
Abstract
Abstract
Background:
Two common missense variants in the melanocortin-3 receptor (MC3R) gene, Thr6Lys (T6K) and Val81Ile (V81I), are presumably correlated with pediatric obesity. This meta-analysis aimed to examine and synthesize evidence on the association between these two common MC3R polymorphisms and the development of childhood obesity.
Methods:
A combination of words relevant to the research question was searched on PubMed, EMBASE, Scopus, and the Cochrane database. Results were restricted to human studies, specifically child and adolescent populations. Articles were excluded based on accessibility of full online texts and availability of pertinent data. Pooled odds ratios (ORs) and 95% confidence intervals (CIs) were calculated using a random effects model to determine the association of the polymorphisms with obesity.
Results:
Searches on the databases using the keywords identified 65 potentially relevant reports. Among them, 32 studies were excluded due to irrelevance, and 28 studies excluded due to lack of access, insufficient data, and investigation of other variants. A final set of five studies included in this meta-analysis found that the risk of overweight/obesity increased by 46.1% per K allele and 21.7% per I allele. Only homozygous genotypes for T6K were associated with a 3.10-fold (95% CI: 1.29–7.43) increased risk of overweight/obesity in children. Data were insufficient to examine if homozygosity for both rare alleles further increases risk.
Conclusions:
Our results supported a recessive inheritance model for MC3R gene as a potential cause of childhood obesity. High clinical heterogeneity existed among studies and thus requires more research of larger participation for future integration of data.
Introduction
Obesity is currently a global health concern, with over 700 million people being overweight or obese worldwide, and it continues to rise in prevalence in certain developing countries.1,2 The prevalence of obesity is projected to increase by 33% over the next two decades. 3 Although the forecast is principally based on data from adults, time trends in childhood obesity prevalence are believed to impact adult obesity prevalence and related healthcare costs, given a strong correlation consistently found between childhood and adulthood weight status and obesity-related behaviors.4,5 In adults, obesity is a major risk factor for certain chronic conditions, including diabetes and cardiovascular diseases,6,7 the latter of which is globally a leading cause of death. 8 Obese children also experience respiratory problems, insulin resistance, hypertension, and psychological effects. 9
Obesity is typically viewed as a multifactorial condition, with both environmental and genetic factors playing a role in its development. Rare cases of monogenetic obesity due to single gene mutations have been better described with an early onset in childhood and in association with endocrine or behavioral disturbances. 10 One gene of interest relevant to obesity belongs to the family of melanocortin receptors 11 ; these receptors bind four ligands as follows: α-, β-, and γ-melanocyte-stimulating hormone (α-, β-, γ-MSH) and the adrenocorticotropic hormone. 12 Disturbances in the leptin-melanocortin axis are likely to cause dysregulation in energy intake and expenditure. 13 For instance, the melanocortin-4 receptor (MC4R) gene is among the common genetic causes of obesity. Mutations in MC4R gene have been widely reported in association with early-onset obesity, hyperphagic eating behaviors, decreased energetic expenditure, and increased fat mass. 14 While substantial research in this field has been targeted mainly toward the MC4R gene, focus on the rest of melanocortin receptor genes has been less extensive with regards to childhood obesity, but a growing number of the melanocortin-3 receptor (MC3R) gene polymorphisms and mutations have recently been reported in support of the correlation with body weight.15,16
Specifically, the MC3R gene is a 7-transmembrane G-protein coupled receptor that regulates several biological functions, 17 including energy homeostasis, energy storage, and the ability to convert food into adipose tissue.12,18 As of yet, the reported polymorphisms and mutations are quite widely spread across the coding and noncoding regions of MC3R gene. 16 For example, two common missense variants, Thr6Lys (T6K) and Val81Ile (V81I), that respectively change amino acids of MC3R, are significantly correlated with increased adiposity in childhood, greater body and fat mass, and higher insulin and leptin levels. 19 Some other rare mutations in MC3R have been sporadically reported in obese cases. 16 In vitro studies have shown diverse functional impairments caused by these mutations, including decreased synthesis and/or increased degradation of MC3R transcripts, reduced cell surface expression, defective ligand binding, and impaired signaling. 15
The prevalence of most MC3R mutations is generally low in humans, so studies reporting these rare variants have limited data for a meta-analysis. 16 In contrast, T6K and V81I are two of the most common variants among MC3R polymorphisms and thus were selected since sufficient data were available to conduct this meta-analysis. However, with the exception of pathological mutations that significantly change molecular functions of MC3R, prior studies on T6K and V81I polymorphisms have generally been limited by the small size of recruitment and thus presented an inconclusive association with childhood obesity.20–22 Therefore, the current meta-analysis aimed to examine and synthesize evidence on the association between these two common MC3R polymorphisms and the development of childhood obesity.
Materials and Methods
Search Strategy
A combination of words relevant to the research question was searched on PubMed, EMBASE, Scopus, and the Cochrane database, using keywords such as “MC3R” OR “melanocortin-3 receptor” AND “polymorphism” OR “variations” OR “T6K” OR “V81I” AND “children” AND “obesity” OR “overweight” OR “diet.”
Inclusion and Exclusion Criteria
Inclusion criteria of studies were that the study must be published in the searchable database, match the relevant specifications as per the research question, and be case–control design related to the incidence of polymorphisms in MC3R gene among overweight/obese subject controls with normal weight. The results were restricted to human studies, and studies were considered if participants were between the ages of 0 and 19. Authors of studies were contacted to obtain any unpublished or additional data. Articles were then excluded based on the accessibility of full online texts and availability of pertinent data, including separate data for T6K and V81I. When considering exclusion, one review author made the initial decision, while two others checked independently. Quality of the studies was based on several criteria, including assessment of internal and external validity, study design, and risk of bias. 23 The study population was evaluated on how clearly it was specified and defined, if participants were recruited from similar populations, and if the same inclusion and exclusion criteria were used for all participants.
Methods for outcome measurements (i.e., BMI), as well as genotyping of the MC3R polymorphisms, were also examined to determine their validity and reliability. Review authors also conducted a risk of bias assessment, particularly a bias resulting from a nonrandom distribution of participants. The quality of the discussion and limitations was also assessed, including whether any confounding variables were acknowledged in the studies and if an evaluation of their impact on the relationship between obesity and MC3R polymorphisms was conducted.
Studies were organized based on the focus of the report and their relevance to the research question. The articles were read by two individuals, and the following information was extracted from each study: authors, year of publication, population, methodology, and findings. The data were subsequently analyzed and summarized.
Statistical Analysis
Pooled odds ratios (ORs) and 95% confidence intervals (CIs) were calculated using a random effects model in STATA to determine the association of the polymorphisms with obesity. The results were considered statistically significant based on the Z test (p < 0.05), with a p value <0.1 being considered in-homogeneous, and the I2 statistic was calculated to determine heterogeneity among included studies. 24 We further used STATA glst command to calculate the generalized least squares trend estimation of pooled dose–response of per K-allele at T6K and per I-allele at V81I loci.
Results
Characteristics of the Studies
A flow chart describing the process of study inclusion and exclusion is displayed in Figure 1. Searches on the databases using the keywords identified 65 potentially relevant reports. All articles were screened against the inclusion and exclusion criteria until all the final studies were chosen. A total of 32 studies were excluded due to irrelevance. Of the remaining 33 studies, 4 articles were excluded since they were literature reviews and in vitro studies, 5 excluded due to lack of access, and another 3 for investigating polymorphisms other than T6K and V81I. Nine studies were relevant to the association between MC3R polymorphisms and obesity but their sample populations involved adults rather than children and, thus, were also excluded. The total number of relevant studies was 12; however, an additional 7 studies were excluded, as they did not provide sufficient data for genotypes and BMI values of participants. Therefore, a total of five studies that investigated T6K and V81I were included in this meta-analysis.19,21,25–27
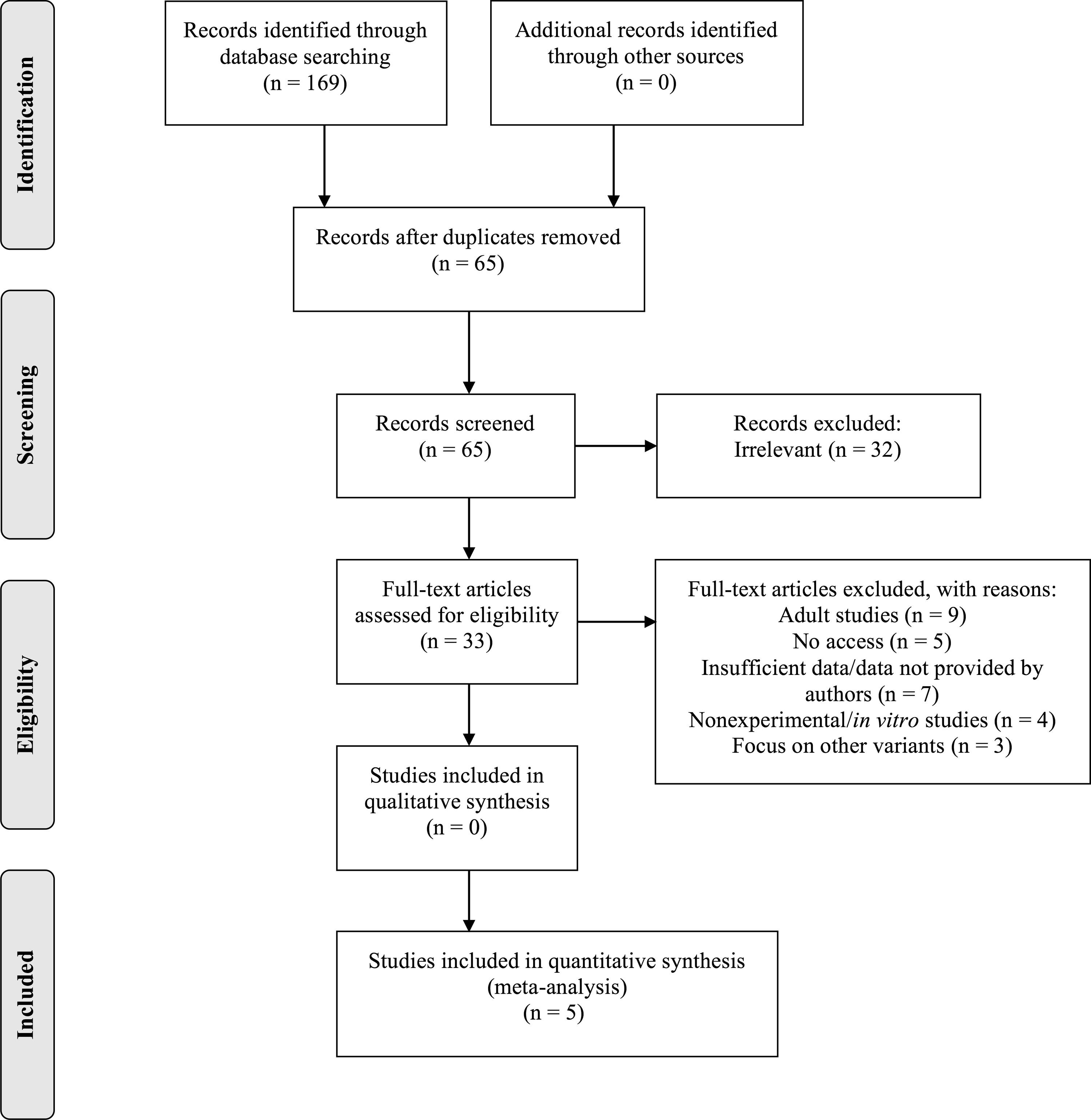
Flow chart illustrating the literature search and exclusion process.
Table 1 classifies the five case–control studies according to author, population, study methodology and design, and results. The overall population was of mixed ethnicities, including one European, 26 one Asian, 25 one South African, 21 and two American studies.19,27 The definition for overweight/obesity varied among these studies. Some studies defined the overweight/obese cases by their BMI values greater than the 95th percentile for age and sex,19,25,27 while the others used the International Obesity Task Force criteria 21 or relative BMI ratios that defined obesity by a ratio of BMI over age- and gender-standardized BMI greater than 120%. 26 For the purpose of synchronizing the outcome measurement, we combined the cases with overweight and obesity categorized according to the original definition.
Characteristics of the Included Case–Control Studies
MC3R, melanocortin-3 receptor; T6k, Thr6Lys.
Meta-Analysis Results
A total of 1118 overweight/obese cases and 1525 controls were included in the meta-analysis. The allele frequency was 0.18–0.42 for K allele at T6K and 0.1–0.44 for I allele at V81I across the studies; this rate was compatible with the Hardy–Weinberg equilibrium in each individual cohort. Heterozygous and homozygous data, analyzed separately for T6K (Fig. 2) and V81I (Fig. 3) polymorphisms, were compared with wild types to determine the relationship between genotyping in MC3R polymorphisms and childhood obesity. As shown in Table 2, when comparing with wild types (TK vs. TT and VI vs. VV), heterozygous genotypes for T6K and V81I resulted in pooled ORs of 1.14 (95% CI: 0.86–1.51) and 1.23 (95% CI: 0.91–1.66), respectively. The pooled ORs for homozygous genotypes, when comparing with wild types (KK vs. TT and II vs. VV), for T6K and V81I, were 3.10 (95% CI: 1.29–7.43) and 2.21 (95% CI: 0.73–6.72), respectively. We also analyzed the associations as a per-risk allele increase and found that the risk of overweight/obesity increased by 46.1% per K allele and 21.7% per I allele, respectively (Table 2). There was significant heterogeneity for the homozygous genotypes (I2 were 79.9% for KK and 75.1% for II).
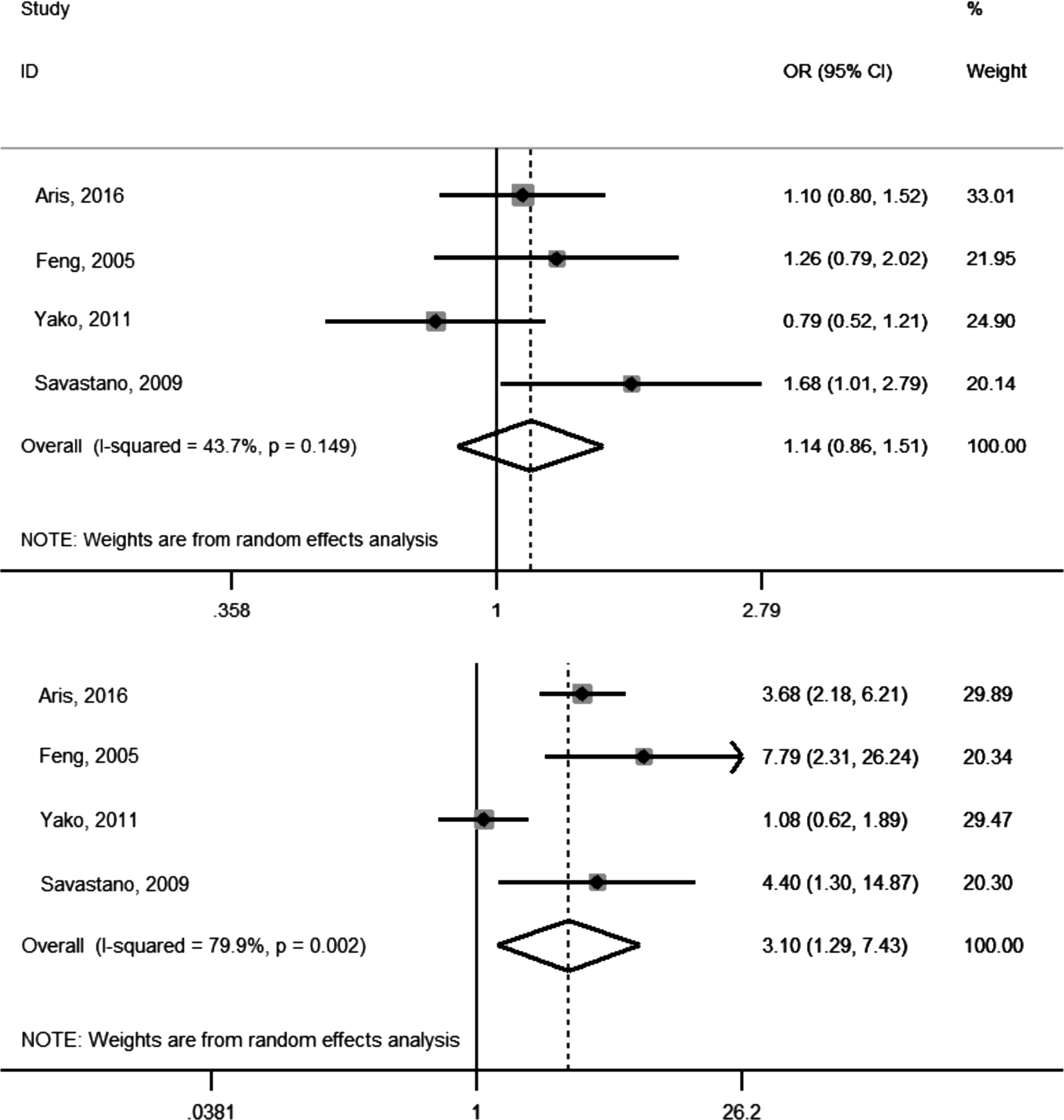
Forest plot of T6K studies describing the association between obesity and heterozygous genotype (top) and homozygous genotype (bottom). OR, odds ratio; CI, confidence interval.
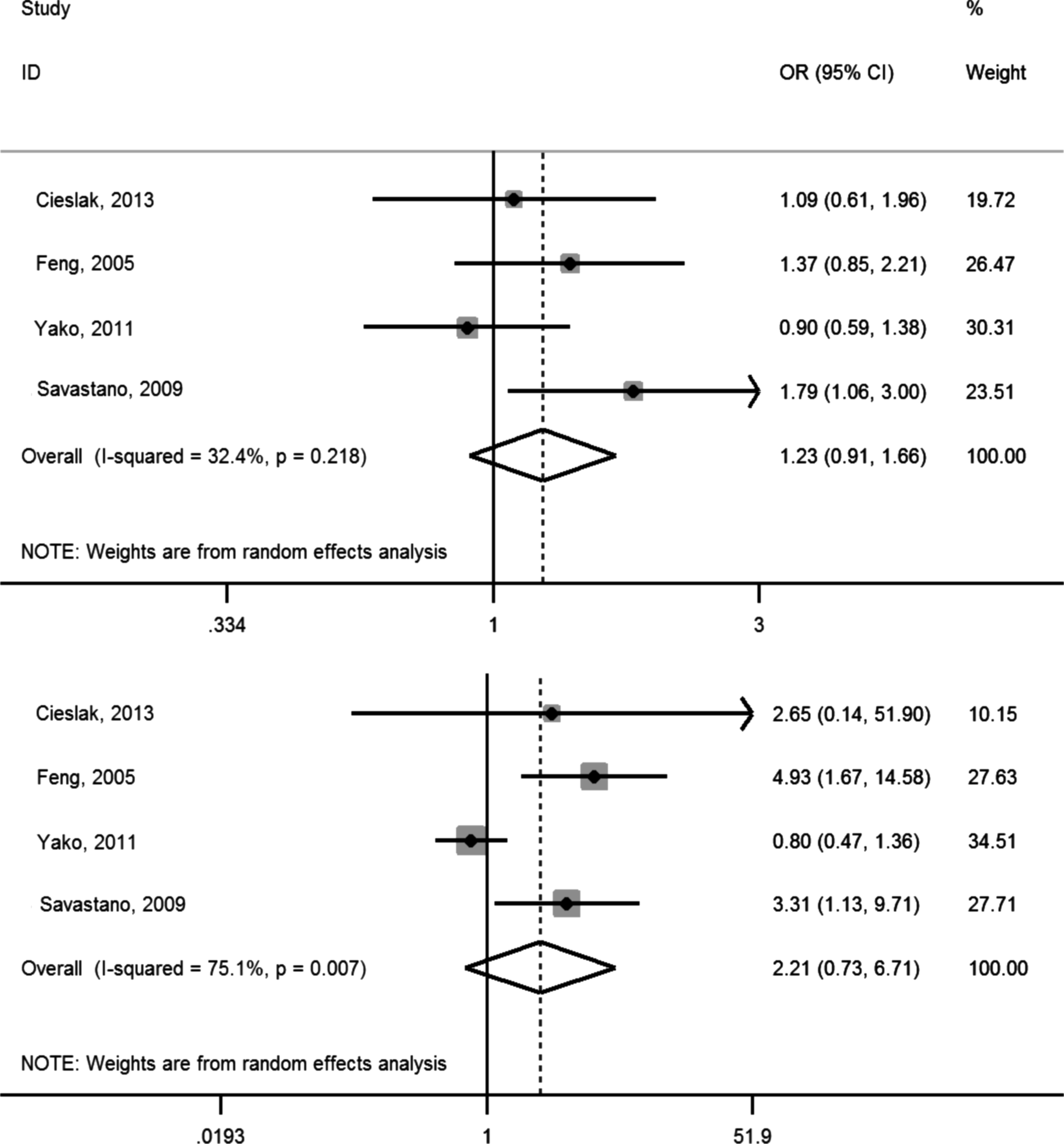
Forest plot of V81I studies describing the association between obesity and heterozygous genotype (top) and homozygous genotype (bottom).
Results of the Meta-Analysis
OR, odds ratio; CI, confidence interval; d.f., degree of freedom.
Discussion
To the best of our knowledge, this is one of the first meta-analyses investigating the association between MC3R polymorphisms and obesity in pediatric populations. In an additive model, we demonstrated a clear dose–response relationship between the number of risk alleles at both T6K and V81I loci and the risk of overweight/obesity. In the analysis stratified by genotypes, the results indicated that homozygous genotypes for T6K were associated with a 3.10-fold increased risk of overweight/obesity in children. However, this association was not significant in this pooled sample of children with homozygous V81I alleles. The heterozygotes of either T6K or V81I were not associated with overweight/obesity. Taken together, our results from the analysis per genotype supported a recessive inheritance model for MC3R gene as the cause of obesity.
The mechanisms through which MC3R affects obesity have not been fully identified, although it has been reported that the T6K and V81I mutations affect receptor function, particularly in the transmembrane region involved in the binding of melanocortin peptides. 15 Functional studies have demonstrated that double T6K and V81I mutant MC3Rs, but not single mutants, significantly decrease binding capacity for α-MSH, as well as reduced intracellular cAMP generation. 27 The decrease in ligand binding capacity, despite normal binding affinity, suggests that the presence of both T6K and V81I may cause decreased translation or increased protein degradation.16,27 In addition to in vitro studies, an animal model has shown that homozygous knock-in mice carrying double-mutant human T6K and V81I MC3R exhibited greater fat mass and feeding efficiency. 28 In humans, the double homozygosity of T6K and V81I variants, as these two polymorphisms were shown in linkage disequilibrium, has been associated with higher BMI, higher body fat mass, greater leptin and insulin secretion, and greater insulin resistance. 27 However, no significant difference in genotypic frequencies was detected among obese and control groups in another Singaporean study. 29 Further research of a larger study population may be needed to examine the potential additive effect of co-occurring polymorphisms on weight status.
One possible mechanism through which MC3R increases adiposity is related to its role in regulating energy homeostasis, specifically by increasing food intake and feeding efficiency. 30 Yang et al. suggested another mechanism, which is that levels of α-MSH, a product of MC3R, are reduced by the agouti signaling protein, an antagonist of MC3R, leading to increased food intake. 11 One study reported that fatty acid oxidation was significantly reduced in the skeletal muscle of MC3R-deficient mice, leading to an imbalance in oxidation and fat intake; this impacts feed efficiency and results in calories being more readily stored as fat.15,31 In MC3R-deficient mice, nutrients are partitioned into fat rather than muscle mass, resulting in changes in body weight. 16 These mice also exhibit adipocyte hypertrophy and increased brown adipose tissue size, due to white adipocyte infiltration, which may contribute to increased fat mass. 32 In contrast, although You et al. reported that MC3R leads to increased body fat through inhibition of energy storage, their study also suggested that MC3R by itself does not play a dominant role in lipid metabolism; rather, its effects may be due to interactions between multiple melanocortin-receptor genes, particularly MC4R. 33
In terms of eating patterns, only two of the five studies investigated the relationship between dietary patterns and MC3R polymorphisms. Savastano et al. reported that under controlled conditions, homozygous children for both variants had significantly greater energy intake in comparison to those with wild-type alleles, 19 suggesting that obesity is due to changes in the regulation of eating behavior. In contrast, Aris et al. reported slowness in eating at 12 months in obese boys with MC3R variants 25 ; although this is contrary to studies that report higher energy intake in those with MC3R polymorphisms, it is possible that children may be consuming more food but taking a longer time to finish eating. 25 Furthermore, heterozygous and homozygous individuals with T6K and V81I were reported to have elevated free fatty acids, low rates of lipid oxidation, and decreased insulin to glucose ratio and fasting glucose due to high rates of glucose oxidation. 34 Another study involving a weight-loss program found that the co-occurrence of these polymorphisms in heterozygous children resulted in an inability to increase feeding efficiency and fatty oxidation, leading to difficulties with weight loss. 35 However, these findings were somehow inconsistent with our negative results of the association between heterozygous genotypes and occurrences of overweight/obesity. This may be explained by the possible presence of obesogenic factors that were not examined in the current analysis. For example, lifestyle factors such as lack of physical activity and unhealthy diets, as well as environmental conditions, including availability and access to healthy foods, 36 were not adjusted in the comparison. Thus, the percentages of obesity attributable to heterozygous genotypes in these two MC3R polymorphisms may be outweighed by these unmeasured risks.
Our analysis was limited by several factors. First, the working definition for childhood overweight and obesity is a critical part of a meta-analysis and can substantially affect the results. We found varying definitions of overweight and obesity among the studies that were included in this meta-analysis. One study defined overweight/obesity based on relative BMI values greater than 120% of standard BMI within the study cohort, 26 which was different from the others using the cutoff values of the population data.19,21,25,27 For our analysis, we chose to combine the number of overweight and obese patients. We also back-calculated data for one of the chosen studies from supplementary graphs, due to lack of individual data. 25 Significant heterogeneity among the included studies may indicate the presence of other unidentified sources of uncertainties, such as limited numbers of pooled subjects and ethnic differences, and may be partly compensated for using a random-effect model to estimate the association. Individual studies had small sample sizes, thus making it possible that the association between the variants and obesity was overestimated due to population stratification, as a reporting bias may have been introduced to support positive results. 19 In addition, Aris et al. reported a low response rate, only 40%, for their Child Eating Behavior Questionnaire, which was used to measure the relationship between slowness in eating and adiposity. 25 Their conclusion on this association is limited due to outcome data being largely unavailable. Furthermore, their use of this self-reported questionnaire may further elevate the risk of bias. Second, the overall number of subjects was relatively limited for a genetic association study, despite our extensive efforts in searching for matched articles. While all relevant articles were inspected and quality assessed, exclusion of studies was based primarily on the availability of required data. 22 Several articles were excluded because only obese subjects were genotyped in the search for causative obesity genes. 20 Some other studies combined data for T6K and V81I, as these two polymorphisms were shown in linkage disequilibrium.27,29 However, not every study included in this meta-analysis concurrently examined the allele frequency at both loci, which makes the examination of potential additive effect of co-occurring polymorphisms beyond the scope of this study. Since we obtained different results for these two polymorphisms, they may not necessarily contribute equally to the occurrence of obesity and thus require further research to verify.
Conclusion
In summary, this meta-analysis confirmed a significant association between childhood obesity and the homozygous genotypes of T6K polymorphisms in the MC3R gene. Future directions for research in this area include using standardized definitions for obesity to reduce heterogeneity and conducting studies with specific focus on weight trajectory in pediatric populations. Studies examining the effects of interaction between different polymorphisms within MC3R gene or between the genes coding for different subtypes of melanocortin receptor in relation to obesity and eating patterns should also be taken into consideration to broaden our knowledge about how genes lead to early-onset obesity and ways to manage childhood weight problems. Identifying the genetic cause of childhood obesity may provide an insight into the development of pharmaceutical treatment. From a clinical perspective, rigorous physical activity and restrictive dietary practice should be indicated in obese individuals with obesogenic MC3R alleles, before an effective MC3R antagonist is proved to be of clinical efficacy.
Footnotes
Acknowledgments
This study was funded by the Summer Undergraduate International Research Internships Scholarship awarded to Koya C from the University of Ottawa and partly by the Medical Science and Technology Research Grant awarded to Tsai MC from the National Cheng Kung University Hospital (NCKUH-10602007).
Author Disclosure Statement
No competing financial interests exist.