
Abstract
Stem cells are key to human tissue maintenance. Because tissue maintenance allows us to live and reproduce, stem cell control is fundamental for animal life and evolution. A team of researchers set out to explore the origins of transcription factors at the core of the induction and the maintenance of stemnss. They focus on the conservation of the Sry-related box 2 (Sox2) and the octamer-binding transcriptor factor 4 (Oct4) in the Pit-Oct-Unc (POU) family. While these have been thought as animal-specific, the authors identified SOX and POU in pre-animal organisms. In particular, the SOX protein from a very simple unicellular organism was functionally conserved enough to reprogram somatic mouse cells to induce pluripotent stem cells. To ponder on the importance of their findings, we first need to step back a couple of hundred million years.
First, the definition of animals is not entirely straightforward. The simple story states that animals are fairly similar to each other (often assumed similar to us, with a backbone), in constant need of oxygen, and with specific stem cells in niches to maintain tissue renewal. The more complex story is that 97% of all animal species lack a backbone (invertebrates), cope well in low-oxygen conditions, and have a much different tissue regeneration, if any. Mature nematodes live and die with the same set of somatic cells (Sulston et al., 1983). Insects can “patch” a wound by having one epithelial cell grow larger (Zhong et al., 2023). Sponges are also considered animals despite lacking true tissues. Indeed, adult sponges reconfigure the architecture and fate of cells continuously, likely several times a day (Fernàndez-Busquets et al., 2002; Sogabe et al., 2019). This means that the role and definition of stem cells varies significantly even within animals. Across the evolution of different versions of cell fate control, we do not yet fully understand the common denominators or how it all began.
Second, the rise of animals can also be told as a simple or more complex story (Mills and Canfield, 2014). The simple story being that the “rise” of animals began some 700 million years ago as a result of oxygen concentrations increasing in the atmosphere. The more complex story is that early animals of low diversity were around long before the clade diversified, and that this diversification was triggered by subtle biological factors (Marshall, 2006). One such biological factor could be the advent of predation and another, newer suggestion, is that variations to stemness control, and selection between these, led to higher fitness and animal diversification (Hammarlund et al., 2018). This suggestion means that we should compare stemness control in animals and in our unicellular ancestors.
Do we know if biological factors, rather than increasing oxygen, led to the rise of animals? The answer is no, even if geobiologists do their best to puzzle together the history of Earth and life. Do we know how molecular innovations could lead to spatiotemporal control of immature or differentiated cell fates? Not yet. However, the recent work by Gao et al. has indicated that SOX and POU factors were not animal-specific innovations (Gao et al., 2024). If key stemness factors appear in unicellular organisms, it supports the model that additional forces selected for stemness-related adaptations, which led to increased population fitness. Then, early cellular mechanisms that were useful for function X could have been co-opted for additional and, at the time, useful functions X and Y.
Gao et al. first found that a Sox-like HMG box sequence is present in two different unicellular “ancestors” to multicellular animals. These unicellular eukaryotic ancestors are called filastereans and choanoflagellates, where the latter are composed of filter-feeder cells similar to the differentiated cells in the sponge animal. Then, they showed that choanoflagellate SOX protein was able to replace SOX2 when inducing pluripotency in somatic mouse cells, cooperating with mammalian OCT4 (which is specific to vertebrates) to interact with DNA. While the choanoflagellate POU had a distinct DNA binding profile and could not induce pluripotency, its gene is present in this unicellular ancestor of ours (Fig. 1).
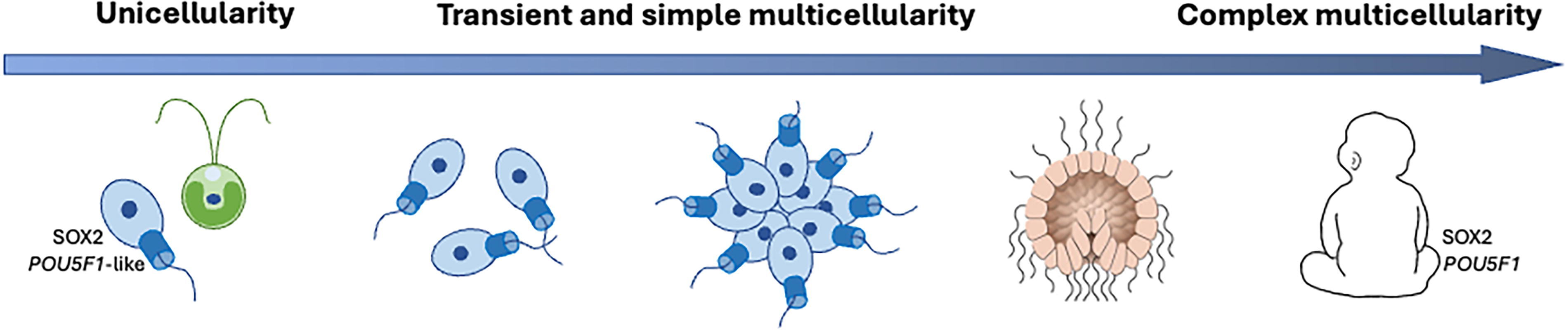
A schematic representation of the evolutionary transitions from eukaryotic unicellularity (protists such as choanoflagellates) to transient and simple multicellularity (e.g., aggregations of choanoflagellates) to animals, defined by e.g., gastrulation, from sponges to humans. Across this evolution over more than 700 million years, SOX2 (protein and gene) and POU-like genes are functionally conserved.
Thus, this work by Gao and colleagues contradicts a strict animal-specificity of key stemness markers and even indicates that the SOX family and SOX protein family have remained functionally conserved for hundreds of iterations and millions of years. The team hypothesizes that these systems were available before and thereafter co-opted for different versions of stemness control. Their results and thoughts are consistent with previous work exploring our unicellular ancestors, where comparative genomics and multiomics suggest they had complex life cycles that contained several cell fates (Sebé-Pedrós et al., 2017). This would mean that rather than controlling several cells with different fates in space, unicellular ancestors could control them primarily in time.
Next, one can follow the roots of SOX into even earlier organisms. However, it is also relevant to ask: what use do unicellular organisms have for SOX and POU? What conditions made it a favorable mechanism to co-opt for advanced stemness control? As multicellular organisms were tried out in varying environments back in time, perhaps its role for handling cell fate or stress proved useful. Although stressors likely differ for single-celled organisms in the ocean and a cell within an early animal tissue, coping well with stressors is likely to confer higher fitness. On that note, cancer cells that are observed to be fit, for example, use the stress-response Sox4 pathway (Jiang et al., 2024).
Regardless of how deep the SOX roots go, Gao et al.’s work opens new pathways to understand the origins of human stemness control which, in the long run, could emphasize the requirements and vulnerabilities of stem cell niches.
Footnotes
Acknowledgment
The author thanks Dr. Mazharul Islam and one anonymous reviewer for comments and advice.
Author’s Contributions
E.U.H. conceptualized, wrote the text, and made the figure.
Author Disclosure Statement
The author declares no conflicts of interest.
Funding Information
Insights are achieved in a project funded from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (grant agreement No 949538).