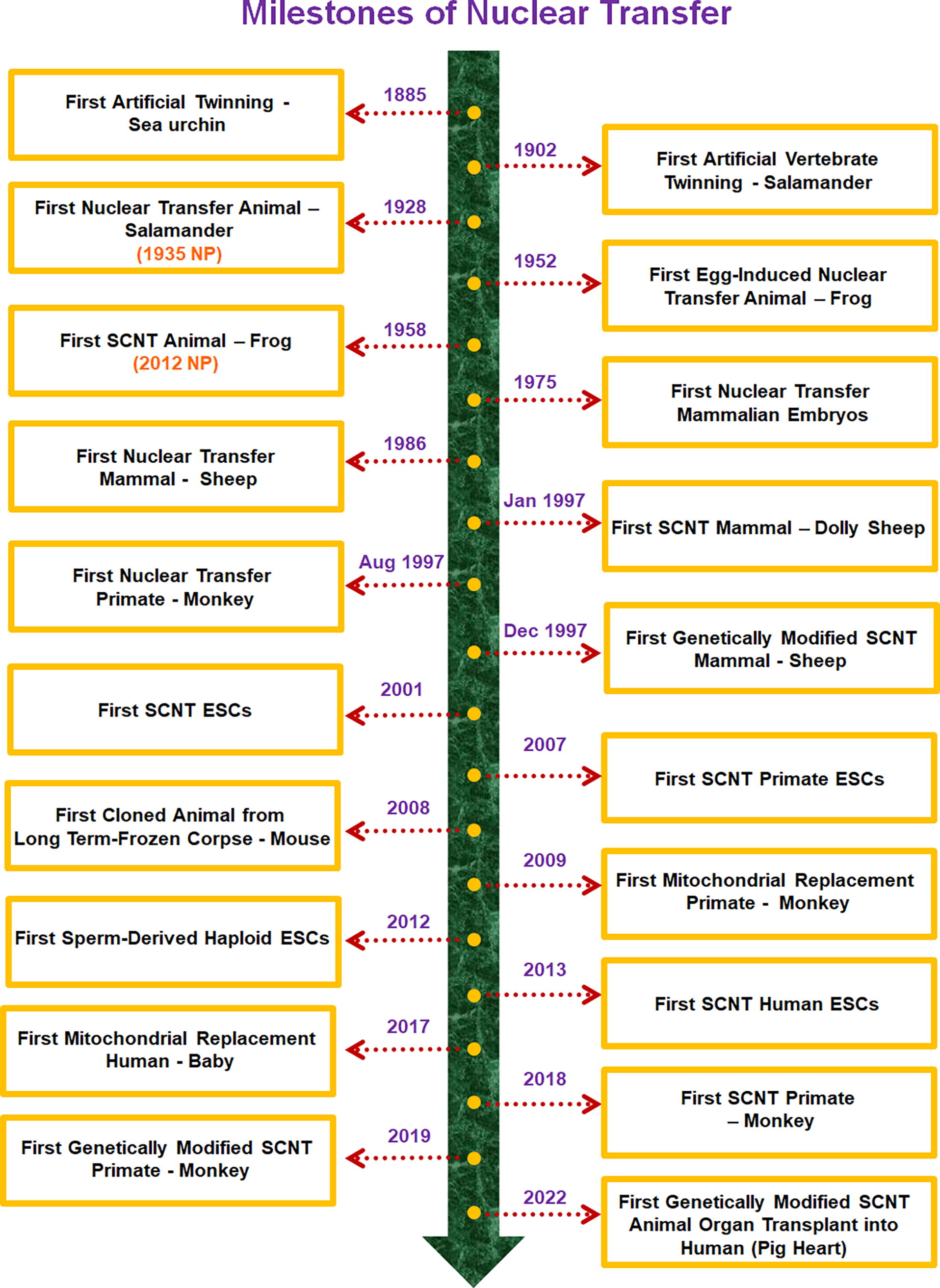
A timeline of major milestones in nuclear transfer (all dates refer to publication in peer-reviewed journals). The studies on the first nuclear transfer animal and the first somatic cell nuclear transfer animal were awarded the Nobel Prize (NP) in 1935 and 2012, respectively.
Low Efficiency of SCNT
The birth rate of cloned animals remains below 5%, significantly lower than clinical in vitro fertilization (IVF) success rates, which are typically ≥50%.
Interspecies cloning has proven exceedingly difficult, with healthy individuals rarely being obtained.
The efficiency of human NT-ESCs remains very low, with only a few laboratories achieving successful outcomes.
Limited Understanding of Epigenetic Reprogramming Mechanisms of SCNT
In the early 2000s, most studies on SCNT focused primarily on producing cloned animals, with limited exploration of the epigenetic barriers to reprogramming. With the development of sequencing technology, significant progress has since been made in understanding these mechanisms. In 2014, Yi Zhang and colleagues identified histone H3 lysine 9 trimethylation (H3K9me3) in donor cell genomes as a key epigenetic barrier to efficient reprogramming. Through comparative transcriptome analyses, they discovered reprogramming-resistant regions (RRRs) that are normally expressed in 2-cell mouse embryos generated by IVF but remain silenced in SCNT embryos. These RRRs were enriched with H3K9me3, and the ectopic expression of the demethylase Kdm4d reactivated most RRRs, dramatically improving SCNT efficiency (Matoba et al., 2014). Building on this foundational work, Liu et al. applied these findings in 2018 to achieve the first successful cloning of primates, demonstrating the translational potential of these epigenetic insights (Liu et al., 2018).
Despite these advances, the understanding of epigenetic reprogramming in SCNT remains incomplete. This limitation continues to hinder further improvements in efficiency and scalability, restricting the widespread use of SCNT-derived cloned primates as accessible animal models. Resolving these barriers is essential for broader applications in research and medicine. Without addressing these issues, SCNT-derived cloned monkeys may not become widely accessible as animal models, and cloning mammoths is merely a fantasy of humanity.
Ethical Concerns in Mitochondrial Replacement Therapy
Mitochondrial disease prevention through nuclear transfer faces significant ethical challenges, which continue to hinder its broader clinical application.
In conclusion, although many difficulties remain, as a researcher in nuclear transfer, I firmly believe that scientific progress will ultimately overcome these challenges, leading to further breakthroughs.
Author’s Contributions
Y.Q. conceptualized and wrote the entire manuscript and created all schematic illustrations.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.