
Abstract
The Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR/Cas9) system is revolutionizing genome engineering and is expected to bring significant advancements in livestock traits, including the treatment of genetic diseases. This study focuses on CRISPR/Cas9-mediated modifications of the CYP19 gene, which encodes aromatase, an enzyme crucial for converting testosterone to estrogen and essential for steroid metabolism. Guide RNAs (gRNAs) were designed to target the CYP19 gene and cloned into the pX459 vector. The recombinant plasmid was then electrotransfected into fibroblast cells from a Lori-Bakhtiari buck, and these transfected cells were used for embryo production via somatic cell nuclear transfer (SCNT). The cloned embryos were evaluated for their progression through embryonic stages, showing no significant difference in blastocyst development between knock-out and unedited groups. The knockout efficiency was 78.4% in cells and 68.9% in goat blastocysts, demonstrating the successful depletion of CYP19. We successfully achieved a high rate of CYP19 gene-edited embryos through the combined application of cell electrotransfection and SCNT technologies, while maintaining the normal developmental rate of the embryos. These embryos can be used for transfer to generate knock-out goats, providing a foundation for further studies on CYP19’s role in male fertility and production traits.
Introduction
Today, it is possible to modify certain traits in animals through genome editing without introducing foreign genes. The latest genome editing method, Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR/Cas9), enables targeted gene modifications. The CRISPR/Cas9 system allows for precise editing at the desired sequence by removing or adding specific DNA fragments (Doudna and Charpentier, 2014; Gasiunas et al., 2012). This method has proven to be an efficient tool for genome modification in genetic model animals such as mice (Eghbalsaied et al., 2020; Horii et al., 2014), monkeys (Kang et al., 2015), pigs (Wang et al., 2015a), and some livestock such as sheep (Crispo et al., 2015) and goats (Wang et al., 2015b).
The primary androgens are testosterone and androstenedione, which are present at much higher levels in males and play significant roles in male traits and reproductive activities. Estrogens are steroid hormones largely produced in the ovaries and testes (Katsu et al., 2010). The cytochrome P450 aromatase enzyme is a microsomal member of the cytochrome P450 superfamily (CYP) and the only member of family 19 (CYP19A1) (Simpson and Davis, 2001). It is assumed that most vertebrates possess a CYP19 gene regulated by several tissue-specific promoter regions (Callard et al., 2001; Simpson et al., 1997). CYP19 encodes aromatase, a crucial enzyme responsible for steroid metabolism, playing a fundamental role in converting testosterone to estrogen (Roselli and Resko, 2001). If CYP19 is knocked out or its activity diminished, estrogen production from testosterone also decreases (Kamat et al., 2005; Mohammadrezaei et al., 2014; Rahaie et al., 2018). This can lead to hormonal imbalances and may affect physical gender development (Wang et al., 2001). It has been verified that CYP19 manipulation leads to sexual inversion in avian and fish species (Jeyasuria and Place, 1998; Kobayashi et al., 2003; Mohammadrezaei et al., 2014; Zhang et al., 2023). Although overexpression of CYP19 in goat was associated with higher litter size (Zhang et al., 2022), disruption of this gene in goat bucks remained to be assessed.
In this study, we selected the Lori-Bakhtiari goat breed, which is not only economically valuable but also an integral part of the agricultural heritage and sustainability of pastoralist communities in Iran (Javdan, 2014). To date, no mutations have been reported in the CYP19 gene of Lori-Bakhtiari goats in Iran. To edit the CYP19 gene, we combined CRISPR/Cas9 technology with somatic cell nuclear transfer (SCNT). Fibroblast cells were derived from a male fetus, genetically modified using CRISPR/Cas9, and subsequently used for embryo production. The use of SCNT helps overcome the issue of mosaicism associated with the microinjection technique (Kalds et al., 2019). Our primary objective was to knock out the CYP19 gene, located on chromosome 10 in goats, and assess its effects on embryonic development.
Materials and Methods
Cloning of guide RNAs into pX459 vector
The guide RNA (gRNAs) (20 base pair sequences) was designed and scored using the CHOPCHOP software. Initially, the target gene sequence was selected from NCBI. Subsequently, two gRNAs and primers were designed for exon 10 of CYP19 (Table 1). Cloning of gRNAs into the Cas9-carrying vector, the pX459 plasmid (9151 base pairs) was carried out based on previously published protocols (Eghbalsaied and Kues, 2023; Guo et al., 2018). Briefly, the plasmid encodes the Cas9 protein along with a puromycin resistance gene and a scaffold gRNA (under the U6 promoter). The pX459 plasmid was digested with BbsI-HF (NEB #R3539) and the purified fragment was used as the backbone for the ligation of the oligonucleotide duplex containing the gRNA sequence. The recombinant vector was transformed into competent bacteria (NEB 5-alpha Competent Escherichia coli, #C2987I) followed by colony selection on agar plates containing ampicillin antibiotic. The cloned plasmids were digested with restriction enzymes BbsI and EcoRV, potentially correct ligated plasmids produce a linearized vector in this informative digest, while the original plasmid will be cut into two fragments. Sanger sequencing confirmed the correct cloning.
The Sequence of Two gRNAs (CYP19-41 and CYP19-64) and Primers for CYP19 Gene Amplification
Cell culture and electrotransfection
The fetal fibroblast cells were derived from a 40-day-old male embryo sired by a distinguished buck for large size from the Iranian Lori-Bakhtiari breed, Iran. Fibroblasts were cultured in DMEM culture medium supplemented with 10% fetal bovine serum (FBS), 2 mM glutamine, 1% sodium pyruvate, and 1% penicillin-streptomycin (Pen Strep) under 5% CO2 at 37°C until approximately 80%–90% confluence was achieved in the dish.
We used the previously optimized electroporation technique for the transfection of plasmids carrying the CRISPR system into the cells (Eghbalsaied et al., 2020; Eghbalsaied and Kues, 2023). Briefly, cultured cells were washed with PBS and dissociated using trypsin. The cells were then washed and mixed with 20 µg of plasmid in a transfection medium containing 250 µL of OptiMEM-GlutaMAX and subjected to electrotransfection using a Bio-Rad device, two pulses of 270 v, 10 ms, 10-second interval in a 4 mm cell holder. Puromycin selection started 6 hours post-electrotransfection and continued for 2 days (Eghbalsaied et al., 2020; Eghbalsaied and Kues, 2023).
Somatic cell nuclear transfer
Ovaries were collected from slaughtered goats at an industrial abattoir in Isfahan and transported to the laboratory in saline solution at 17°C. Follicles measuring 2–4 mm in diameter were aspirated using a vacuum pump, and the aspirated material was processed at 38°C. The aspiration medium was the HEPES-buffered tissue culture medium 199 (HTCM) supplemented with 15% FBS and 10 μl/mL heparin. Under a stereo microscope, cumulus-oocyte complexes (COCs) with granular cytoplasm and compact cumulus layers were selected. These COCs were then washed in HTCM medium supplemented with 15% FBS. For in vitro maturation (IVM), COCs were cultured in TCM199 medium enriched with 15% FBS, 10 µg/mL luteinizing hormone, 10 µg/mL follicle-stimulating hormone, 1 µg/mL Estradiol, 100 ng/mL IGF (insulin-like growth factor), and 10 ng/mL EGF (epidermal growth factor). Groups of five COCs were placed in a microdrop of 50 µL IVM medium and incubated at 38.5°C with 5% CO2 and maximum humidity for 20–22 hours. Matured oocytes displaying uniform cytoplasm were treated with hyaluronidase to remove cumulus cells in preparation for nuclear transfer. Enucleation of matured oocytes adhered to Royan Research Institute protocols (Sadeghi et al., 2023), The zona pellucida was removed using pronase treatment, and oocytes were incubated in TCM medium containing 400 ng/mL demecolcine, 3 mg/mL polyvinyl alcohol (PVA), and 10 mg/mL BSA for 20 minutes. Enucleation was performed manually by aspirating the protruded pronucleus, induced by demecolcine incubation, using a thin needle connected to a mouth pipet. Successfully enucleated oocytes were incubated in a maturation medium for 30 minutes before the cell fusion step.
To prepare the donor cells, fibroblast donor cells were synchronized at the G0 phase of cell division by culturing them in media supplemented with 0.5% FBS for 3–5 days. On the day of oocyte-cell fusion, the donor cells were trypsinized and resuspended in HTCM containing 0.5% FBS. For the fusion process, enucleated oocytes and fibroblast cells were placed in a medium containing 0.15 mg/mL BSA, 1 mg/mL PVA, 5 ng/mL Lectin, 10.98 g Mannitol, and 0.004 g MgCl2·6H2O, with an osmolality of 280–300 mOsmol. The reconstructed oocytes were then washed in a fusion medium (0.3 M Mannitol, 0.1 mM MgSO4) and positioned in a fusion chamber between electrodes spaced 0.5 mm apart. Fusion was induced by applying an electric pulse (1.75 kV/cm, 80 µs), while an AC pulse (800 kHz) facilitated cell alignment. The success of fusion was assessed 1 hour after the pulse. Activated oocytes were first exposed to ionomycin (5 µM) for 1 minute, followed by a 5-minute inactivation in TCM medium supplemented with 30 mg/mL BSA. They were then incubated for 2 hours in 6-dimethylaminopyridine (DMAP)-supplemented medium, consisting of synthetic oviductal fluid, 10% FBS, 0.2 mg/mL PVA, and 10 µL 6-DMAP in a total volume of 500 µL. Cloned embryos derived from fused fibroblast cells were individually cultured in a handmade well of well (WOW) system within 4-well plates. Cleavage was assessed on day 1, while blastocyst formation was evaluated on day 7. Two sets of 10 blastocysts were randomly selected from each group, pooled together, and used for PCR and gene amplification (Eghbalsaied et al., 2013).
Statistical analysis
SCNT embryo production was conducted in vitro across eight independent runs. An independent t-test was used to compare the mean cleavage and blastocyst rates between CYP19-edited and non-edited groups, with a significance level of α = 0.05.
Results
CYP19 gene knock-out in cells
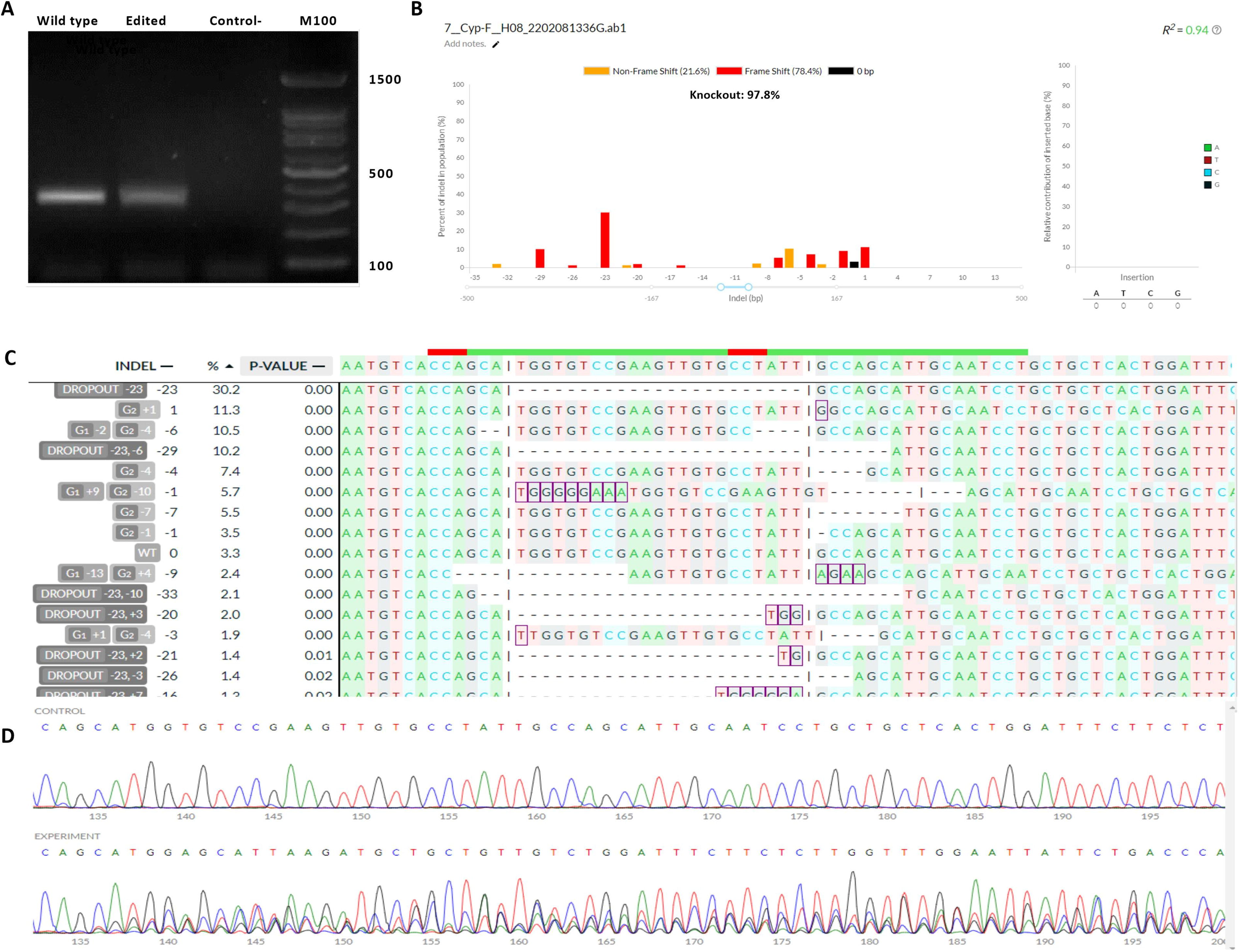
Cells were assessed for knock-out efficiency through PCR testing and sequencing. The agarose gel analysis related to genotype determination based on PCR is illustrated in Figure 1. Positive control cells exhibited the expected product on the gel, confirming successful amplification. Edited cells for the CYP19 gene were sequenced alongside controls to assess editing (Fig. 1). The sequencing results from both the control and gene-edited groups were analyzed using deconvolution of complex DNA repair (DECODR) software. This analysis confirmed a 97.8% gene editing efficiency in the bulk cells, with 78.4% of the edits associated with a frameshift mutation or gene knockout rate and 49.6% associated with an on-target fragmental deletion between two gRNA targeting sites (Fig. 1).
Development of CYP19 gene knockout embryos
The developmental progression of cloned embryos derived from fibroblast cells was assessed, focusing on their advancement to the cleavage and blastocyst stages. In eight independent SCNT runs using CYP19-edited cells, 410 matured oocytes were used, yielding 376 cleaved embryos—equivalent to a 91.7% developmental rate for embryos with CYP19 gene knockout. However, this cleavage rate did not differ significantly from the non-edited control group, which consisted of 324 matured oocytes and 298 cleaved embryos (p = 0.43).
To visualize these findings, we plotted graphs illustrating the number of cleaved embryos relative to total oocytes, the ratio of blastocysts to cleaved embryos, and the ratio of blastocysts to total oocytes used (Fig. 2). The blastocyst development rate was 44.1% for CYP19-edited cells and 39.5% for non-edited cells. However, no significant differences in blastocyst percentages were observed between any groups compared with the control (p = 0.29) (Fig. 2).

Developmental progression of SCNT-derived embryos. Shown are the cleavage rate, blastocyst rate relative to the number of embryos, and blastocyst rate relative to the total number of oocytes. The control group is represented in red, and the CYP19-edited group is also shown in red. SCNT, somatic cell nuclear transfer.
CYP19 gene knockout in blastocysts
The blastocysts were examined prior to Sanger sequencing using PCR. The agarose gel related to genotype determination based on PCR is illustrated in the accompanying Figure 3A. Our gene-edited groups using double gRNAs showed two bands, a larger band corresponds to the band in the non-edited group and a shorter band corresponds to a fragmental deletion between two gRNAs. Each 10 blastocysts were pooled and directly used for PCR. The edited blastocysts were sequenced alongside controls to ensure the accuracy of the gene editing. The Sanger sequencing results indicated that the editing performed using the CRISPR/Cas9 system resulted in a 68.9% (±16.4%) knockout rate (due to frameshift mutations) in the CYP19 gene (Fig. 3B–D). As shown in Table 2, output data from DECODR software revealed that in the first trial, 70.5% of cells were associated with gene editing, with 52.5% resulting from a frameshift mutation and 18% from non-frameshift mutations. In the second trial, a 90.9% mutation was observed, with 85.3% due to frameshift mutations and 5.6% due to non-frameshift mutations. In addition, using two gRNAs caused 46.2% of on-target fragmental deletion between two gRNAs.

The Rate of Mutations Created in CYP19 Gene Embryos
CYP19-1 and CYP19-2 are two pooled repeats of embryos.
SD, standard deviation.
Discussion
Genome editing technology, especially CRISPR/Cas9, enables precise modifications of genomes through targeted insertions or deletions (indels). In this study, we used CRISPR/Cas9 and somatic cell nuclear transfer (SCNT) to produce CYP19-edited goat embryos. This gene plays a role in sexual differentiation and reproductive traits. We achieved a knockout efficiency of 78.4% in cells and 68.9% in embryos, confirming the feasibility of editing the CYP19 gene in goat fibroblast cells and embryos. Although electrotransfection was highly efficient, cell electroporation and puromycin selection were not entirely effective. Consequently, a portion of the bulk population likely includes non-transfected cells. However, the high mutation rate in bulk cells (>70%), encompassing both transfected and non-transfected cells, suggests that most mutations were biallelic.
CRISPR-based systems have been initially employed in small ruminants to investigate biological functions of economically valuable genes, such as MSTN (myostatin), which enhances meat production and DAZL, involves in sex differentiation (Pirali et al., 2024; Wang et al., 2015b). Most researchers have injected mRNA encoding Cas9 and single-guide RNA (sgRNA) into single-cell stage embryos (Ni et al., 2014). However, somatic cells, as utilized in this study, can serve as donors for SCNT to produce targeted gene-modified animals. This can greatly help with pre-screening the donor cells for desired mutations before making embryos (Kwon et al., 2024). Moreover, the use of non-edited donor cells from the same cell line can provide a specific control group with the same nuclear genetic content except for the edited gene (Oback, 2009). Notably, there are currently no reports on producing edited goat cells and embryos for the CYP19 gene using CRISPR and SCNT techniques aimed at improving any traits, including reproductive characteristics.
Utilizing powerful genome editing tools such as TALEN and CRISPR/Cas9, the gonadal-specific aromatase gene CYP19a1a was disrupted in zebrafish (Callard et al., 2001). They found that all three mutant lines produced were male, confirming that gonadal aromatase and the estrogens it produces are vital for guiding gonadal differentiation. Notably, no abnormalities in spermatogenesis were observed in the mutant fish, and their fertility remained normal (Callard et al., 2001). Interestingly, a study investigated spermatogenesis in mice lacking aromatase due to targeted disruption of the CYP19 gene (Robertson et al., 1999). These male mice were initially fertile but exhibited progressive infertility until their ability to sire offspring was severely compromised (Robertson et al., 1999). Their findings suggest that local expression of aromatase is essential for spermatogenesis and provide evidence for the direct effect of estrogen on the growth of male germ cells and consequently fertility (Robertson et al., 1999). While previous research has targeted CYP19 in species like zebrafish and mice, no studies have examined its knockout in goat embryos using CRISPR/Cas9, making this research novel.
In conclusion, the results of this study showed that compared with the non-edited group, disrupting the CYP19 gene does not affect the embryonic development rate until the blastocyst stage is reached. The present achievement may also open up new possibilities for efficient CRISPR/Cas9-mediated generation of not only goats but also other ruminant and mammalian species displaying genetically engineered reproductive traits (e.g., fertility, fecundity/prolificacy) with the aid of either somatic cell cloning (Davies et al., 2022; Hao et al., 2018; Samiec et al., 2012; Samiec et al., 2022) or by editing genomes/multiple genes in the in vitro-fertilized oocytes and zygotes (Fernández et al., 2024; Hashimoto et al., 2016; Pi et al., 2024; Piñeiro-Silva and Gadea, 2024; Samiec, 2024). However, the long-term effects on goat fertility remain unknown and warrant further investigation. This study may enhance livestock production by modifying reproductive characteristics and other traits, though it requires careful consideration of ethical, health, and economic impacts.
Footnotes
Acknowledgment
The authors would like to appreciate all the staff of Royan Institute for Biotechnology.
Authors’ Contributions
Conceived the idea: S.E. and M.N.; designed the experiment: S.E., W.K., M.N., F.J., and N.H.; carried out the experiments: A.P., F.J., M.H., S.H.H., R.M., N.T., M.R., T.I., H.S., Z.S., and S.E.; analyzed the data: S.E.; wrote the article: A.P., F.J., W.K., M.N., and S.E.; all authors read and confirmed the article content.
Confirmation Statement
Each author confirms that their research is supported by an institution that is primarily involved in education or research.
Disclaimer
At the time of the project, S.E. was affiliated with the Royan Institute for Biotechnology, the Isfahan (Khorasgan) Branch of Islamic Azad University, and the University of Melbourne.
Ethics Declaration
The Research Committee of Royan Institute, Tehran, approved this project on November 26, (Certificate Number: IR.ACER.ROYAN.REC.1401.045). The committee was chaired by Dr. Abdolhossein Shahverdi, with Dr. Reza Omani Samani serving as the Committee Secretary.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This project was partly funded by the grant number: Ref 3.4—IRN—1191261—GF-E (Georg-Forster-Fellowship to S.E.) from the Alexander von Humboldt Foundation, Germany, and also partly funded by the following grant: ID: CT1402091612 from Biotechnology Development Headquarters, Technology and Knowledge-Based Economy, Iranian Vice President for Science.