Metabolic flux analysis (MFA), a key technology in bioinformatics, is an effective way of analyzing the entire metabolic system by measuring fluxes. Many existing MFA approaches are based on differential equations, which are complicated to be solved mathematically. So MFA requires some simple approaches to investigate metabolism further. In this article, we applied continuous-time Markov chain to MFA, called MMFA approach, and transformed the MFA problem into a set of quadratic equations by analyzing the transition probability of each carbon atom in the entire metabolic system. Unlike the other methods, MMFA analyzes the metabolic model only through the transition probability. This approach is very generic and it could be applied to any metabolic system if all the reaction mechanisms in the system are known. The results of the MMFA approach were compared with several chemical reaction equilibrium constants from early experiments by taking pentose phosphate pathway as an example.
1. Introduction
The metabolic network can be analyzed from various angles, and metabolic flux analysis (MFA) is the most popular one (Bailey, 1991; Stephanopoulos et al., 1998). MFA is in fact a computational model of the intracellular metabolic network (Fischer et al., 2004). The idea of MFA is to build a model containing flows and metabolites. Considering that the inflow is equal to the outflow when the metabolic process reaches its steady state, differential equations can be established to analyze the concentration of each metabolite at the steady state (Selivanov et al., 2004). In addition, the minimum error between analytical data obtained from the model and actual measurement data at the steady state can be considered as an indicator for assessment. MFA can be simulated by the Monte Carlo method (Kadirkamanathan et al., 2006; Schellenberger et al., 2012), Bayesian estimation simulation (Jayawardhana et al., 2008), and Markov process (Zhang, 2013), or a hybrid method, such as Markov chain Monte Carlo (Kadirkamanathan et al., 2006). Analytical data can be obtained through computer programming (Chen and Ji, 2011) and then analyzed using Lagrange multiplier methods (Wiechert et al., 1997; Xu et al., 2013) and finally processed with MFA.
In order to figure out the reaction mechanism in the metabolic process, generally, we use the method of 13C isotope labeling. From the perspective of experimental analysis, MFA is carried out according to the relative concentration of metabolites containing 13C, which can be obtained through hydrogen-1 nuclear magnetic resonance (1H NMR) spectroscopy experiment. Theoretically, a model can be built to simulate the metabolic pathway of metabolites containing 13C (Benevolensky et al., 1994; Schellenberger et al., 2012). The 13C marking method can be expressed by Monte Carlo random simulation (Schellenberger et al., 2012). A binary representation was used to express the reaction mechanism of metabolites containing 13C by Selivanov et al. (2004). Then, experimental and theoretical analysis can be compared to calculate the relative error that serves as the assessment indicator of the model (Gombert et al., 2001).
In this article, chemical dynamics, bioinformatics, and mathematical statistics are combined to carry out MFA. On the basis of chemical dynamics, we use the simple mathematical relations between the instantaneous rate (v) of the reaction and the concentration of the reaction products (A and B) as follows:
\documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
\begin{align*}\nu = k [ A ] ^{ \alpha} [ B ] ^{ \beta} \tag{1}\end{align*}
\end{document}
where k is a constant indicating the chemical reaction rate that can be measured by NMR experiments (Milstein and Bremermann, 1979; Greenfield et al., 1988) or calculated by establishing correspondent differential equations. Furthermore, the metabolic process can be seen as the transition process of carbon atoms that conforms to the Markov process, that is, the no memory property (Maruyama, 1955). Therefore, we can establish a Markov model to carry out MFA (MMFA), and thus obtain the relative concentrations of various metabolites when the metabolic process reaches the steady state.
For the purpose of verification, we select the pentose phosphate pathway to calculate the relative concentrations of individual metabolites at the steady state. From the result, we calculate the equilibrium constants of some reversible reaction in the metabolic process, and compare them with the measured data.
2. Methods
Consider a simple metabolic system that contains m metabolites denoted as \documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
$$A_i \ ( i = 1 , 2 , \, \ldots , m )$$
\end{document}, and Ni represents the number of carbon atoms in the carbon backbone of Ai. First, suppose that an initial metabolite A1 is selected. In fact, we can select any metabolite \documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
$$A_i \ ( i = 1 , 2 , \, \ldots , m )$$
\end{document}. Besides, suppose that A1 has n carbon atoms, that is, N1=n (n ≥ 1). Our problem is to derive the relative concentration for each metabolite \documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
$$A_i \ ( i = 1 , 2 , \, \ldots , m )$$
\end{document} compared with the initial concentration of A1 when the entire metabolic system reaches its steady state. We assume that the initial concentration of A1 is defined as 1, while the initial concentration of \documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
$$A_i \ ( i = 2 , \, \ldots , m )$$
\end{document} is defined as 0. The metabolic process can be regarded as a transfer process for carbon atoms existed in A1, so our problem can be transformed to study the distribution of each metabolite when every carbon atom in A1 reaches the steady state.
In order to solve the above problem, we first assign a serial number for each of the carbon atoms in A1 referred to as \documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
$$1 , 2 , \, \ldots , n. \ X^h ( t )$$
\end{document} represents the state of the hth carbon atom at time t, where the state space is \documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
$$M = \{ 1 , \, \ldots , m \} $$
\end{document}. For example, \documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
$$X^h ( t ) = a \ ( a \in M )$$
\end{document} indicates that the hth carbon atom of A1 is in Aa at time t. \documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
$$P_{ab}^h ( t , t^{ \prime} )$$
\end{document} stands for the transition probability for the hth carbon atom from Aa to Ab after a time interval t′, that is,
\documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
\begin{align*}P_{ab}^h ( t , t^{ \prime} ) = Pr \,\left( X^h ( t + t^{ \prime} ) = b \mid X^h ( t ) = a \right) \tag{2}\end{align*}
\end{document}
The transition matrix for the hth carbon atom of Ai is expressed as follows:
\documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
\begin{align*}P^h ( t , t^{ \prime} ) = ( P_{ab}^h ( t , t^{ \prime} ) )_{m \times m}\end{align*}
\end{document}
Thus, we can obtain the density matrix Q by
\documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
\begin{align*}Q^h ( t ) = \frac { dP^h ( t , t^ { \prime } ) } { dt^ { \prime } } = \left( \frac { dP_ { ab } ^h ( t , t^ { \prime } ) } { dt^ { \prime } } \right) _ { m \times m } : = \left( q_ { ab } ^h ( t ) \right) _ { m \times m } \tag { 3 } \end{align*}
\end{document}
If \documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
$$a \neq b$$
\end{document}, for the continuous-time Markov chain, we have
\documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
\begin{align*} q_ { ab } ^h ( t ) = \frac { dP_ { ab } ^h ( t , t^ { \prime } ) } { dt^ { \prime } } \\ = \lim_ { t^ { \prime } \rightarrow 0 } \frac { P_ { ab } ^h ( t , t^ { \prime } ) } { t^ { \prime } } \end{align*}
\end{document}\documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
\begin{align*}= \frac { \nu_ { a \rightarrow b } ^h ( t ) } { [ A_a ( t ) ] } \tag { 4 } \end{align*}
\end{document}
where [Aa (t)] represents the relative concentration of Aa at time t.
If a=b, then
\documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
\begin{align*}q_{ab}^h ( t ) = - \sum \limits_{b = 1 , b \neq a}^m q_{ab}^h ( t ) \tag{5}\end{align*}
\end{document}
We assume that there are gh reactions in the transfer process from Aa to Ab of the hth carbon atom. When combined with Equation 1,
\documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
\begin{align*}\nu_{a \rightarrow b}^h ( t ) = \sum \limits_{w = 1}^{gh} k_w [ A ] ^{ \alpha} [ B ] ^{ \beta} \tag{6}\end{align*}
\end{document}
where kw represents the reaction rate constant of the wth \documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
$$( w = 1 , 2 , \, \ldots , g_h )$$
\end{document} reaction in Equation 1.
For other reactions without the transfer process of the hth carbon atom, then
\documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
\begin{align*}\nu_{a \rightarrow b}^h ( t ) = 0 \tag{7}\end{align*}
\end{document}
All reactions are assumed to be noncomplex here.
Assuming that \documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
$$\pi_j^h$$
\end{document} is the probability that the hth carbon atom existed in Ai transfers to Aj when the system reaches the steady state. So the steady-state equation is
\documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
\begin{align*}\left( \pi_1^h , \pi_2^h , \ldots , \pi_m^h \right) Q^h ( \infty ) = 0 , \ { \rm and} \ \sum \limits_{j = 1}^m \pi_j^h = 1 \ h = 1 , 2 , \, \ldots , n \tag{8}\end{align*}
\end{document}
The entire metabolic process in fact is the transition of carbon atoms existed in A1, so that at the steady state, the total number of carbon atoms that have transferred to Aj is
\documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
\begin{align*}[ A_1 ( 0 ) ] V_0 ( \pi_j^1 + \pi_j^2 + \cdots + \pi_j^n ) j = 1 , 2 , \, \ldots , m \tag{9}\end{align*}
\end{document}
where V0 stands for the entire volume (assumed to be unchanged during the metabolic process). Since each metabolite Aj contains Nj carbon atoms, the total number of metabolites at the steady state is
\documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
\begin{align*} \frac { [ A_1 ( 0 ) ] V_0 ( \pi_j^1 + \pi_j^2 + \cdots + \pi_j^n ) } { N_j } \tag { 10 } \end{align*}
\end{document}
Therefore, the relative concentration of Aj at the steady state can be derived as
\documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
\begin{align*}[ A_j ( \infty ) ] = \frac { [ A_1 ( 0 ) ] V_0 ( \pi_j^1 + \pi_j^2 + \cdots + \pi_j^n ) } { V_0N_j [ A_1 ( 0 ) ] } = \frac { ( \pi_j^1 + \pi_j^2 + \cdots + \pi_j^n ) } { N_j } \tag { 11 } \end{align*}
\end{document}
By combining Equations 4–8 and 11, a set of quadratic equations can be obtained. Then we can calculate \documentclass{aastex}\usepackage{amsbsy}\usepackage{amsfonts}\usepackage{amssymb}\usepackage{bm}\usepackage{mathrsfs}\usepackage{pifont}\usepackage{stmaryrd}\usepackage{textcomp}\usepackage{portland, xspace}\usepackage{amsmath, amsxtra}\pagestyle{empty}\DeclareMathSizes{10}{9}{7}{6}\begin{document}
$$\pi_j^h$$
\end{document}, which is again plugged into Equation 11 so that the relative concentration for each metabolite compared with the initial concentration of Ai at the steady state can be obtained (see Supplementary Material at www.liebertonline.com).
3. Application
In this section, we apply the proposed MMFA approach to flux distribution analysis in the pentose phosphate pathway.
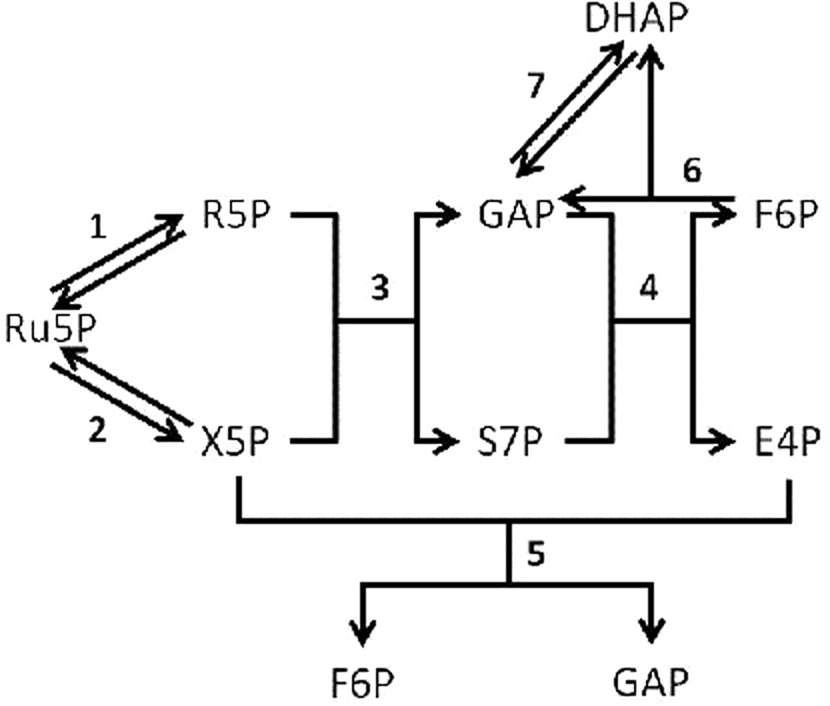
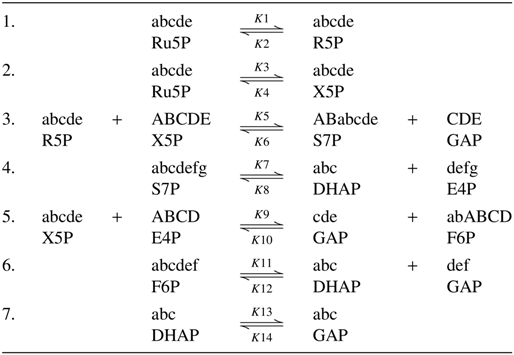
First, we can draw a schematic diagram of the pentose phosphate pathway (Fig. 1). From Figure 1, we can easily discover that the entire system contains eight metabolites and seven reactions. The eight metabolites are represented as A1(Ru5P), A2(R5P), A3(X5P), A4(S7P), A5(GAP), A6(F6p), A7(E4P), and A8(DHAP). The mechanisms of these seven reactions are listed in Table 1.
The process of the pentose phosphate pathway simulated in the model. Reaction 1 is Ru5P-R5P; reaction 2 is Ru5P-X5P; reaction 3 is R5P+X5P-S7P+GAP; reaction 4 is S7P-DHAP+E4P; reaction 5 is X5P+E4P-GAP+F6P; reaction 6 is F6P-DHAP+GAP; reaction 7 is DHAP-GAP. Details of the reaction are shown in reaction mechanism. DHAP, dihydroacetone phosphate; E4P, erythrose 4-phosphate; F6P, fructose 6-phosphate; GAP, glyceraldehyde 3-phospate; R5P, ribose 5-phosphate; Ru5P, ribulose 5-phosphate; S7P, sedoheptulose 7-phosphate; X5P, xylulose 5-phosphate.
The Mechanisms of The Seven Reactions
In Table 1, kw is a constant indicating the chemical reaction rate, shown in Table 2 (Milstein and Bremermann, 1979).
Exact Value of kw
kw
Exact value
k1
11.52
k2
26.72
k3
9.67
k4
6.91
k5
5.26
k6
4.46
k7
0.0017
k8
22.85
k9
267.78
k10
21.27
k11
0.000280
k12
1.83
k13
0.066
k14
1.38
In this pathway, by the analysis of the carbon atom's reaction mechanism in the entire process, we find that the first carbon atom (i.e., “a” in Table 1) in A1 can transfer back to any position (i.e., a–e) in A1. So the transfer process of each carbon atom is equivalent; this means Q1=Q2=Q3=Q4=Q5. Then we can only calculate the value of Q1. After simulating the transfer process of the first carbon atom, we find that it is hard to obtain the accurate probability of which product contains the first carbon atom if there are two or more products in the reaction. In fact, in the entire pathway, the rate of each reaction is different. So in this article, we distribute the probability by the ratio of products' factors in the reaction, that is, 1:1 in Table 1 obviously.
To test the effect of the MMFA approach, we evaluate the relative concentration through the reaction equilibrium constant (K) in Section 4.
4. Results
Using MATLAB, we input an initial concentration of A1 and we got a group of probabilities when the system reached the steady state. Also we obtained the related concentration of each metabolism at the steady state. To test the approach, we selected the reaction equilibrium constant K to be the assessment indicator because each reaction in the pentose phosphate pathway is a reversible one.
The pentose phosphate pathway has been studied by many researchers (Kleijn et al., 2006; Luo et al., 2007; Zhao et al., 2008). Although it is simple and small, the most reactants and products in the pentose phosphate pathway are related with other metabolic systems. In this article, we selected the second, sixth, and seventh reactions to test the approach. Each K of these three reactions is listed in Table 3.
Each Chemical Reaction Equilibrium Constant K of the Second, Sixth, and Seventh Reactions
In MATLAB, we verified the initial concentration (defined as u) from 10 to 50 mmol/L and the distance value is 5 mmol/L, and the results are shown in Tables 4–6.
The Actual Error and Relative Error Between the K1's Estimation Value and Exact Value When u Changes from 10 to 50 mmol/L with the Distance Value of 5 mmol/L
Value of u (mol/L)
Estimation value
Exact value
Actual error
Relative error (%)
0.01
1.399351
1.4
0.000649
0.0464
0.015
1.399334
1.4
0.000666
0.0476
0.02
1.39932
1.4
0.00068
0.0486
0.025
1.399309
1.4
0.000691
0.0494
0.03
1.3993
1.4
0.0007
0.0500
0.035
1.399309
1.4
0.000691
0.0494
0.04
1.399284
1.4
0.000716
0.0511
0.045
1.399277
1.4
0.000723
0.0516
0.05
1.399271
1.4
0.000729
0.0521
Average
1.3993061
1.4
0.0006939
0.0496
The Actual Error and Relative Error Between the K2's Estimation Value and Exact Value When u Changes from 10 to 50 mmol/L with the Distance Value of 5 mmol/L
Value of u (mol/L)
Estimation value
Exact value
Actual error
Relative error (%)
0.01
0.048174
0.047
0.001174
2.50
0.015
0.048364
0.047
0.001364
2.90
0.02
0.048555
0.047
0.001555
3.31
0.025
0.048746
0.047
0.001746
3.71
0.03
0.048937
0.047
0.001937
4.12
0.035
0.048746
0.047
0.001746
3.71
0.04
0.049321
0.047
0.002321
4.94
0.045
0.049515
0.047
0.002515
5.35
0.05
0.049709
0.047
0.002709
5.76
Average
0.048896
0.047
0.001896
4.03
The Actual Error and Relative Error Between the K3's Estimation Value and Exact Value When u Changes from 10 to 50 mmol/L with the Distance Value of 5 mmol/L
Value of u (mol/L)
Estimation value
Exact value
Actual error
Relative error (%)
0.01
0.097444
0.086
0.011444
13.31
0.015
0.086965
0.086
0.000965
1.12
0.02
0.081333
0.086
0.004667
5.43
0.025
0.077802
0.086
0.008198
9.53
0.03
0.075388
0.086
0.010612
12.34
0.035
0.077802
0.086
0.008198
9.53
0.04
0.072343
0.086
0.013657
15.88
0.045
0.071343
0.086
0.014657
17.04
0.05
0.070562
0.086
0.015438
17.95
Average
0.078998
0.086
0.009761
11.35
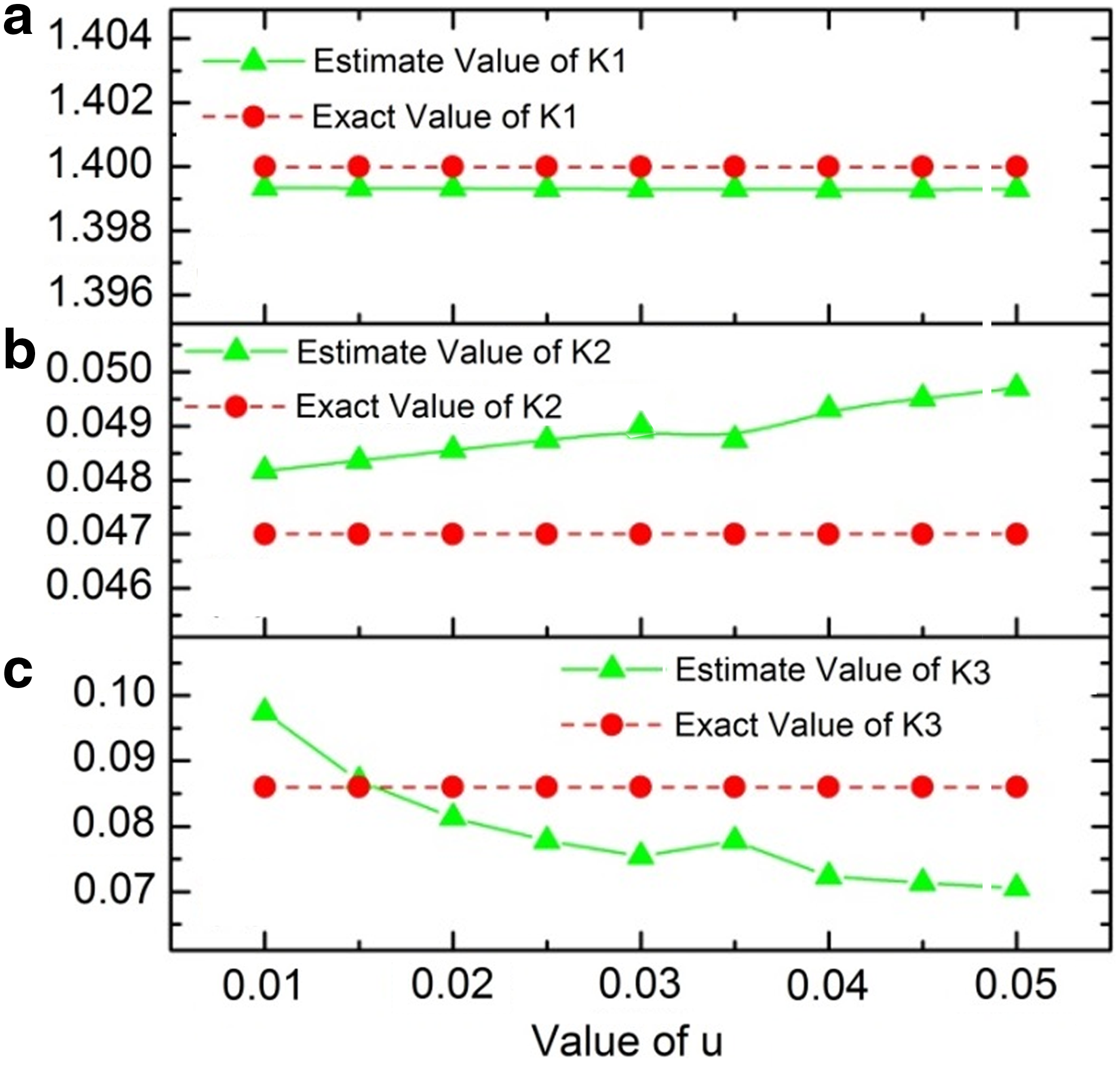
The results are also shown in Figure 2, where solid lines represent the estimation values of K1, K2, and K3, while dotted lines are the exact values. From Figure 2a, the relative error of K1 between the estimation value and the exact value is extremely small, so the estimation value is quite accurate. Also, the estimation value of K2 is very close to its exact value, and its relative error is less than 6%, as seen in Table 5 and Figure 2b. The relative error of K3 is a little more than K1 and K2; however, the estimation value is very close to its exact value. The relative error of K3 is a little bit large because the corresponding metabolite F6P can also be produced by G6P in the entire metabolic system, but G6P is not included in the pentose phosphate pathway. In all, we can conclude that the results of this approach are quite reliable and accurate.
(a) The estimation value of K1 when w changes from 10 to 50 mmol/L with the distance value of 5 mmol/L. (b) The estimation value of K2 when u changes from 10 to 50 mmol/L with the distance value of 5 mmol/L. (c) The estimation value of K3 when u changes from 10 to 50 mmol/L with the distance value of 5 mmol/L.
5. Conclusion
MFA is a powerful technique for determining intracellular metabolic fluxes in living cells. MFA has been found its widespread use in metabolic engineering, systems biology, biomedical research, and biotechnology.
In this article, a brand new approach, which combines MFA and continuous-time Markov chain, has been put forward to analyze metabolic flux in the metabolic system. On the basis of the study of the pentose phosphate pathway discussed in the Application section, this approach calculated the steady-state concentration by the distribution of each carbon atom. We obtained the reaction equilibrium constant for analysis and comparison, and found that this approach is feasible and the results are accurate. The approach proposed in this article is generic,mjm, which can be applied to any metabolic systems.
Footnotes
Acknowledgments
The work described in this article was supported by a grant from The Hong Kong Polytechnic University (Project G-UA71).
Author Disclosure Statement
The authors declare that no competing financial interests exist.
References
1.
BaileyJ.E.1991. Toward a science of metabolic engineering. Science, 252, 1668–1675.
2.
BenevolenskyS.S.V., CliftonD., and FraenkelO.D.G.1994. The effect of increased phosphoglucose isomerase on glucos metabolism in Saccharomyces cerevisiae. J. Biol. Chem., 269, 4878–4882.
3.
CasazzaJ.P., and VeechR.L.1986. The interdependence of glycolytic and pentose cycle intermediates in ad libitum fed rats. J. Biol. Chem., 261, 690–698.
4.
ChenC.S.H., and JiP.2011. Decomposing flux distributions into elementary flux modes in genome-scale metabolic networks. Bioinformatics, 27, 2256–2262.
5.
ConnettR.J.1985. In vivo glycolytic equilibria in dog gracilis muscle. J. Biol. Chem., 260, 3314–3320.
6.
FischerE., ZamboniN., and SauerU.2004. High-throughput metabolic flux analysis based on gas chromatography-mass spectrometry derived 13C constraints. Anal. Biochem., 325, 308–316.
7.
GombertA.K., dos SantosM.M., ChristensenB., and NielsenJ.2001. Network identification and flux quantification in the central metabolism of Saccharomyces cerevisiae under different conditions of glucose repression. J. Bacteriol. 183, 1441–1451.
8.
GreenfieldN.J., McKenzieM.A., AdebodunF., et al.1988. Metabolism of D-glucose in a wall-less mutant of Neurospora crassa examined by 13C and 31P nuclear magnetic resonances: effects of insulin. Biochemistry, 27, 8526–8533.
9.
JayawardhanaB., KellD.B., and RattrayM.2008. Bayesian inference of the sites of perturbations in metabolic pathways via Markov chain Monte Carlo. Bioinformatics, 24, 1191–1197.
10.
KadirkamanathanV., YangJ., BillingsS.A., and WrightP.C.2006. Markov Chain Monte Carlo Algorithm based metabolic flux distribution analysis on Corynebacterium glutamicum. Bioinformatics, 22, 2681–2687.
11.
KleijnR.J., van WindenW.A., RasC., et al.2006. 13C-labeled gluconate tracing as a direct and accurate method for determining the pentose phosphate pathway split ratio in Penicillium chrysogenum. Appl. Environ. Microbiol., 72, 4743–4754.
12.
LuoB., GroenkeK., TakorsR., et al.2007. Simultaneous determination of multiple intracellular metabolites in glycolysis, pentose phosphate pathway and tricarboxylic acid cycle by liquid chromatography–mass spectrometry. J. Chromatogr. A, 1147, 153–164.
MilsteinJ., and BremermannH.J.1979. Parameter identification of the Calvin photosynthesis cycle. J. Math. Biology, 7, 99–116.
15.
SchellenbergerJ.D., ZielinskiC., ChoiW., et al.2012. Predicting outcomes of steady-state 13C isotope tracing experiments using Monte Carlo sampling. BMC Syst. Biol., 6, 9.
16.
SelivanovV.A., PuigjanerJ., SilleroA., et al.2004. An optimized algorithm for flux estimation from isotopomer distribution in glucose metabolites. Bioinformatics, 20, 3387–3397.
17.
StephanopoulosG.N., AristidouA.A., and NielsenJ.1998. Metabolic Engineering. Academic Press, San Diego, CA.
18.
VeechR.L., RaijmanL., DalzielK., and KrebsH.A.1969. Disequilibrium in the triose phosphate isomerase system in rat liver. Biochem. J. 115, 837–842.
19.
WiechertW., SiefkeC., de GraafA.A., and MarxA.1997. Bidirectional reaction steps in metabolic networks: II. Flux estimation and statistical analysis. Biotechnol. Bioeng., 55, 118–135.
20.
XuZ.X., SunX., and SunJ.B.2013. Construction and analysis of the model of energy metabolism in E. coli. PLoS ONE, 8, e55137.
21.
ZhangY.2013. A dynamic Bayesian Markov model for phasing and characterizing haplotypes in next-generation sequencing. Bioinformatics, 10, 1–8.
22.
ZhaoZ., KuijvenhovenK., RasC., et al.2008. Isotopic non-stationary 13C gluconate tracer method for accurate determination of the pentose phosphate pathway split-ratio in Penicillium chrysogenum. Metab. Eng., 10, 178–186.
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.