
Abstract
Background:
Obesity in children continues to be a global epidemic. Given the recent focus on the potential role gut microbiota play in the development of obesity and the continuous rise of obesity in childhood worldwide, it has become necessary that the association between gut microbes and weight is explored in previously unexplored regions such as the Caribbean island of Trinidad. As such, our objective was to characterize the composition of the gut microbiota in children with obesity/overweight and children of normal weight to determine if there are any gut microbes associated with overweight and obesity.
Methods:
Fifty-one children provided stool samples and their BMIs were calculated and classified. 16S amplicons from stool samples were individually barcoded and sequenced in multiplex in the NextSeq 500 platform in a 150 bp paired-end modality and a complete gut microbiome profile was generated.
Results:
The relative abundance and diversity of the microbiota of the lean and children with obesity/overweight were analyzed against each other and showed that children with obesity/overweight recorded significantly more (p = 0.001) Firmicutes and significantly less Bifidobacterium (p = 0.039) than the children of normal weight. Children with obesity/overweight also recorded significantly (p = 0.009) reduced alpha diversity compared with the lean children.
Conclusion:
This study has shown associations of Firmicutes to obesity/overweight and Bifidobacterium species with healthy weight in children.
Introduction
There is a global epidemic of obesity in childhood. The World Health Organization (WHO) has shown that there has been a 33% increase in the prevalence of obesity in childhood between 1990 and 2016. 1 As more countries adopt a westernized high-caloric diet, obesity in their populations also increases. 2 Children who affected by overweight/obesity are prone to become adults who are also affected by obesity 3 and are at increased risk for serious diseases such as cardiovascular diseases, type 2 diabetes, and cancer. 2 While an increased intake of energy dense foods and less physical activity among children contribute to overweight/obesity, there is also growing awareness of variation in the gastrointestinal microbiota of children with obesity compared with those of normal weight. Specifically, research has shown individuals prone to obesity have higher concentrations of gut microbes that are more efficient at extracting energy from food than those in nonobese individuals; therefore, contributing to excessive caloric uptake compared to expenditure. 4 The human gastrointestinal tract contains a diverse and abundant microbial community, Taxonomically, these microorganisms are divided into specific categories which are phyla, classes, orders, families, genera, and species.5,6
After birth, the intestinal tract of infants moves gradually from sterility to dense microbial colonization within the first 3 years of life. Once an equilibrated colonization with commensal bacteria is formed, it prevents invasion and colonization with pathogens and helps in immune modulation, nutrient metabolism, host physiology, and the maintenance of the gut barrier.7,8 However, disruption to this balance, or dysbiosis, has been linked to the pathogenesis of many diseases, including obesity, metabolic syndrome, diabetes, asthma, atherosclerosis, and irritable bowel syndrome.9,10
How the gut microbiota composition might lead to the development of obesity has been postulated to be due to three mechanisms. First, gut microbes could affect the levels of lipopolysaccharide in the blood, which would create systemic chronic inflammation resulting in a predisposition to developing obesity and diabetes. Second, microbes can extract energy from indigestible polysaccharides, thereby providing extra calories to the host. Third, gut microbes can also regulate host genes associated with the consumption and storage of energy.11,12 The relative abundance of specific bacterial phyla has been shown to be more common in obese animals and humans. An elevated Firmicutes-to-Bacteroides ratio has been shown in obese leptin-deficient mice and in children with obesity when compared with lean counterparts.13,14
In Trinidad, a high-income country, serial studies of obesity among Trinidadian school-aged children showed that the prevalence of obesity in this population had risen from 5% in 2001 to 15% by 2010. While the authors of these studies did not specifically investigate the causes of this significant increase, they did suggest that lifestyle interventions such as increased physical activity and dietary changes were necessary.15,16 As far as we are aware, there has been no study of the microbiota in school-aged children with and without obesity in Trinidad to determine if postulated variation in microbiota phyla and diversity, noted in other parts of the world, are equally applicable to this Caribbean island. Indeed, there is limited information on microbiome diversity in Caribbean populations. Therefore, this study is the first to characterize the composition of the gut microbiota in children with and without obesity in any Caribbean island, specifically Trinidad, and we hypothesized that given the increase in American-style fast foods and other high-caloric foods now widely consumed in this island that children with obesity/overweight would have a higher relative abundance of Firmicutes and less Bacteroidetes than children without obesity, as seen in other westernized societies.
Methods
Subject Enrollment Criteria
The study proposal was approved by Ethics Committees of The University of the West Indies, St. Augustin Campus, and the North Central Regional Health Authority (NCRHA). Fifty-one school-aged children (6–14 years) who attended either of two health facilities, the Eric Williams Medical Sciences Complex (EWMSC) or the Arima Health Facility, for management of allergies, were recruited at random. These children were healthy, apart from their allergies, and were specifically excluded if there was a history of gastrointestinal infection or diarrhea; had taken antibiotics within the prior 3 months; were using a probiotic, or had a diagnosis of immunodeficiency. Signed consent was obtained from parents and participants.
BMI Measurement and Classification
Study participants height and weight measurements were taken for BMI calculations. The BMI formula used was weight (kg)/height (m2) and patients were classified into lean, overweight, obese using WHO Child Growth Standards Curves. Children with BMI between the 85th and 97th percentile was deemed overweight and greater than the 97th was classified obese. Children who were classified based on their percentile as being overweight and obese were grouped as one category against the children of normal weight.
Sample Collection
Study participants were each provided with a uBiome (California) kit and collected their stool following the instructions contained (www.uBiome.com). The Kits contain a stabilizing fluid that preserved the sample in time for processing.
Sequencing of Stool Samples
16S amplicons from stool samples were individually barcoded and sequenced in multiplex in the NextSeq 500 platform in a 150 bp paired-end modality. Raw data from the sequencer was first demultiplexed, and the forward and reverse reads obtained in each of the four lanes per sample were filtered as follows; both forward and reverse reads in a pair must have an average Q-score >30; primers, and any leading random nucleotides (used to increase diversity of the library being sequenced) were trimmed, and forward reads were capped at 125 bp and reverse reads were capped at 124 bp; forward and reverse reads of each pair were appended, and those sequences that contain more than eight consecutive same nucleotide repeats were discarded; the remaining sequences were clustered using a distance of one nucleotide using the Swarm algorithm1, 17 and the most abundant sequence per cluster was considered the representative of the cluster and it was assigned a count corresponding to the sum of sequences that compose the cluster; a chimera removal using these centroid representative sequences was performed using the VSEARCH uchime_denovo algorithm2. 18 Singletons that remain after chimera removal were also discarded. Finally, both forward and reverse reads that match with at least 77% sequence identity to the same sequence in version 123 of the SILVA database3 19 were assumed to be 16S sequences. The most abundant forward-reverse read pair per Swarm cluster was assigned taxonomic annotation according to similarity thresholds. If the best hits for the forward and reverse reads have >97%, >95%, >90%, >85%, >80%, and >77% identity to the same sequence in SILVA, the reads were annotated to the same species, genus, family, order, class, and phylum of the hit in SILVA, respectively.
Statistical Analyses
Differential abundance
Differential abundance analysis was done to compare the microbial population among samples representing multiple conditions. A prevalence filter was first conducted whereby only dominant taxa present in samples were included in the analysis. The p-values were calculated using an analysis of variance test of each model after transformation.
Alpha diversity comparison
Metrics of alpha diversity 20 were compared among groups. First, normal distribution of data was tested by the Shapiro–Wilk test for normality. Then, pairwise comparisons were done using the Wilcoxon rank-sum test for nonparametric data or T-tests when the data displayed a normal distribution. Chi square statistics analyzed the relationships between categorical variables from the Global Asthma Network questionnaires completed by participants. Analyses were performed by using IBM SPSS software Ver. 21 and p-values <0.05 were considered significant.
Results
Forty-five percent of the study participants were between the ages of 6 and 10, with 55% between the ages of 11 and 14. There were more male (59%) than female (41%) participants. Of the three self-reported ethnicities, most were Mixed (65%), followed by Africans (31%), with Indians recording the least at 4%. Forty-one percent of the children were overweight/obese, with 59% being of normal weight (Table 1).
The Relative Abundance (%) of the Most Dominant Bacterial Phyla from Stool Samples at Varying Demographic and Anthropometric Measure
The main bacterial phyla detected in the complete cohort (Fig. 1a) were, in order of highest relative abundances, Firmicutes (61.15%), Actinobacteria (26.28%), Bacteroidetes (5.76%), and Proteobacteria (2.73%). Children between the ages of 6 and 10 had more Actinobacteria, Bacteroidetes, and Proteobacteria than those aged 11–14 years, but the differences did not achieve significance. Higher relative levels of Firmicutes, compared with the other phyla assessed, were a significant finding (p = 0.001) in children with obesity/overweight. There were no detectable differences in bacterial phyla among children of different ethnic groups and gender (Table 1).
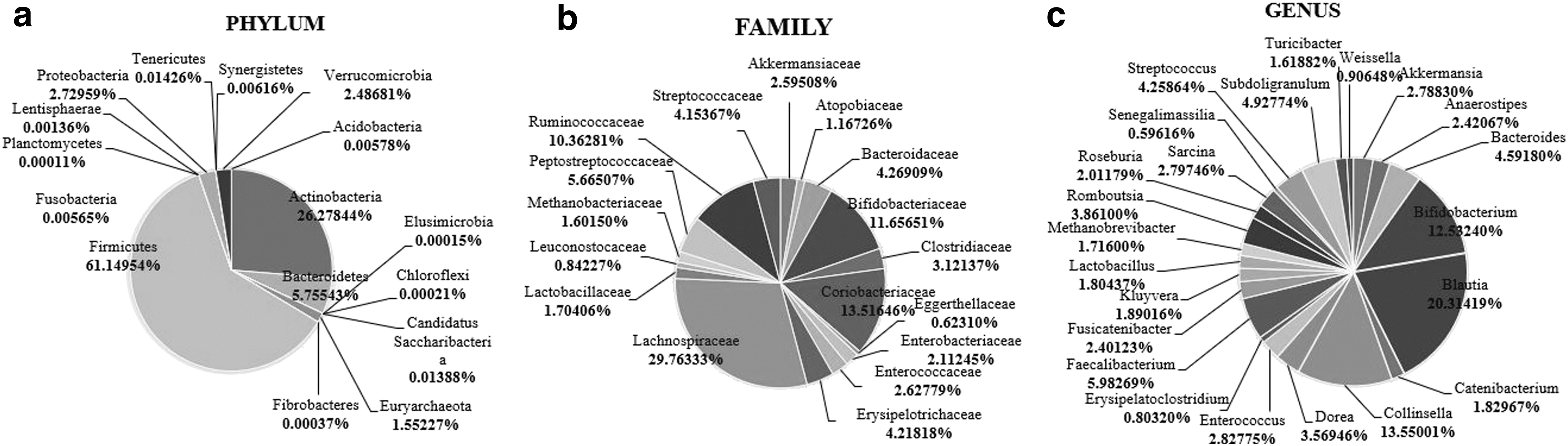
Aggregated microbiota composition of all children in study cohort at the phylum
The dominant bacterial families were Lachnospiraceae (29.76%), Coriobacteriaceae (13.51%), Bifidobacteriaceae (11.66%), and Ruminococcaceae (10.36%) (Fig. 1b). There were gender differences in the relative proportions of these bacterial families, with female participants having more Lachnospiraceae (p = 0.002) than males. Children with obesity/overweight were also found to have significantly less Bifidobacteriaceae than those of normal weight (Table 2). There were no significant differences between the dominant bacterial families and age and ethnicities.
The Relative Abundance (%) of the Most Dominant Bacterial Families from Stool Samples at Varying Demographic and Anthropometric Measure
At the level of bacterial genera (Fig. 1c), the dominant genera were Blautia (20.31%), Collinsella (13.55%), Bifidobacterium (13.53%), and Faecalibacterium (5.98%). Females recorded significantly more (p = 0.038) Blautia than males, and children with obesity/overweight had significantly more Lactobacillus and significantly less Bifidobacterium than those of normal weight (Table 3). There were no significant differences between the dominant genera and age and ethnicities. When phylogenetic diversity (rooted) was explored (Fig. 2) there were significant differences (p = 0.009) between the normal weight children and children with obesity/overweight, with the latter group having less phylogenetic diversity in their gastrointestinal microbiota.
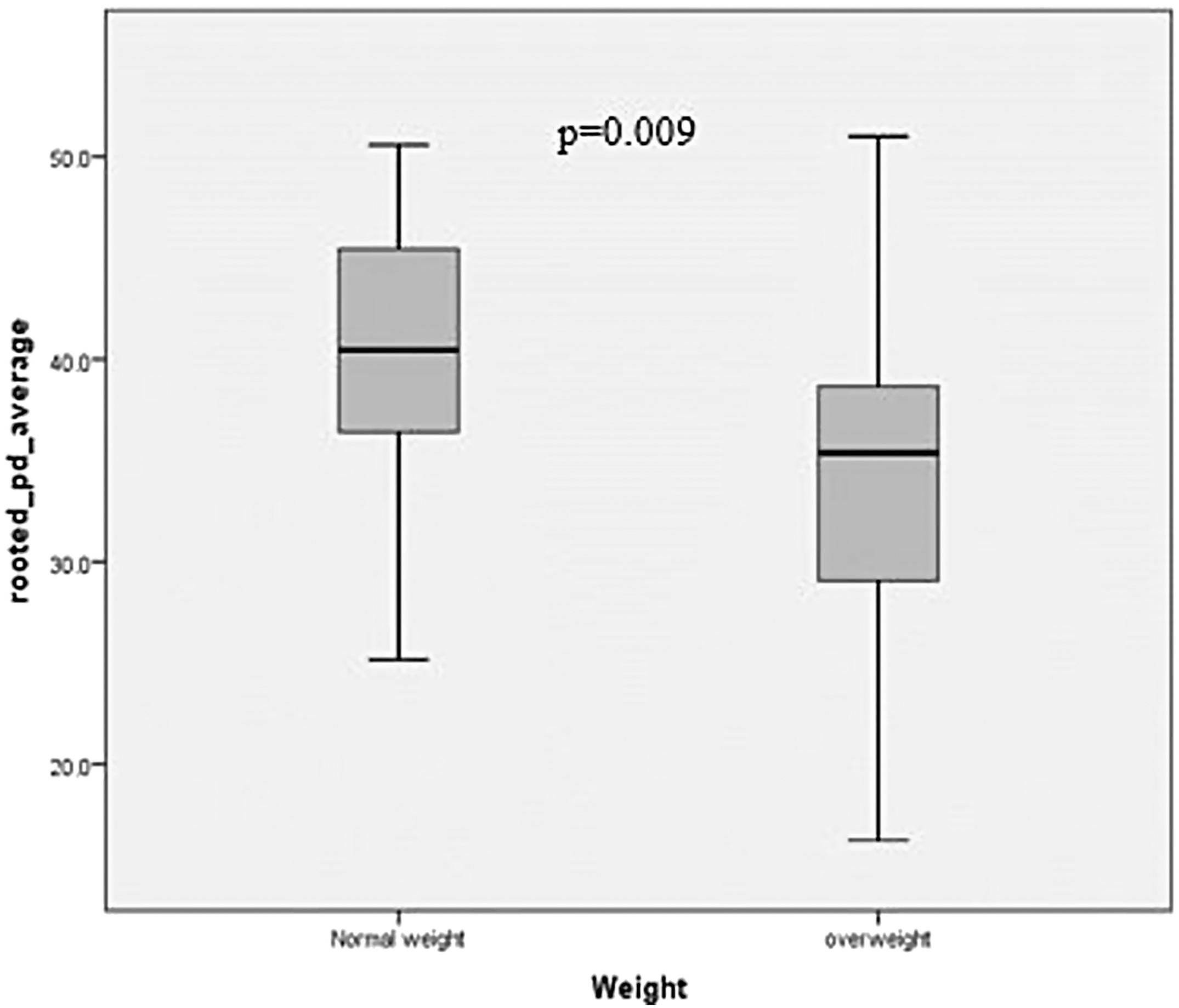
A metric of alpha diversity of differences between children with and without obesity; p = 0.009.
The Relative Abundance (%) of the Most Dominant and Applicable Bacterial Genera from Stool Samples at Varying Demographic and Anthropometric Measure
Discussion
We hypothesized in this study that children with obesity/overweight growing up in Trinidad, a high-income island in the Caribbean, with a rapidly increased prevalence of childhood obesity,15,16 would have gastrointestinal microbiota compositions similar to those seen in other high-income countries. In addition, this hypothesis was based on the observation that in the Caribbean and Latin America, there has been a general reduction in consumption of traditional meals and an increased consumption of ultraprocessed products, 21 and in Trinidad and Tobago the costs of fruit and vegetables have become too expensive for many parents to give their children healthy meals. 22 Further, to date, there has been no comparative study of the microbiota in overweight/obese and children of normal weight in Trinidad and Tobago and, as far as these authors are aware, in the English-speaking Caribbean.
The gastrointestinal bacterial composition in fecal samples from a cohort of 51 children were analyzed at 3 taxonomic levels; phylum, family, and genus to determine if there were differences in relative abundance of gut bacteria in relationship to participants weight (overweight/obese versus lean), age, gender, and self-reported ethnicity. Our findings show that children with obesity have more Firmicutes, and that there was a positive linear correlation between Firmicutes and BMI. It was also noted that children with obesity had lower levels of Bacteroidetes and Bifidobacteria, and that there was a negative correlation between Firmicutes and Bacteroides. The increased abundance of Firmicutes in children with obesity in our study cohort is consistent with reports from other parts of the world.23,24 The presence of this phyla has been noted in mouse studies to correlate with a diet of high sugar and fat 25 as would be found in diets rich in ultraprocessed foods, which have become more prominent in local diets in Trinidad and Tobago.
Most studies showed a corresponding decrease in Bacteroidetes in obese individuals.26,27 Although not significant, children with obesity had less Bacteroidetes than the children without obesity in this study. In addition, Firmicutes had a significant negative linear relationship with Bacteroidetes. An increase in the ratio of Firmicutes to Bacteroidetes may contribute to obesity as it is associated with increased energy extraction from colonic fermentation and short chain fatty acids.13,28 Children with obesity/overweight from this study had 16 times in amount of Firmicutes than Bacteroidetes.
At the family level, only Bifidobacteriaceae showed significance. Children of normal weight had significantly more Bifidobacteriaceae than children with obesity/overweight. While at the genus level, the related Bifidobacterium showed similar association. A study showed that Bifidobacterium Species reduced body and fat weights, harmful enzymatic activity, and blood serum levels in Sprague-Dawley rats fed high-fat diet. 29 This suggests that Bifidobacterium spp. may have antiobesity and lipid lowering capabilities.
We found lactobacillus to be significantly more in children with obesity/overweight than the lean participants. Lactobacillus has been used as a probiotic to promote growth in farm animal. Specifically, animals treated had significantly greater weight gain and food conversion efficiency than controls. 30 Similarly, in humans, lactobacillus was found to be in significantly higher concentrations in patients with obesity than those without obesity. 31
We also noted lower bacterial diversity in the fecal samples from children with obesity/overweight compared with those of normal weight. This is of concern as reduced microbial diversity has been associated with obesity, insulin resistance, impaired glucose homeostasis, and inflammation 32 and has been linked to an altered representation of metabolic pathways and bacterial genes. 25 A species rich gut microbiota is important as they are more adaptive to environmental influences and more resistant to diseases. The association between reduced bacterial diversity and chronic disease development is concerning in the context of Trinidad as the island already has one of the highest prevalences of diabetes in the western hemisphere at 13% of its population. 33
The strong association of Lachnospiraceae and Blautia with female gender in our study is interesting. Both Lachnospiraceae and Blautia have been linked to a number of chronic diseases, including depression, diabetes, and lupus. Org et al., 34 found that members of the family Lachnospiraceae were more abundant in female mice, but the significance of this finding remains obscure.
There were some limitations to this study. Although stool sampling is a relatively simple and noninvasive procedure; the stool collected may not fully reflect the environment that exists on the wall of the gut, where microbiome host interactions are expected to take place. In addition, most nutrients are absorbed in the small intestines where it is difficult to obtain a sample. Finally, analyses were done on the most dominant bacteria, however, there exists the possibility that microbiota found in small quantities may be playing a role in adiposity.
Conclusions
This study is the first to investigate the composition of the gut microbiota of children in Trinidad and provided entire characterization of the relative abundances of all the microbiota at the phylum and most at the family and genera levels. With rising obesity rates in children in Trinidad, it has become important that new etiologies are explored in regions it was previously unexplored. This study found that a combination of gut microbes may be reflective of obesity/overweight in children. Children with obesity/overweight recorded significantly more microbes that are associated with adiposity and a reduced abundance of microbes shown to help in healthy weight. In addition, children with obesity/overweight had a less adaptive gut microbiome reflective in reduced diversity. Whether this gut microbial dysbiosis is reflective of a more westernized diet adopted by children in Trinidad is open to further interpretations, however, it leaves room for possible therapeutic interventions such as the use of beneficial microbes (pro- and prebiotic) to establish balance of gut microbes and possibly assist in weight loss.
Footnotes
Acknowledgments
Special thanks to the Preclinical department of the Faculty of Medical Sciences, Dr. George Legal who assisted with data analyses, and Ms. Samantha Jagmohan for subject recruitment.
Funding Information
Author Disclosure Statement
No competing financial interests exist.