
Abstract
Abstract
1. Introduction
There are many formulations modeling the behavior and interaction of the lac genes. The first model was proposed by Goodwin two years after the discovery of the lac operon (Goodwin, 1963). Since then there has been a steady flow of models following the advances in biological insight of the system, with the majority describing operon induction using artificial nonmetabolizable compounds such as IPTG and TMG (Ozbudak et al., 2004; van Hoek and Hogeweg, 2006, 2007; Santillán et al., 2007). The first model to consider catabolite repression and inducer exclusion with cells grown in both glucose and lactose and lactose used as the inducer was presented by Wong et al. (1997). Their model consisted of up to 13 ordinary differential equations involving 65 parameters. Further, Santillán and coauthors have presented mathematical models and analysis purporting bistability in that the operon is either induced or uninduced (Yildirim et al., 2004; Santillán and Mackey, 2004; Santillán et al., 2007; Santillán, 2008), which has been observed experimentally (Novick and Weiner, 1957; Ozbudak et al., 2004). These findings have given rise to the analogy of the lac operon acting as a biological switch (Ozbudak et al., 2004; Halász et al., 2007).
Most mathematical models of the lac operon, as well as other genetic systems, are given as systems of differential equations which are appropriate for understanding mechanisms. However, discrete modeling frameworks are receiving more attention in systems biology for their use in analyzing entire state spaces and facilitating global-level predictions. In fact it has been argued that network topology and interaction type (activation/inhibition) are sufficient for capturing dynamics of gene networks (for proofs of concept in other model organisms, see Albert and Othmer, 2003; Mendoza and Xenarios, 2006). A consequence of this observation is that the use of qualitative networks eliminates the need to estimate parameters, thereby reducing model complexity.
Finite dynamical systems (FDSs), which are discrete modeling frameworks suitable for describing qualitative aspects of a network including topology, provide a computational advantage in that entire state spaces can be computed and explored analytically using methods such as generalized logical analysis (Mendoza and Xenarios, 2006; Thomas et al., 1995), in contrast to continuous modeling frameworks. The advantage of having the state space given explicitly is that features such as steady states and limit cycles can be essentially read from the state space. Such predictions, as well as others provided by FDSs, are often easier to verify experimentally than their continuous counterparts, due to their qualitative description; that is, precise experimental conditions are not required to verify qualitative predictions. Boolean networks (BNs), a special class of FDSs, permit an intuitive, yet formal mathematical description of network dynamics by encoding network topology and interaction type in Boolean expressions (Albert and Othmer, 2003; Davidich and Bornholdt, 2008).
Currently few discrete models of the lac genes exist. Gianchandani and coauthors introduced a Boolean model of the lac operon, excluding the regulatory effects of glucose and lactose and demonstrated that an alternate method for identifying all steady states using a matrix representation of the Boolean model (Gianchandani et al., 2006). Setty et al. (2003) defined a logical function, a specialization of FDSs, for the transcription of the lac genes in terms of the proteins regulating the operon, namely CRP and LacI. Although the authors initially aimed to construct a simple Boolean function to mimic the switching behavior of the operon, they discovered that AND-like and OR-like expressions could not reproduce the complexity that the lac genes exhibited. Instead they found that a logical function on four states (as opposed to two states—0 and 1) was more biologically relevant. Mayo et al. (2006) tested and showed that this logical function was robust with respect to point mutations, that is, given the formulation in Setty et al. (2003), the operon is still functional after point mutations.
One limitation in both models is that they do not predict bistability. We propose a logical model for the lac operon which predicts bistability (when stochasticity is included) and includes the two main control mechanisms of glucose, namely catabolite repression and inducer exclusion. In order to facilitate interpretation, we have added variables so as to present the model as a BN. Therefore, we are able to show that the lac operon has only two steady states, corresponding to the operon being either ON or OFF (induced or uninduced).
An important question is to identify the key players in a network, in this case for the purpose of determining the drivers of the dynamics. Aguda and Goryachev (2007) provided a systematic approach for reducing a network derived from literature to a subnetwork which can be thought of as the core of the essential qualitative behavior. Albert and Othmer (2003) showed that the topology and interaction type are the determining factors in producing the steady-state behavior. We corroborate these findings by reducing the Boolean model to one involving only the lac genes and lactose. We show that the dynamics of the reduced model matches that of the full model. Our results further support the hypothesis that the topology is the key to dynamical properties.
This article is organized as follows. In Sections 2 and 3, we provide the biological and modeling background. We present a BN as a model for the lac operon in Section 4, and we show its network topology, associated dynamics, and bistability analysis. In Section 4.2, we use a novel model reduction algorithm and show the core subnetwork supporting bistability. We close with a discussion of potential future directions in Section 5.
2. Theory
2.1. Biological background
Here we describe the components and features of the lac operon which we include in the model. This description is summarized largely from the material provided in the online book (Griffiths et al., 1999). Additional citations are given as necessary.
The lac operon contains three structural genes, lacZ, lacY, and lacA, and is a negative inducible system: the repressor protein LacI prevents transcription of the lac genes, and the operon is induced by allolactose, an isomer of lactose. Extracellular lactose is thought to be readily available, but can diffuse into the cell at low concentrations. Once in the cell, lactose can induce the operon, though with lower probability than allolactose. Transcription of the lac genes produces a single mRNA, whose translation gives rise to the following proteins: β-galactoside permease (LacY), a membrane-bound protein which transports lactose into the cell; β-galactosidase (LacZ), an intracellular protein which cleaves lactose into glucose and its stereoisomer galactose, and in a separate reaction converts lactose into allolactose; and β-galactoside transacetylase (LacA) which transfers an acetyl group from acetyl-CoA to β-galactosides.
Glucose is thought to regulate the lac operon through two key mechanisms: catabolite repression and inducer exclusion. In the absence of glucose, the catabolite activator protein CAP (also known as CRP for cAMP receptor protein) forms a complex with cAMP which binds to a site upstream of the lac promoter region. Binding of the cAMP-CAP complex makes a conformational change in the DNA, thereby allowing RNA polymerase to bind to the DNA and enhancing transcription of the lac genes. Transcription continues until extracellular glucose is available. However, when glucose is abundant, cAMP synthesis is inhibited (Wong et al., 1997) and the repressor protein LacI can bind to the operator region of the operon, preventing transcription of the lac genes. Furthermore, the transport of lactose into the cell by permease is inhibited by external glucose, a mechanism referred to as inducer exclusion. Finally, the presence of (sufficient amounts of) intracellular glucose shuts off the operon, a phenomenon referred to as catabolite repression.
2.2. Modeling background
A BN
“Network topology” refers to the connectivity structure of a network and is typically represented as a directed graph. For a BN, a wiring diagram is a directed graph on the variables of the system (in this case, mRNA, proteins, and sugars) with edges defined in the following way: there is a directed edge from variable xi to xj if the function Fj for xj depends on xi. An edge from xi to xj has a circle at its head xj if NOT xi, denoted by ¬xi, appears in Fj (we consider this edge to correspond to an inhibitory interaction); otherwise, edges have arrows at their heads. We call directed cycles feedback loops. The parity of a feedback loop (or a path) can be either +1 or −1 and is calculated as follows: assign −1 to an edge if it is inhibitory and +1 otherwise. The parity of a feedback loop is the product of the signs on the edges of the loop. If the parity of a feedback loop is +1, we call the loop positive; otherwise, it is negative.
“Dynamics” refers to the state transitions of the network as a whole. To generate the dynamics of a BN F on n variables, we evaluate its functions on all possible combinations of 0/1 n-tuples. The dynamics can be viewed as a directed graph, called the state space of F. In this graph each node is an n-tuple, called a state of the system F; there is a directed edge from a to b if F evaluated at the current state a gives state b; that is, if F(a) = b. Hence, b represents that next state of the system. Directed cycles are called limit cycles. If length of the cycle is 1, then it is called a fixed point. In the context of scientific applications, fixed points are also referred to as steady states, which we use in this discourse. We draw the state space using the visualization software DVD (Jarrah et al., 2004).
3. Methods
The Boolean network model that we present in Section 4 consists of variables and functions, each representing mRNAs, proteins and sugars. We assume that each biomolecule can be either 0 or 1 (absent/inactive or present/active). In order to perform a comparison with an ODE model we considered that extracellular lactose can be in three states (low/0, medium/1, high/2) which we model using the variables Le and Lem; they indicate high concentration and medium concentration of extra cellular lactose, resp. (Le, Lem) = (0, 0), (0, 1), (1, 1) means that the concentration of extracellular lactose is low, medium and high, resp. The same was done for lactose, allolactose and the repressor protein.
The key Boolean variables are labeled as follows:
M = lac mRNA P = lac permease L = lactose C, R = CAP and LacI, resp. B = lacβ-galactosidase A = allolactose
where the subscript m indicates medium concentration and e indicates extracellular concentrations.
3.1. Deriving Boolean networks
Here we illustrate how to derive a Boolean function for mRNA based on the information in Section 2.1. The operator ∧ indicates that both variables need to be present, that is, activation occurs by the synergistic action of its regulators; on the other hand, the operator ∨ indicates that either variable is enough for activation to occur, that is, activation occurs independently of the other variable. The operator ¬ indicates that the variable is a repressor and that it has to be absent for activation to occur. For any variable a, the function Fa determines the value of a after one time unit.
Boolean function for M: To have mRNA being transcribed we need the activator protein (CAP), but also the repressor to be absent. That is, the future Boolean value of M is given by the PRESENCE of C AND the ABSENCE of R AND the ABSENCE of Rm. Hence, the Boolean function for M is FM = C ∧ ¬ R ∧ ¬Rm.
Boolean functions for P, B: To have the lac proteins being produced we need mRNA. Hence, the Boolean function for P, B are FP = FB = M.
Boolean function for C: To have CAP in high concentrations we need glucose to be absent. That is, the future Boolean value of C is given by the ABSENCE of Ge. Hence, the Boolean function for C is FC = ¬Ge.
Boolean function for R: The repressor will be active as long as there is no allolactose. That is, the future Boolean value of R is given by the ABSENCE of A AND the ABSENCE of Am. Hence, the Boolean function for R is FR = ¬A ∧¬Am.
Boolean function for Rm: The repressor will be active at (at least) a medium level as long as there is no allolactose or if the repressor was active to begin with. That is, the future Boolean value of Rm is given by {the ABSENCE of A AND the ABSENCE of Am} OR the PRESENCE of R. Hence, the Boolean function for R is FRm = (¬A∧¬Am) ∨ R.
Boolean function for A: To have allolactose in high concentrations we need both β-galactosidase and lactose to be present. That is, the future Boolean value of A is given by the PRESENCE of B AND L. Hence, the Boolean function for A is FA = B ∧ L.
Boolean function for Am: To have allolactose in medium concentrations we need lactose to be present in at least a medium concentration (it will be transformed to allolactose by a basal number of β-galactosidase molecules which we are not including in the model). That is, the future Boolean value of Am is given by the PRESENCE of L OR Lm. Hence, the Boolean function for Am is FAm = L ∨ Lm.
Boolean function for L: To have lactose in high concentrations we need both permease and extracellular lactose to be present, but also glucose to be absent. That is, the future Boolean value of L is given by the PRESENCE of P AND Le AND the ABSENCE of Ge. Hence, the Boolean function for L is FL = P ∧ Le ∧¬ Ge.
Boolean function for Lm: To have lactose in medium concentrations we need extracellular lactose to be present in at least a medium concentration together with permease, or extracellular lactose to be present in concentrations high enough so that it will enter the cell (by diffusion or a basal number of permease molecules which we are not including in the model); we also need glucose to be absent. That is, the future Boolean value of Lm is given by the {{PRESENCE of Lem AND P} OR the PRESENCE of Le} AND the ABSENCE of Ge. Hence, the Boolean function for Lm is
We use F to denote the Boolean model comprising all of the above functions.
3.2. Bistability
To prove that F exhibits bistability we have to show that for a range of parameters a population of “cells” may exhibit both stable steady states at the same time, corresponding to the lac operon being ON and OFF (Santillán and Mackey, 2004); that is, there exists a region of bistability. We will show that if we consider stochasticity in the uptake of the inducer, then bistability can occur.
We performed experiments similar to those in Ozbudak et al. (2004). In order to do this, we considered a stochastic version of our model, and we considered glucose to be absent (Ge = 0):
where extracellular lactose is given as the discretization of a random variable,
3.3. Reducing Boolean networks
We provide a method to reduce a BN and its corresponding wiring diagram. The idea behind the reduction method is the following: the wiring diagram and Boolean functions should reflect direct regulation and hence nonfunctional edges and variables should be removed; furthermore, vertices can be deleted, without losing important information, by allowing its functionality to be “passed on” to other variables.
We simplify the Boolean functions and wiring diagram: (a) Reduce Boolean expressions using Boolean algebra. This will delete variables that are nonfunctional. (b) Delete edges that do not correspond to Boolean expressions. That is, we delete edges that are nonfunctional. We delete vertices with no self loop, that is, vertices whose Boolean function does not depend on it. Let xi be a vertex such that fxi does not depend on xi. (a) For all vertices xi → y, that is, for all vertices whose Boolean function depends on xi, replace the Boolean function for y, (b) Replace edges y → xi → z by y → z and delete xi (and edges from/to xi).
4. Results
4.1. Boolean network
In this subsection, we present the BN for the lac operon that models gene regulation such as the two main control mechanisms of glucose, namely catabolite repression and inducer exclusion.
The complete BN is given as follows (∧, ∨, and ¬ are the logical AND, OR, and NOT operators, respectively):
FM = C ∧ ¬R∧¬Rm FB = M FR = ¬A ∧ ¬Am FA = L ∧ B FL = P ∧ Le ∧ ¬Ge FP = M FC = ¬Ge
where Le and Ge (lactose and glucose) are considered as parameters in the model. We use F to refer to the model consisting of this BN.
4.1.1. Network topology
The network topology for the model F is shown in Figure 1 and is displayed as a wiring diagram (Le, Lem are shown as a single node, as well as R, Rm, A, Am, and L, Lm); see Section 2.2 for definitions. From the diagram we can identify topological features such as the feedback loops in F. We see that there are at least two positive feedback loops involving M, namely M → P → L → A → R → M and M → B → A → R → M. Note that there are no negative feedback loops.
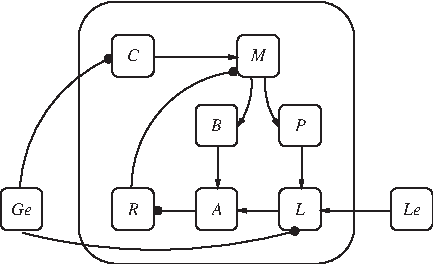
Wiring diagram for the model F. Edges in this diagram represent interactions between variables. Arrows indicate positive interactions, and circles indicate negative interactions.
4.1.2. Dynamics
The dynamics of F can be computed by evaluating the functions on all possible combinations of vectors (M, P, B, C, R, Rm, A, Am, L, Lm) with 0-1 entries (see Section 2.2 for more details). We say that the operon is OFF when the value of the triple (M, P, B) is (0, 0, 0) and ON when (M, P, B) = (1, 1, 1). For simplicity, we will focus on the steady states.
When glucose is high (Ge = 1) we have the unique steady state ( For low/0 extracellular lactose (that is, Le = Lem = 0), there is only one steady state, ( For medium/1 extracellular lactose (that is, Le = 0, Lem = 1), there are two steady states, ( For high/2 extracellular lactose (that is, Le = Lem = 1), there is only one steady state, (
In summary, the model predicts that the lac operon is OFF when extracellular glucose is available. When glucose is not available, the model predicts that the operon is OFF, bistable, or ON when extracellular lactose is low, medium, or high, respectively. This is consistent with the reports of bistability, see for example (Santillán, 2008; Ozbudak et al., 2004). The comparison of our Boolean model with ODE models can be illustrated with the bifurcation diagram shown in Figure 2. This figure shows that both models have the same qualitative behavior.
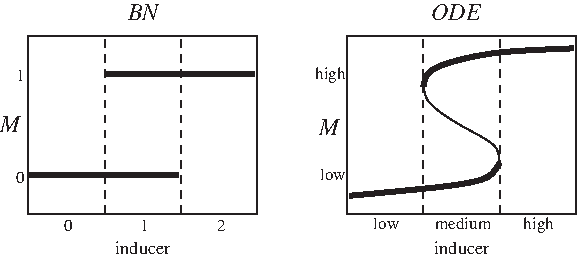
Bifurcation diagram of our Boolean network and a typical bifurcation diagram showing bistability of an ODE (two saddle-node bifurcation).
4.1.3. Bistability
The results of our experiments are in Figure 3 (left). We can see the same qualitative behavior as in the experiments performed in Ozbudak et al. (2004), shown in Figure 3 (right). We can see that as we decrease the concentration of the inducer (top panel) the population of cells start turning their operon OFF; also, as we increase the concentration of the inducer (bottom panel) the population of cells start turning their operon ON.

Results of the bistability experiments performed in the Boolean model F (
4.2. Reduced model
An important question is whether the dynamical properties of the model is a direct consequence of its topology and the type of interactions in the model. If network topology and interaction type are sufficient for maintaining certain dynamical properties, we expect a reduced model to have dynamics equivalent to the original model. This would suggest that the qualitative dynamical properties of the lac operon are dependent on intrinsic characteristics such as topological features and interaction type and not on extrinsic characteristics such as the specific proteins, sugars involved or the size of the network. This in turn implies that in order to obtain an accurate qualitative description of the lac operon we need to focus on using the correct network topology and interaction type. The importance of network topology and interaction type has been proposed by Albert and Othmer (2003), among others, and studied by Aguda and Goryachev (2007). Alternatively, if the dynamics of the model is highly dependent on the size, the model would not be useful to study the dynamics of the biological system: changing the size of the network could significantly alter the predictions.
4.2.1. Reducing the Boolean network
We provide a reduced network that retains topological features, such as feedback loops, present in the original model. Since many existing models with bistable behavior include glucose and lactose, we will construct a reduced model using M, L, Le, Ge only. We denote the reduced model by f.
Boolean function for M: To have mRNA being transcribed we need the activator protein (CAP) which will be present if glucose is absent; we also need the repressor to be absent which will happen if there is lactose in the cell. That is, the future Boolean value of M is given by the ABSENCE of Ge AND {the PRESENCE of Lm OR L}. Hence, the Boolean function for M is fM = ¬Ge ∧ (L ∨ Lm).
Boolean function for L: To have lactose in high concentrations we need both permease (which means we need mRNA) and extracellular lactose to be present, but also glucose to be absent. That is, the future Boolean value of L is given by the PRESENCE of P AND Le AND the ABSENCE of Ge. Hence, the Boolean function for L is fL = M ∧ Le ∧ ¬Ge.
Boolean function for Lm: To have lactose in medium concentrations we need extracellular lactose to be present in at least a medium concentration together with permease (which means we need mRNA), or extracellular lactose to be present in concentrations high enough so that it will enter the cell; we also need glucose to be absent. That is, the future Boolean value of Lem is given by the {{PRESENCE of Lem AND M} OR the PRESENCE of Le} AND the ABSENCE of Ge. Hence, the Boolean function for Lm is
The final reduced model is presented in the next section.
4.2.2. Reduced model
We reduced the Boolean model F by deleting P, B, C, R, Al, Ll, A (for details, see Supplementary Material online at www.liebertonline.com/cmb) to show that a core subnetwork exists which exhibits bistability. The reduced model, denoted by f, contains the variables M, L, Le and Ge. The model f is given by
where Le and Ge are treated as parameters.
4.2.3. Network topology
The wiring diagram for the model f is shown in Figure 4 (Le, Lem are shown as a single node, as well as L, Lm). From the reduced model, we can clearly see the roles of the parameters on the core subnetwork whose size is 30% of that of F. We can see that the paths from Ge and Le to M are still present in the model; also, the signs of these paths have not changed. Ge inhibits M and L (catabolite repression and inducer exclusion, resp.) whereas Le activates L. Furthermore, we still have a positive feedback loop involving M.
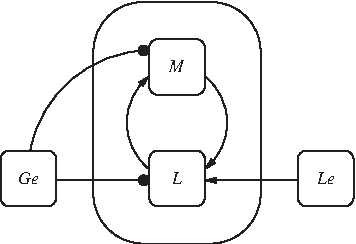
Wiring diagram of f.
Here we have identified the core of the network to be M and L. From the reduced model, we can clearly see the roles of the parameters on the core subnetwork whose size is about 22% of that of F.
4.2.4. Dynamics
The dynamics of f can be computed by evaluating the functions on all possible combinations of vectors (M, L, Lm) with 0–1 entries. We say that the operon is OFF when the value of
When glucose is high (Ge = 1), we have the unique steady state ( For low/0 extracellular lactose, there is a single steady state, ( For medium/1 extracellular lactose, there are two steady states, ( For high/2 extracellular lactose, there is a single steady state, (
In summary, the model predicts that the lac operon is OFF when extracellular glucose is available. When glucose is not available, the model predicts that the operon is OFF, bistable or ON when extracellular lactose is low, medium, or high, respectively. Hence, the reduced network has the same dynamics as the full network. We observe that the reduced model has only one positive feedback loop, whereas the model F has several more. Since the reduced model still exhibits bistability, this suggests that it does not depend on the number of positive feedback loops but simply on the existence of such a loop.
4.2.5. Bistability
The region of bistabilty for the model f is similar to that shown in the left panel of Fig. 3 (data not shown for f). This suggests that bistability does not depend on the number of positive feedback loops but simply on the existence of such a loop and stochasticity. Furthermore, because the feedback loop involves only M and L, this suggests that bistability is maintained by stochasticity and the interaction between the operon (represented by M) and lactose.
It is important to notice that the state space of the reduced model has 23 states which is about 0.8% of the size of the state space of F, that is, 210 states. We also considered models with different numbers of variables, ranging from four to nine vertices (plus two parameters), and obtained the same results. Hence, we conclude that the dynamical properties of the model are independent of its size.
4.3. Alternative models
Since it has been argued that inducer exclusion is not necessary for bistability (van Hoek and Hogeweg, 2006), we considered alternative models without inducer exclusion and without catabolite repression (Fig. 5) and bistability was also observed.

Wiring diagram of the model without inducer inclusion and without catabolite repression.
FM = C ∧ ¬R∧¬Rm FB = M FR = ¬A ∧ ¬Am FA = L ∧ B FL = P ∧ Le FP = M FC = ¬Ge
FM = C ∧ ¬R∧¬Rm FB = M FR = ¬A ∧ ¬Am FA = L ∧ B FL = P ∧ Le ∧ ¬Ge FP = M FC = 1
Furthermore, their reduced versions (Fig. 6) also showed bistability.
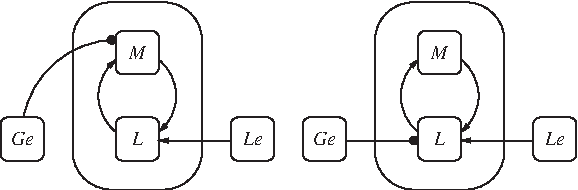
Wiring diagram of the reduced model without inducer inclusion and without catabolite repression.
5. Conclusion
Many authors have studied the problem of inferring dynamical properties of a system from the network structure, including Jarrah et al. (2010) and Sontag et al. (2008). Furthermore, for special classes of BNs and ODEs it has been proven that the network structure contains all the information needed to determine certain dynamical properties (Halász et al., 2007), such as bistability. On the other hand, it has been claimed that network topology and sign of interactions are more important than quantitative functionality of the components of a system (Albert and Othmer, 2003). To test this hypothesis, we applied the ideas of Albert and Othmer (2003) to lactose metabolism.
The lac operon has been studied extensively and is one of the earliest discovered gene systems that undergoes both positive and negative control. While there are numerous continuous models of the lac operon, few discrete models exist (Gianchandani et al., 2006; Setty et al., 2003). Gianchandani et al. (2006) introduced a Boolean model of the lac operon along with a method for identifying all steady states; however, their model does not include the two main glucose control mechanisms. The model in Setty et al. (2003) is a logical (on four states) function for the lac genes written in terms of the regulators CRP and LacI that is capable of accurately predicting induction of the operon based on concentration levels of the regulators. Further, this function has been shown to be robust with respect to point mutations (Mayo et al., 2006). One limitation of this model, however, is that it does not predict bistability, as has been reported and confirmed previously (Novick and Weiner, 1957; Yildirim et al., 2004; Santillán and Mackey, 2004; Ozbudak et al., 2004; Halász et al., 2007; Santillán et al., 2007; Santillán, 2008).
The contributions in this work are twofold. First, we proposed a BN as a discrete model for the lac operon and included the glucose control mechanisms of catabolite repression and inducer exclusion. We showed that our model exhibits the ON/OFF switching dynamics and that bistability is also observed, in accordance with the work of Santillán and coauthors (Yildirim et al., 2004; Ozbudak et al., 2004; Santillán and Mackey, 2004; Santillán et al., 2007; Santillán, 2008). Since it has been argued that inducer exclusion is not necessary for bistability (van Hoek and Hogeweg, 2006), we considered alternative models without inducer exclusion and bistability was also observed.
Second, we presented a reduced model which shows that lac mRNA and lactose form the core of the lac operon, and that this reduced model also exhibits the same dynamics. This result suggests that the presence of positive feedback loops is a better indicator of dynamics than the individual constituents that comprise the feedback loops. Taken together, this work supports the idea that topology is the driver of dynamics (Albert and Othmer, 2003; Mendoza and Xenarios, 2006).
The use of BNs in modeling has many advantages, such as their mathematical formulation typically being more intuitive than that of differential-equations-based models. BNs are particularly useful in the case where one is interested in qualitatively behavior. For example, our Boolean models exhibit bistability when stochasticity is incorporated, and this depends on topological features rather than on the network itself.
A future work may be to extend this model to a multi-state framework, which has the potential to provide a more refined qualitative description of the lac operon. Such a framework may allow for inclusion of other features of the operon, such as multiple promoter and operator regions.
Footnotes
Acknowledgments
We thank Reinhard Laubenbacher for his support of this work.
Disclosure Statement
No competing financial interests exist.
1
We make the first three entries bold as these are the entries of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.