
Abstract
The expression of genes is inevitably subject to intracellular noise. Noise, for some regulatory networks, is constructive but detrimental to many others. The intensity of the noise is a determinant factor and the method of tuning it is of great value. In this study, we illustrated that the transcriptional delay in an incoherent feedforward loop (FFL) grants the target protein modulation the intensity of noise. Remarkably, for a wide range, the coefficient of variation (COV) of the target protein appeared to be about linear to the time span of the transcriptional delay. Without a noise-buffering method, the COV of the target protein is 0.455. While applying incoherent FFL, the COV reduced to 0.236. Then, it changed from 0.236 to 0.630 as the transcriptional delay raised from 0 to 1000 seconds. If we further increased the delay out of the linear range, the COV finally reached 0.779. In addition, we incorporated the distribution of the transcriptional delay in the delay stochastic simulation algorithm. This distribution is based on the experimental observation in the literature. The outcome suggested that the distributed delay slightly improved the ability of tuning noise. In conclusion, we demonstrated a noise-tuning method that altered only the intensity of noise without changing the deterministic steady-state behaviors. It is ready to be applied to various systems in the field of synthetic biology.
1. Background
Stochasticity is ubiquitous in biological systems. The protein level notably deviates from cell to cell due to the intracellular noise. The noise propagation in gene regulatory networks has important consequences for cellular behaviors. Even in a homogeneously environmental condition, the gene expression varies across genetically identical cells. This is because the underlying events of reactions involve the small number of molecules. The intracellular noise is implicated in diverse processes such as the decision of cell fates (Tian and Burrage, 2006; Shu et al., 2016; Xu et al., 2016), the induction of the bimodality (To and Maheshri, 2010), and the detection of signals (Paulsson et al., 2000; Shu et al., 2013). Without a doubt, the intensity of the noise is the determinant factor for all these processes. Thus, cells developed a means to cope with stochastic fluctuations.
The noise buffering methods, such as negative feedback control (Dublanche et al., 2006) and the incoherent feed-forward controls (Osella et al., 2011), were discovered in recent decades. It has been reported that the feedforward loops (FFLs) are one of the critical motifs in gene regulatory networks (Chepyala et al., 2016). Among these, the most well-known one is the microRNA-mediated FFL. An upstream transcription factor (TF) regulates the target gene as well as the microRNA (abbreviated as miRNA). There are coherent and incoherent FFLs (Estrada and Guantes, 2013). For coherent FFLs, both TF and miRNA have the same effect on the protein production of the target gene. For incoherent FFLs, the TF and miRNA have opposite effects on the generation of the target protein. One of the most popular motifs of the incoherent FFLs is the co-expression of the miRNA and the target gene (Fig. 1) by TF, but the miRNA represses the generation of the target protein. Such an incoherent FFL remarkably dampens stochastic fluctuations at the protein level (Strovas et al., 2014).

The incoherent FFL with a transcriptional delay of miRNA. The TF activates the generation of miRNA and the target RNA. The miRNA represses the translation of target RNA. In this study, we discussed the influence of the transcriptional delay of miRNA in the incoherent FFL. FFL, feedforward loop; TF, transcription factor.
In general, the noise-buffering method not only attenuated the noise but also reduced the level of the target protein. To the best of my knowledge, there are three models claiming the possibility of tuning the intensity of the noise without changing the deterministic steady state (Lu et al., 2008; Ribeiro et al., 2010; Baker et al., 2012). Baker applied multiple sRNA instead of single sRNA to the system. The multiple sRNA cause a different intensity of the protein noise from that of the single sRNA. Diverse combinations of the sRNA offer the possibility of tuning the protein noise without altering its mean. Ribeiro proposed a noise-tuning method by simultaneously adjusting the delay of the promoter open complex and the initiation rate of the transcription. The protein noise was then tuned independently from the protein level. Lu used the fact that the translation caused much more protein noise than transcription. The increment of the translation rate constant leads to the incremental noise intensity of protein. By adjusting the transcription rate accordingly, it is possible to maintain the deterministic protein level regardless of the change of the protein noise. Nevertheless, all of these methods require trial and error. It is somehow impracticable in wet laboratories. It is also possible to tune the noise by altering the DNA sequence (Murphy et al., 2010). Murphy applied mutation to the TATA box to change the expression noise. Nevertheless, this method inevitably leads to the reduction of expression level. The main goal of this study is to propose a method to tune the noise intensity at the protein level.
2. Methods
Our model (Fig. 1) was adopted from the framework of the incoherent FFL discussed in the literature (Osella et al., 2011). The gene circuit is composed of three genes. They are the gene encoding TF, the gene producing miRNA, and the target gene. The TF and miRNA have opposite effects on the generation of the target protein. We accounted for the generation and degradation of each variable except for the DNA that is one per cell all the time. The activation of transcription by the TF [Eq. (1) for the target gene and Eq. (2) for the gene of miRNA] and the repression of translation by miRNA [Eq. (3)] were described by the following:
The values of the parameters were acquired from the same article (Osella et al., 2011). In addition, we further accounted for cell division. The distribution of division time is assumed to be a Gaussian distribution, where the mean and standard deviation were obtained according to the experimental observation in the literature (Song et al., 2015). In brief, the standard deviation is 10% of the mean. We then applied binomial distribution to cell partitioning (Rosenfeld et al., 2005). Furthermore, we also take the transcriptional delay of miRNA into account.
For stochastic simulations, we applied the stochastic simulation algorithm (SSA) (Gillespie, 1977) to the reactions in Table 1. The nomenclature is in Table 2 and the values of parameters in Table 3. We applied delay SSA (Bratsun et al., 2005) to the transcriptional delay of miRNA. While applying the distributed delay, we generated a Gaussian random number and assigned it to the time delay. The distributions of target protein were sampled after reaching stationary distributions, which is suggested by time-independent first and second moments. We took the results at 50,000 seconds and the distributions shown in figures were composed of 10,000 trajectories. We obtained the variance also at 50,000 seconds from at least 1000 trajectories. The same initial condition was implemented in all cases. The initial condition is 0 for each intracellular variable except that of DNA is 1. Note that the initial conditions have no influence on the outcome because the outcome was obtained from the stationary distribution. All data analysis, as well as the simulation, were conducted in Matlab.
The Reactions of the System
Nomenclature of the Variable
The Values of Parameters
These values of the parameters are slightly different from the literature (Osella et al., 2011). The values used in the literature are in the bracket. We adjusted the value to have better performance or to reduce computational burden.
3. Results
3.1. miRNA buffering the noise of the target protein
The stochastic fluctuations of the TF contribute noise to the target RNA and drive the target protein away from its mean value. Namely, the target protein inherited the noise of the TF. Such noise can be buffered by the incoherent FFL. Figure 1 showed the reaction networks discussed in this study. Figure 2A showed the trajectories of the miRNA and the target RNA with 100 seconds as the time span of sampling. Clearly, the miRNA and the target RNA are highly correlated. The origin of buffering noise by miRNA in incoherent FFLs is following. The miRNA and the target RNA tend to fluctuate in a similar pattern because they both are regulated by the TF. Under the circumstance of no transcriptional delay, miRNAs reduced the noise of the target protein. The red dash in Figure 2B is the distribution of the target protein with its noise buffered by miRNA, but the blue circle is that without miRNA. Note that we adjusted the transcription rate of the target protein to keep the same protein level. The number above the distribution is the coefficient of variation (COV). The distribution of the target protein in the case of incoherent FFL is much sharper because the noise is buffered.

The influence of the transcriptional delay on the distribution of the target protein.
3.2. Manipulating noise through transcriptional delay
The green curve of Figure 2B illustrated the distribution of target protein with the transcriptional delay of miRNA as 1000 seconds. The COV of the target protein became even higher than that in the system without an FFL. While accounting for the transcriptional delay of miRNA, the fluctuations of miRNA and the target RNA were in a similar pattern but with a time span difference. Figure 2C demonstrated the trajectories of miRNA and target RNA with the transcriptional delay of miRNA as 1000 seconds. When the time delay is large enough, it makes miRNA fail to buffer the noise of the target protein. The correlation between miRNA and the target RNA drops from 0.8 to nearly 0 (0.01) while applying the delay of miRNA from 0 to 1000 seconds. Consequently, the noise of the target protein raised up. This conclusion is valid for various TF numbers as shown in Figure 2D.
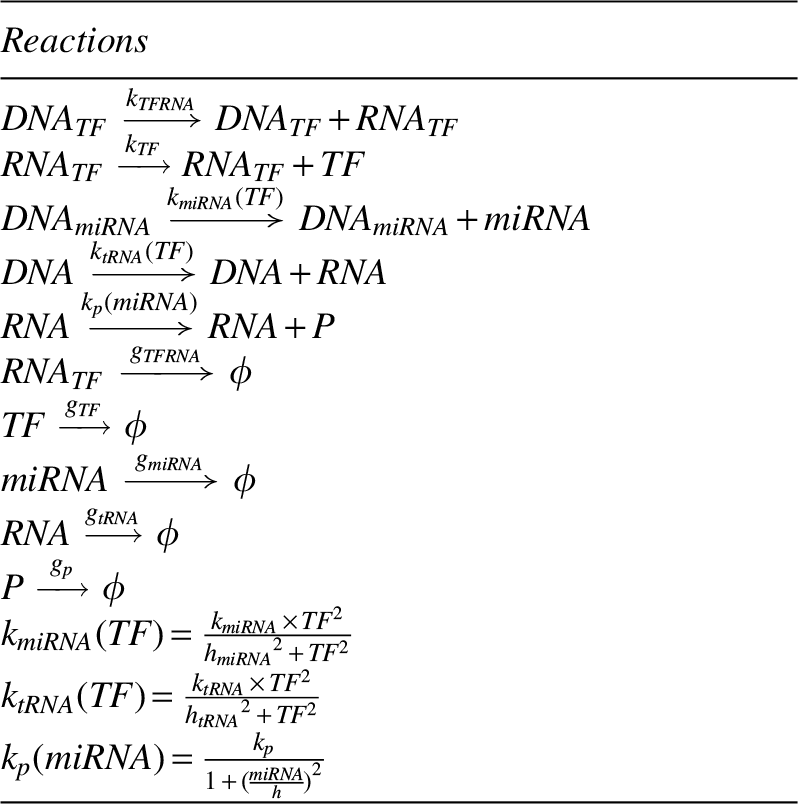
Figure 3 shows that the increment of transcriptional delay increased the COV. The COV of the target protein finally reached a value larger than that of the system without an FFL (blue cross in Fig. 3). Remarkably, for ∼80% of the tunable range, the noise intensity of the target protein appeared to be linear to the time span of the transcriptional delay. Note that the present method of tuning noise does not change the steady-state deterministic behaviors. This fact is clear while taking Laplace transfer of the deterministic equations. The time delay can be easily handled by second shifting theorem, which results in an additional exponential term in the s domain. By applying final value theorem to obtain the steady-state value in the time domain, this additional exponential term became unity. Namely, the transcriptional delay has no effect on the deterministic steady state. Because of this feature, the intensity of the protein noise can be manipulated independently. It provides the possibility to investigate the role of random fluctuations without altering deterministic steady state.
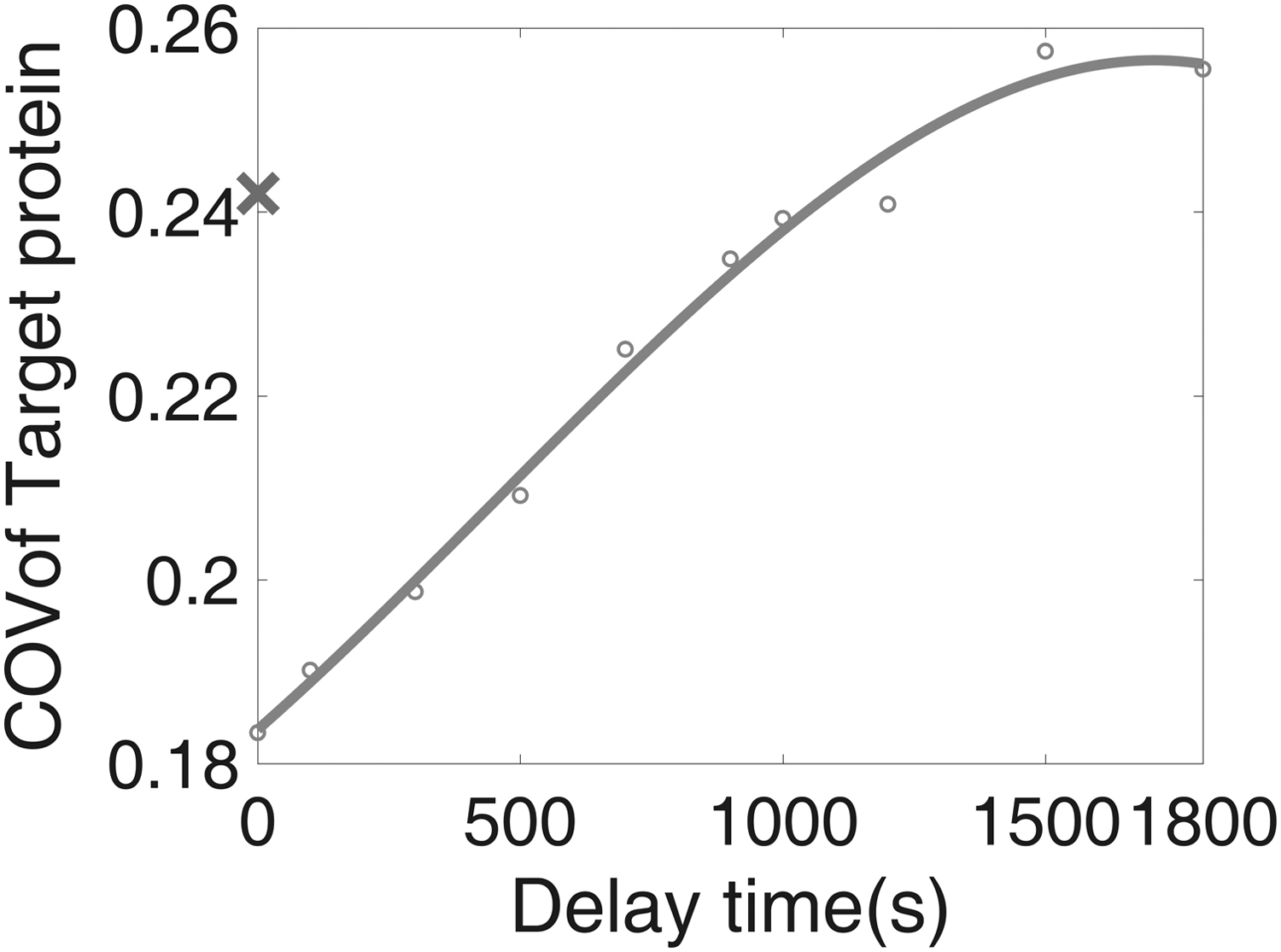
Manipulating noise by the transcriptional delay of miRNA in the incoherent FFL. The circle is the COV of the target protein. For a wide range, the COV of the target protein appears to be linear to the time span of the transcriptional delay. The cross indicates the COV of the target protein without a noise-buffering FFL.
3.3. The distributed delay improves the ability to tune the protein noise
In natural, the time spans of the transcriptional delay appear to be a distribution instead of a single value. Based on experimental observation in the literature (Honkela et al., 2015), we applied the distributed delay. In Figure 4A, the blue cross is the COV of the target protein in the case that the transcriptional delay is a distributed delay. In comparison with the case that the time delay is a fixed value (red circle in Fig. 4A), the blue cross showed improvement of tuning noise. For the delay as a fixed value, the transcriptional delay of 1000 seconds increased the COV of the target protein by 30%, but it became 36% while applying the distributed delay. The distributed delay introduces more noise and this helps manipulate the noise intensity of the target protein.

The linearity of the COV to the time delay.
3.4. The influence of the degradation rate on the noise-tuning method
The degradation rate usually notably affects the noise. We examined the influence of degradation rate on the noise-tuning method. We decreased the degradation rate constants of target RNA and miRNA to 1/10 of the nominal values. To get rid of the effect from the change of particle number, we adjusted the transcription rate accordingly. Namely, the level of the target RNA and miRNA remained the same. Figure 4B suggested that the increment of RNA lifetime slightly shrunk the tunable range.
3.5. The incremental noise of the TF improves the noise-tuning method
We examined how the noise of TF affects the present method of tuning noise at the protein level. We increased the noise intensity of TF by increasing its translation rate constant to 10-folds of the nominal value. To keep the same level of TF, we decreased the transcription rate of the gene encoding TF. In comparison with Figs. 3 and 5A showed that the increment of the noise intensity of the TF facilitated the present method of tuning noise. It enlarged the tunable range to roughly twofolds. This improvement may come from the better noise attenuation of the incoherent FFL. To verify it, we increased the influence of miRNA on the target RNA. Specifically, we changed the value of the parameter h to 20 (particles). To keep the same level of the target protein, we adjusted the transcription rate constant of the target gene,
The influence of the TF noise on the present noise-tuning method.
4. Discussion
In this study, we demonstrated that the noise at protein level was tuned by the transcriptional delay of miRNA in the incoherent FFL. Remarkably, for a wide range, the COV of the target protein appeared to be linear to the time delay of miRNA (Fig. 3). To mimic the situations in nature, we further incorporated the distributed delay and it improved the present method of tuning noise (Fig. 4A). In addition, we explored the role of the noise intensity of the TF. We found that the higher the noise intensity of the TF is, the larger the tunable range of the COV of the target protein (Fig. 5A). Furthermore, when miRNA became strongly interactive with the target RNA, the present methods showed even better ability to tune the noise at the protein level (orange circle in Fig. 5B). The origin of tuning noise comes from the transcriptional delay diminishing the correlation of miRNA and the target RNA, as shown in Figure 5B. Note that, without an FFL, the COV of the target protein is 0.45. While applying the noise-tuning method, the tunable range of COV is from 0.24 to 0.78. It implies the present method can tune the noise intensity in both directions, up or down.
From the experimental observation in the literature (Honkela et al., 2015), the transcriptional delay can be as large as 100 minutes and the peak of its distribution located from 0 to ∼70 minutes, depending on the DNA sequences. There are various origins of transcriptional delay. The elongation of RNA polymerase takes roughly 20 nt/s (Ucker and Yamamoto, 1984; O'Brien and Lis, 1993). The splice of one intron takes ∼0.4–7.5 minutes and the process of transferring matured RNA to cytosol causes a delay of several minutes (Audibert et al., 2002; Zhang et al., 2017). The results of the simulation suggested that the transcription delay within 1000 seconds (16.6 minutes), the COV of the target protein is linear to the time span of the transcriptional delay. The COV reached the maximum value when the delay time approach 30–40 minutes (Figs. 3 and 5). Without a doubt, it is possible to observe all these delay time spans in the real world. In addition, the distributed delay showed the improvement of tuning noise (Fig. 4A). It further concluded that the present method is practically useful. For some other types of delay, that is, transcriptional pause may result in the decrease of RNA and thus changes the deterministic steady states. Our model did not account for it. We, thereby, suggest the present method to be implemented on the system with the transcriptional delay showing little effect on decreasing RNA level.
5. Conclusions
In summary, we demonstrated that the noise at the protein level can be tuned by applying the transcription delay to miRNA in the incoherent FFL. Remarkably, the COV of target protein appears to be linear to the time delay. In addition, the performance of the present method got improved while applying the distributed delay. It implies the present method has the potential to perform better in a practical situation. In conclusion, the proposed method can be incorporated into synthetic biology for manipulating protein noise without changing the deterministic steady-state behaviors.
Footnotes
Acknowledgment
The authors thank Dr. Hsu-Yuan Fu for valuable suggestions.
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This study was supported by grants (MOST 107-2221-E-027-037-) and (MOST 108-2221-E-027-074-MY2) from the Ministry of Science and Technology, R.O.C.